A few concepts should be discussed at the outset before attempting linkage
of functional biology with phylogenetics. This is in order to explain and differentiate
among issues that have not been uniformly understood, advocated, or espoused
by workers in any of the methodological areas related to functional analysis.
To most naturalists functional biology has a unifying significance for evolutionary
biology. This holistic, and Darwinian, perspective of naturalists should axiomatically
apply in the pursuit of phylogenetics; organisms have always represented specific
strategies to differentially survive and reproduce, so this fact should have
a bearing on any estimation of phylogeny.![]() A broadly conceived notion of functional biology is certainly not evolutionary
biology in toto, but it is at the core of biological as well as macroevolutionary
explanations for the history of life. But there are issues related to the term
"function" that need to be examined because these relate not only
to the clarification of the term but also to phylogeny reconstruction itself.
A broadly conceived notion of functional biology is certainly not evolutionary
biology in toto, but it is at the core of biological as well as macroevolutionary
explanations for the history of life. But there are issues related to the term
"function" that need to be examined because these relate not only
to the clarification of the term but also to phylogeny reconstruction itself.
In an attempt to unravel and clarify the several meanings of the term function, a more restrictive use was suggested by Bock and von Wahlert (1965) in a paper on adaptation and the form-function complex. These authors refined the distinction between the mechanical-physiological and behavioral aspects of features of organisms, on the one hand, and the selection-caused (-mediated) biological role (biorole or role; or adaptive or biological function) of these, on the other. Such a conceptual difference was previously well understood by a number of biologists and paleontologists, but it was not appreciated by many others. The concepts of mechanical and biological functions (i.e., function and biorole) were not only clearly differentiated by Bock and von Wahlert but were presented in a theoretically detailed functional and ecological context. This clarification of terms to cover definitionally well-differentiated (but really only empirically divisible) research domains remains important. But the theoretical relationships between mechanics and adaptation are more complex than normal definitional procedures permit (see later).
I and others have followed the terminology introduced by Bock
and von Wahlert (1965) because such practice has the salutary consequence
of forcing both an author and his or her reader to think more clearly about
the complexity of functional, adaptational, and evolutionary issues. In any
meaningful sense, the mechanical and biological functions of organisms in realtime
are inseparable (see Szalay,
1999a for discussion of "realtime"). Yet a combination of historical
and realtime factorsontologically but not epistemically distinctclearly drive
evolutionary changes of functioning attributes.![]() These are wholly interdependent processes and causalities. Note again, however,
that many researchers continue to use the single term "function" with
a clear meaning where the context renders it obvious that they mean either mechanical
or biological (adaptive) function. Others, however, continue to conflate the
meanings at the expense of both operational and explanatory clarity.
These are wholly interdependent processes and causalities. Note again, however,
that many researchers continue to use the single term "function" with
a clear meaning where the context renders it obvious that they mean either mechanical
or biological (adaptive) function. Others, however, continue to conflate the
meanings at the expense of both operational and explanatory clarity.
Lauder (1996), in spite of his attempted endorsement of the functional conceptualization of Bock and von Wahlert (1965), either intentionally or inadvertently confused this approach with "ahistoricity" versus "historicity." When Lauder (1996, p. 63) states that "... a bone might have the mechanical function of stiffening the limb against gravity ..." he is not only referring to mechanical function. In making such a statement that connects an organism (and, needless to say, the size of the animal, and the manner of its locomotor and other positional behaviors) to its specific environmental context, Lauder clearly adds an adaptational, and therefore selectional force-related, component to his characterization, which he refers to as "anatomical function." He obviously believes that there is a difference among the relationships of an anatomical structure, its biological role(s), and what he pursues as the neurologically controlled behaviors of an organism. In making this distinction, he either rejects or misses the universality of meaning attached to the function and biorole dichotomy so well explicated by Bock and von Wahlert. Differently structured mechanical systems (e.g., his examples of forearm mechanics), although performing the "same" roles, surely have a distinct combination of mechanical factors and resolution of forces acting differently in concert. Such systems certainly do differ in their mechanics. Model-T Ford and late-model BMW automobiles all roll along the same road from point A to B, but no one would seriously entertain the notion that their detailed (progression-related) mechanics are comparable.
This issue of function is further confounded by Lauder (1996, p. 63) when he notes that "[t]he biological context of structure and function is referred to as the `biological role' of a structure or mechanical function..." (emphasis added). In spite of what I believe to be Lauder's misuse of the conceptual and terminological meanings of function, these distinctions continue to serve a significant purpose in comparative biology. When viewed theoretically, both the mechanical and biological functions can change (e.g., from bones to neuronal networks). Both can change independently of one another and have independent historical components. It is important, therefore, not to front-load the historical component (the structural-functional heritage) with fitness considerations within the already enormously complicated theoretical maze regarding function. By not keeping the biological role independent of mechanical explanation, Lauder has done just that.
The "new adaptationism" that has become wedded to a taxic conceptualization of parsimony-based cladistics, and has been hailed by many (e.g., Novacek 1996) as the solution to the problem of functional conceptualization has not advanced the cause of either terminological or theoretical clarity. The "causal role function" notion in this "new adaptationism" is an unnecessary and invalid subordination of the adaptational component of phylogeny to a theoretically truncated (= ontologically unrealistic) epistemic paradigm of numerical cladistics. This view of the cladistic component of phylogeny has come to mean not only that taxograms are phylogeny, but that evolutionary explanations (and even theory) should be based on such taxic constructs.
Gould
and Vrba (1982) offered a terminological scheme through which they asserted
that one can clarify differences between adaptations due to natural selection
for their current role, and various aspects of such features that were not built
by selection for their current role, but became useful for it after their inception.
These authors argued that this was partly an effort to replace "preadaptation",
a concept they considered teleological. According to Gould
and Vrba (1982), "aptation" was the general phenomenon of something
being fitted for a role, whereas the term "adaptation" was restricted
for traits built by selection for current roles and the term "exaptation"
was used for the concept of preadaptation.![]()
The example they used, feathers (without a much- needed clarification of the kind of feathers), has a considerable semantic element to it. The term "feather" covers various kinds of feathers, the expected state of affairs for all evolutionary transformations of attributes through phylogeny, each representing part of a continuous, but at any historical moment possibly distinct, condition of that trait or group of traits. Although feathers are a general developmentally circumscribed group of features, it is obvious that whatever kind of feather came first in a particular lineage (probably the protofeather for improved thermoregulation, from a preexisting condition that we would call a type of scale) was not the same kind of feather that was subsequently developed through selection for flight, or the dazzling variety of display feathers developed through sexual selection in birds. As noted, we could also have begun by calling feathers a particular type of denticulate scale along a continuum of scales (and of bushy phylogenetic patterns), a sequence that undoubtedly was part of the transformation series of scales in general. The same applies for vertebrate limbs, from fins to wings of all sorts. This highly semantic, as well as "lurching" and "jerky," conceptualization, just one of the many possible, played a major role in Gould and Vrba's theoretical presentation.
But the proposed sharp division between "exaptations" and adaptations does reflect two important underlying messages, and the agenda is clearly punctuationist. First, and this view is not particularly controversial, lots of features just happen; they are non-adaptive. The second point, however is that the advocated (punctuationist and taxic) view of evolutionary change should have a distinct "aptive" component assigned to (punctuated) species taxa (either the first or the second kind of "aptation"). Furthermore, and subsequently, a lineage perspective on evolutionthe Darwinian viewshould be abandoned in favor of a "stable-taxon"-based notion of punctuationism.
This was a very different conceptualization from Simpson's (1953, p. 160-199) expansion on Darwin, namely that the prospective functions in a dynamic overlap of the shifting relationship between a lineage and the environment become realized functions (including their phenotypic expressions as adaptations). Simpson envisaged the adaptive process as an ongoing seamless relationship between the total environment and the evolutionary unit at a given moment in the history of that lineage. Gould and Vrba (1982), on the other hand, based their discussion on the underlying assumption (never made explicit) of the mysterious and abrupt shifts of speciation. Their conceptualization, and the resultant and ungainly terminology loaded with unacceptable theory, is based on the coded core of punctuationism, paleontological species taxa that happen to coincide with the terminal species taxa of cladistics.
The mental imagery offered by the punctuationist and taxic terminology is clear:
Species give rise to other species with sharply distinctive adaptations, a perspective
that studiedly underplays the rate-independent gradualism of phyletic continuity,
and exhorts terminology related to species taxa that serves a speciational theory
of evolutionary change.![]()
An issue of seemingly remote relevance to the theoretical notions embedded
in "function" is the dichotomy of "proximate" versus "ultimate"
designation of causes in biological explanations. Mayr's
(1961) early, popular, and much repeated thesis (Mayr
1982) was that there are both proximate and ultimate causes operating in
biology. The former (the mechanics, physiology, etc., of an organism) was pursued
by the "functionalists," and the latter (the adaptations and history
of species) by the "naturalist-evolutionists." Mayr believed that
sharply different conceptual approaches were needed to understand these causes.
I take a contrary position. The concept of causality in realtime (Szalay
1999a) should play a fundamental role in any theoretically rigorous connection
among the various components of both "nonhistorical" and "historical"
causation and explanations in holistic functional biology. (Note that the latter
is considered biosystemism by Mahner
and Bunge 1997, who maintain the special position of organisms in the various
levels of organismic and suborganismic diversity.) Therefore a loose designation
of cause can be obfuscating when it comes to either functional or evolutionary
analysis, both in the broad and narrower senses (see later). Causes occur in
realtime (and that rules out "causal" past history), although obviously
the initial and boundary conditions of organisms living at a particular time
play a significant constraining and facilitating role throughout their entire
life. The initial and boundary conditions together are a rough, if not exact,
equivalent of Aristotle's material cause from his "techne" model of
causation. I adopt the view of Mahner
and Bunge (1997) on causality, except for their insistence that ontogeny
is not causal.![]() The arguments of Mahner and Bunge challenge the notion of "ultimate causality"
outside of the context of realtime causality in any lineage.
The arguments of Mahner and Bunge challenge the notion of "ultimate causality"
outside of the context of realtime causality in any lineage.
It is important to frame both questions and research strategy not in terms
of almost invariably nebulous "ultimate causes" lost in the past,
but rather in terms of the knowable but obviously constrained material realities
of the organism we study, be it fossil or extant.![]() It is obvious that any manifestation of the organism throughout its entire life
history is deeply functional in the broad sense, and any such functions are
as historical as they are adaptive. Furthermore, a phylogeny is tested against
these, in contradistinction to the "new adaptationism" paradigm that
would map features discovered in these contexts onto a taxogram. Organisms themselves
reflect both history and adaptations in an inexorably intertwined manner that
should be analyzed to factor out the historically shared (or sequentially related)
components of phylogenetics.
It is obvious that any manifestation of the organism throughout its entire life
history is deeply functional in the broad sense, and any such functions are
as historical as they are adaptive. Furthermore, a phylogeny is tested against
these, in contradistinction to the "new adaptationism" paradigm that
would map features discovered in these contexts onto a taxogram. Organisms themselves
reflect both history and adaptations in an inexorably intertwined manner that
should be analyzed to factor out the historically shared (or sequentially related)
components of phylogenetics.
Historical narratives are histories of successful and successive ontogenies
wastefully managed by ecology,![]() and lineages represent ontologically the past history of evolving populations
or species (anywhere along that lineage in time). Therefore, a lineage (a mere
record of a species) does not have "causes" acting on it, beyond those
that acted on the aggregate of living individuals that made up each segment
in the past (a species, or other evolutionary subunits of a species, in an instant
in time; such realtime units are not species taxa; see discussions in Bock
1979; Szalay 1999a).
Real causes that affect, for example, the becoming of a single organism are
demonstrated by the newly emerging "hybrid" fields (e.g., developmental
genetics dealing with issues of evolutionary homology, often called "EvoDevo").
These disciplines show with increasing precision how inseparable functional
explanations are, not only from history, but also from adaptive existence. Mutations
and their developmental consequences are inseparable from the context of a particular
genotype. This is well beyond the occasionally agenda-laden and quasi-engineering
concepts of function that eschew adaptations and adaptedness, such as the kind
of functional conceptualization advocated by Cummins
(1975), Amundson
and Lauder (1994), and others.
and lineages represent ontologically the past history of evolving populations
or species (anywhere along that lineage in time). Therefore, a lineage (a mere
record of a species) does not have "causes" acting on it, beyond those
that acted on the aggregate of living individuals that made up each segment
in the past (a species, or other evolutionary subunits of a species, in an instant
in time; such realtime units are not species taxa; see discussions in Bock
1979; Szalay 1999a).
Real causes that affect, for example, the becoming of a single organism are
demonstrated by the newly emerging "hybrid" fields (e.g., developmental
genetics dealing with issues of evolutionary homology, often called "EvoDevo").
These disciplines show with increasing precision how inseparable functional
explanations are, not only from history, but also from adaptive existence. Mutations
and their developmental consequences are inseparable from the context of a particular
genotype. This is well beyond the occasionally agenda-laden and quasi-engineering
concepts of function that eschew adaptations and adaptedness, such as the kind
of functional conceptualization advocated by Cummins
(1975), Amundson
and Lauder (1994), and others.
I am not critical of the epistemologically significant distinction found in practical research programs that pursue sundry important functional aims in the laboratory without immediate concern for field-related bioroles. But we are repeatedly reminded of the fact that the concept of function is understood very differently by many who consider themselves to be structuralists, functionalists, phylogenists, or adaptationists; a somewhat unfortunate, although often understandable, compartmentalization of what should be a theoretically cohesive whole.
In addition to the varied meanings of function, for example in the community of students who study musculoskeletal morphology, there are a variety of technical approaches that require novel conceptualizations. New approaches addressing questions on sundry levels and that apply equally to skeletons, bones, trabecular structure, and bone tissue types have been, and are being, developed. Such epistemological avenues rest on different assumptions from level to level, as researchers consider bone tissues, bones, and skeletons from different perspectives. Nevertheless, beyond the basic epistemological issue (namely that there are clear conceptual differences between mechanical functions and biological roles), there has been an increasing conceptual redefinition and narrowing, as well as confusion, concerning the theoretical meaning of functional studies. Theoretical positions have been taken by authors proposing various exclusive meanings for the concept of function. Whereas some of these positions are largely epistemological in nature, some have taken on the cloak of ontological propositions in claiming some special theoretical virtue for them.
For example, in a long series of papers going back two decades, Lauder (see
Lauder 1981; Amundson
and Lauder (1994); Lauder
1996; and references therein) advocated a rather narrow view of functional
morphology that sought connections between the parsimony-based taxograms of
"pure" systematists and a quasi-engineering type of functional analyses.
These efforts either implicitly or explicitly eschewed connections to adaptation,
and they were devoid of any input from a selectional (= Darwinian) perspective.
In fact, in Amundson
and Lauder (1994) the claim was made that adaptations are all but impossible
to identify and therefore the search for the "why" of attributes (namely
what selectional causes molded a feature) is doomed to failure. The justification
for this narrow view of functionalism (actually a form of structuralism, see
later) is that they believed that the mechanical transformation of structures
is what "functional" biology is about, without regard for the specific
time-contextual roles of natural and sexual selection. Yet, this theoretically
restricted perspective did not prevent Amundson
and Lauder (1994) from calling their conceptualization, inexplicably, "causal
role function."![]()
This "causal role function" perspective (with its thinly disguised
disdain and its rejection of the "less rigorous" research under "old
adaptationism" aimed at uncovering adaptive functions; see Rose
and Lauder 1996) is a direct outgrowth of the "cladogram first"
research program (e.g., Eldredge
and Cracraft 1980; Lauder
1981). This strictly conformist and fundamentally Kuhnian research program
subjugates biological character analysis to taxic schemes of bifurcations, and
foregoes the necessary prerequisite of testing character transformations with
the aid of methods derived from tested evolutionary theory (i.e., the phyletics
of features). But any evolutionary scheme or theory is wanting and distorted
without adaptation as part of the evolutionary process.![]() Ironically, such a pattern of cladogeny is just as unknowable in terms of certainty
(particularly without phyletics, and not only because the concept of scientific
corroboration of an ancestral stage is excluded from cladistic theory) as is
the adaptive function jettisoned by Lauder and colleagues. In fact, there are
gradational or incremental probabilities in the testing procedure (corroboration,
verification, or rejection), often with a considerable overlap between both
phylogenetic and adaptational hypotheses. It is such overlaps in the analysis
that allow for the distinction between homologous and homoplasious similarities.
Ironically, such a pattern of cladogeny is just as unknowable in terms of certainty
(particularly without phyletics, and not only because the concept of scientific
corroboration of an ancestral stage is excluded from cladistic theory) as is
the adaptive function jettisoned by Lauder and colleagues. In fact, there are
gradational or incremental probabilities in the testing procedure (corroboration,
verification, or rejection), often with a considerable overlap between both
phylogenetic and adaptational hypotheses. It is such overlaps in the analysis
that allow for the distinction between homologous and homoplasious similarities.
On theoretical grounds that seriously considers the role of developmental biology (particularly modularity) in the evolutionary process and the irrelevance of parsimony-based decision making for character transformation, I do not believe that an engineering analysis of taxa based on their position on taxograms is a meaningful approach to phylogeny estimation. This approach, which depends on a misplaced fealty to someone else's cladogram (and to the parsimony paradigm) to explain evolutionary transformations, is flawed at its foundations by its circularity. Yet, obviously functional, phyletic, and cladistic, evaluations of distributional data must be an integral part of reliable phylogeny estimation. Both the establishment and the explanation of meaningful patterns (not just the structural and mechanical manifestations of shape) require some adaptationally-framed assessment to contextualize polarity determination, as well as the stratigraphical and biogeographical contexts, all at a level that information and the applications of valid methods permit. This is an attempt to understand the phyletics of features. These strictures apply proportionately to the degree that any of these activities can be brought to bear on a research project. This, in turn, obviously depends on the nature of available evidence (including both deductive and inferential information). But the various levels of inference permitted by data and context do not alter the validity of a conceptual methodology derived both from descent and from natural and sexual selection in a specific space and time. To claim theoretical primacy for a phylogeny based on statistical sorting of structural patterns, combined with only a sensu stricto functional analysis which would transfer functional evolution into a non-Darwinian vacuum is not based on sound theory. It is a flawed conceptual method for attaining an understanding of "functional evolution" as its professed goal. The concluding remarks of Leroi et al. (1994, p. 398) regarding phylogenetics are significant in this context. "Such analyses will be all the more robust for lacking unsupported assumptions, unwarranted inferences, and untestable hypotheses about the history of evolutionary mechanisms." It can be added that these linear analyses of nonlinear relationships are also not only sterile, but theoretically vacuous.
 The
theoretical stance of Amundson
and Lauder (1994), in which they attempted to exile
Darwinism from functional biology, is in essence a narrow
functionalist (structuralist) exegesis of the early wave of
zealotry of Anglophone cladistics that declared its total
independence from evolutionary biology and claimed primacy
for nested cladograms of taxa, taxograms, before all else.
The additional claim for primacy for a pure mechanics-based
view of function in biology also reflects a view that
regards deduction from physical laws as superior to the
inductive procedures necessary for historical explanations.
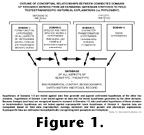
Because a strong inductive component is required in all
adaptational and historical assessments of patterns (see Bock,
1981; Szalay
and Bock 1991), the connections of functional-adaptive
analysis (including transformational analysis) to
phylogenetics (see Figure
1) is usually unacceptable for doctrinaire cladists and
"new adaptationists." Although parsimony-based
cladistics strives to recover history, it also aims for a
completely deductive, Popperian methodology (openly admitted
only by pattern cladism; see especially Schuh
2000).
The
theoretical stance of Amundson
and Lauder (1994), in which they attempted to exile
Darwinism from functional biology, is in essence a narrow
functionalist (structuralist) exegesis of the early wave of
zealotry of Anglophone cladistics that declared its total
independence from evolutionary biology and claimed primacy
for nested cladograms of taxa, taxograms, before all else.
The additional claim for primacy for a pure mechanics-based
view of function in biology also reflects a view that
regards deduction from physical laws as superior to the
inductive procedures necessary for historical explanations.
Because a strong inductive component is required in all
adaptational and historical assessments of patterns (see Bock,
1981; Szalay
and Bock 1991), the connections of functional-adaptive
analysis (including transformational analysis) to
phylogenetics (see Figure
1) is usually unacceptable for doctrinaire cladists and
"new adaptationists." Although parsimony-based
cladistics strives to recover history, it also aims for a
completely deductive, Popperian methodology (openly admitted
only by pattern cladism; see especially Schuh
2000).
The combined perspectives of parsimony-based cladistics and "causal role
function" protocols that consider adaptational understanding unattainable
are being made obsolete by many multidisciplinary efforts created from the fusion
of genetic, morphological, developmental, ecological, and paleontological perspectives
and practices, to mention only a few.![]() The deduced engineering patterns in a "causal role function" approach,
of course, can be superimposed onto, if not meaningfully interdigitated with,
parsimony-based cladograms. Such practice of circularity is essentially similar
to attempts at a "transformational analysis" that hope to procure
evolutionary transformations from taxograms.
The deduced engineering patterns in a "causal role function" approach,
of course, can be superimposed onto, if not meaningfully interdigitated with,
parsimony-based cladograms. Such practice of circularity is essentially similar
to attempts at a "transformational analysis" that hope to procure
evolutionary transformations from taxograms.![]() So, in contradistinction to the paradigm of "causal role function"
advocated by Amundson
and Lauder (1994), such restrictive functional approaches lack a valid framework
for evaluation without the ecological (adaptational) component, as vague as
adaptive meaning may be in some instances. The pivotal role of contingency-based
evolutionary analysis means a contextually variable dependence on adaptation-related
considerations. These are manifested (e.g., in vertebrate paleontology) mostly
as structural patterns of aspects of skeletal remains.
So, in contradistinction to the paradigm of "causal role function"
advocated by Amundson
and Lauder (1994), such restrictive functional approaches lack a valid framework
for evaluation without the ecological (adaptational) component, as vague as
adaptive meaning may be in some instances. The pivotal role of contingency-based
evolutionary analysis means a contextually variable dependence on adaptation-related
considerations. These are manifested (e.g., in vertebrate paleontology) mostly
as structural patterns of aspects of skeletal remains.
I include here a relevant, albeit rhetorical, query. What does the comparative assessment of engineering parameters mean without any conceptual room left for the reasons for these differences? Whenever civil or aeronautical engineers plan the construction or analysis of structures, there is always a particular goal in mind. That goal is, properly, the Aristotelian final cause, namely "what something is for." This implicit factor is absolutely and fundamentally connected to any mechanically oriented enterprise, and if such an effort is separated from its context it loses not only its directives and limits but also its meaning. Unlike the "causal role function" protocols, Bock (1999), who also maintains a sharp divide between function and adaptation epistemologically, makes the obvious connection between these efforts. In Bock's view, functional and adaptational or utilitarian analysis come under the same conceptual and theoretical heading; both are functional explanations.
It is hard to contemplate how a particular well-substantiated biorole (or one
inferred with a lesser degree of probability than samples of direct observations
provide), correlated with form-function, would not have played a major causal
evolutionary role in the becoming of that complex, except in a strongly structuralist
conceptualization of biodiversity. An increasingly narrow emphasis on engineering
function can result in a peculiar evolutionary conceptualization, not to mention
the problem of discordant semantics. Is such conceptualization really good theory?
The fundamental insight of Lamarck, reworked within Darwinian theory, that the
combined function and role of attributes is part of the causal interaction responsible
for their evolution has been pursued by many biologists and philosophers; its
theoretical consequences are not discussed here.![]() In a definitional sense, at any moment in lineage history a form-function complex
that exists is a set of initial conditions and the sundry causes that act on
the organisms are, strictly speaking, the causes that mediate subsequent evolution
(survival and resulting fitness differences). But it cannot be overlooked that
the very phenotype-based activities of organisms generate a great deal of the
specific causal forces acting on them, as has been repeatedly pointed out. The
feedback loop is mutually dependent, and the understanding of the process fully
justifies an expansion of the conceptual methodologies based on it. Epistemological
advances follow such a theoretical stance.
In a definitional sense, at any moment in lineage history a form-function complex
that exists is a set of initial conditions and the sundry causes that act on
the organisms are, strictly speaking, the causes that mediate subsequent evolution
(survival and resulting fitness differences). But it cannot be overlooked that
the very phenotype-based activities of organisms generate a great deal of the
specific causal forces acting on them, as has been repeatedly pointed out. The
feedback loop is mutually dependent, and the understanding of the process fully
justifies an expansion of the conceptual methodologies based on it. Epistemological
advances follow such a theoretical stance.
But what about the specific research protocols that should rest on conceptual
foundations different from those of engineers, if the goal is an evolutionary
understanding? Strictly functional explanations, although they may justify deductive
issues dealing with features, are incomplete for species-specific phenotypic
attributes. To recast and paraphrase Dobzhansky, they lack explanations for
their species-specific, hence idiosyncratic, workings. The perspective criticized
here, repeated for decades both by doctrinaire cladists, as well as by the pure
"functionalists" such as Lauder and colleagues, is quite different
from Bock's frequently taken stance (e.g., Bock
1988) that functional (sensu stricto) and ecological components of, for
example, morphology, are independent and complimentary. Nevertheless, Bock does
profess a belief that a sharp theoretical dichotomy regarding function and adaptations
is the proper view, and that the investigations of the latter should depend
on the former.![]() Bock's theoretical perspective appears to be tied to Mayr's
(1961) claim regarding distinct proximal (which Bock called functional)
and ultimate (which Bock called evolutionary) causes (which Bock more meaningfully
called explanations). Bock
(1999) maintained that functional morphology answers "how" questions,
namely how things work. I, and others, obviously do not disagree. But the questions
and problems of functional biology (sensu lato, the sense endorsed here for
the theoretically most inclusive concept) arise from a much broader context
than just structural statics and mechanics, or chemistry. Bock's own view on
evolutionary explanations that these are responses to questions of "...
why attributes of organisms came into being originally and have modified (=
evolved) over historical time" (1999,
p. 49), indicates, however, that a certain critical theoretical perspective
is missing.
Bock's theoretical perspective appears to be tied to Mayr's
(1961) claim regarding distinct proximal (which Bock called functional)
and ultimate (which Bock called evolutionary) causes (which Bock more meaningfully
called explanations). Bock
(1999) maintained that functional morphology answers "how" questions,
namely how things work. I, and others, obviously do not disagree. But the questions
and problems of functional biology (sensu lato, the sense endorsed here for
the theoretically most inclusive concept) arise from a much broader context
than just structural statics and mechanics, or chemistry. Bock's own view on
evolutionary explanations that these are responses to questions of "...
why attributes of organisms came into being originally and have modified (=
evolved) over historical time" (1999,
p. 49), indicates, however, that a certain critical theoretical perspective
is missing.
This quasi-artificial "functional" versus "evolutionary"
theoretical dichotomy highlights a lack of formal recognition that in any evolutionary
change the usual "why" question alone is not an adequate formulation
of the nature of evolutionary explanations. Evolutionary functional explanations
(sensu lato) have an ineluctable transformational component that render mechanical
or role-related notions of function incomplete and therefore theoretically inadequate.
In fact, any full evolutionary explanation that deals with specific lineages
of organisms has, as its integral component, the issue that how something works,
in light of what it is for. And therefore it is critically dependent on how
a particular trait prevalent in a lineage came to be historically transformed
and ontogenetically constructed that way. At first this may appear to be insignificant.
But on second thought, it should be realized that contingent lineage-specific
changes are not "merely what happened" (contra Bock
(1999), p. 56). Such changes, the selectional guidance notwithstanding,
were profoundly influenced or guided by preexisting stages (as they will have
influenced subsequent changes). There is virtually universal agreement that
these contexts (the heritage) guide both the nature of transformations as well
as their adaptive "goodness," "latitude," and even built-
in seeds of doom (in retrospect), or new opportunities of given lineages in
given environmental or competitive contexts they will encounter.![]() Realtime constraints (rooted in history) do not only constrain, but they direct
and facilitate subsequent modifications.
Realtime constraints (rooted in history) do not only constrain, but they direct
and facilitate subsequent modifications.
Such conceptualization should have profound implications for methodologies that aim to decipher evolutionary history. For example, the literature on the history of understanding the origin, transformation, and function of tribosphenic teeth of therian mammals, and the resultant evolutionary patterns constrained by the genetic and occlusal dynamics of the original molar system supply some of the most numerous and persuasive examples of the meaning of constraint. Both the historical and the functional (occlusal and ecological) constraints profoundly affected morphology, function, and biological roles throughout phylogeny. Examples abound in virtually all systems of vertebrates, the skeleton being no exception.
Bock's (1988, p. 207) view that adaptational explanations are not evolutionary but functional, therefore, is also problematic, particularly because Bock (1999) continues to maintain the legitimacy of the theoretical independence of functional (proximate) and evolutionary (ultimate) explanations. Explanations of adaptations are in fact inseparably both; they are "temporally-looped" rather than hierarchically and dependently related to form-function. While natural and sexual selection are law-like explanations, their consequences in lineages are also fundamentally historical and time-dependent processes that are never disembodied from the particular lineage histories (e.g., Szalay and Costello 1991). As Bock (1993) so aptly emphasized, natural selection is cause, mechanism, and process, depending on our use of and the context of the concept. The acquisition of new adaptations or a new form of adaptedness (both are processes) is a large component of evolutionary history, and much of phenotypic evolution, as most evolutionists suspect (and long championed by Bock), is fully Darwinian; it is causally propelled at the organism level (see also the organism-centered biosystemist approach of Mahner and Bunge 1997).
Ecomorphology (or any aspect of the phenotype related to bioroles) is conceptually very much a part of this endeavor to understand attributes because this field forms the context with its inseparable feedbacks into the strictly functional efforts in laboratories. The deductive base of these efforts does not make them superior or axiomatically antecedent to ecomorphology. Although Bock (1999) argued for the distinction of ecological morphology from functional morphology and the notion that the former is built on the latter, he nevertheless fully endorses the importance of adaptational analysis. This is in contrast to Lauder and colleagues (see Lauder, 1990, 1996; Leroi et al., 1994), whose expressed futility regarding adaptations is more in service of "causal role function" than any theoretically and empirically tenable position in Darwinian theory. In spite of his stance, however, Bock (e.g., 1977, 1981) applied what I believe to be a somewhat restrictive, more logic- than probability-based perspective when it comes to extinct organisms. Given good and complete specimens, specific adaptive strategies can be understood in fossil taxa with high confidence, because the judicious and phyletically contextual use of combined mechanical, adaptational, and ecological models of living organisms can render great certainty to this type of paleobiological analysis. This type of modeling through the use of carefully delineated aspects of living species can yield an important understanding for both adaptational and phylogenetic analysis (Szalay and Sargis, in press).
It bears repeating here that very often we can corroborate far better the adaptive strategies of well- known fossil species than their phylogenetic affinities. I suggest, therefore, that perhaps the construction of completely deductive, syllogism-like, ontological statements that are deduced from the logical consequences of otherwise clear empirical research-based definitional differences (such as the distinction between function and biorole) is not the most useful approach to theorizing in this area of evolutionary analysis. Such assertions carry well-corroborated definitional concepts to their logical extreme, but not necessarily to new, theoretical heights. Both the deductive and the purely epistemological limitations, as in the case of strict functionalist theory, are unjustified.
Let me also reiterate the obvious. First, the level at which adaptations and adaptedness may be ascertained in organisms is unequivocally the highest for living species observed in nature. But lest we forget, it is equally obvious that existing features in an organism may reflect causal correlations between these form-function complexes and the use they had in the past. In fact, the very logic of descent with adaptive modification demands that, given the temporal nature of the evolutionary process, it be understood clearly that the current attributes of organisms are the products of selectional forces acting on past generations. Activities at a given time (in the particular context of the existing environment) generate the selectional forces that either maintain or transform the frequencies of "fit" individuals with the appropriate phenotypic attributes. But this frequency-changing (or -maintaining) response regarding the adaptations is in the succeeding populations. This temporal relationship is critically important.
The only (probability-based) factor that is likely to assure successful programs in adaptational studies (those with high truth content) is based on the realistic assumption that the rate of environmental change is usually slow in relation to generation time (certainly gradual or static, short of extraterrestrial impacts or violent local disturbances), or often approaches zero (in generational terms), resulting in stabilizing selection, often with a fully Darwinian progressive change for the same biological functions. What organisms do, therefore, in their currently observed environment is probably causally connected to their features.
In addition to the foregoing general theoretical point regarding the absolutist perspective on the study of adaptations in our own time, there are several other issues that have surfaced in regards to an exegesis of the logic of "current utility." The statement by Bock (1999, p. 55; and personal communications, 1999) that the "[s]tudy of biological roles must be done by observations of the organism living naturally in its environmentthey cannot be determined by observations made in the laboratory or other artificial conditions" is a case in point. No one would argue that for highly specific activities contained in the form-function features of living organisms information from the natural setting is critical. But we can counter that a host of highly relevant explanations about fossil taxa, using rigorously constructed models already alluded to, are equally justified, often approaching a high degree of probability at given levels comparable to those made about living organisms. In fossil mammals, based on this type of rigorous and relevant modeling, there are role-related attributes at these levels such as cursorial modifications, aquatic adaptations, digging adaptations, habitual hanging, obligate arboreality or terrestriality, and hosts of obligate dietary adaptations such as grazing, myrmecophagy, piscivory, and hypercarnivory, all on a level of generality that can be securely ascertained (Court 1994; Szalay 1994; Szalay and Lucas 1996; Szalay and Schrenk 1998; Szalay and Sargis, in press; and references therein). A particularly excellent example is Court's (1994) analysis of limb posture and gait in the primitive Eocene fossil proboscidean Numidotherium. Such modeling employs ecologically and functionally well-understood living species that cross size and higher taxon limits, thus providing a degree of adaptational assessment through the convergence method, which is not only highly reliable, but which forms a critical link to other activities such as phylogenetic estimations (e.g., Szalay 1981; Szalay and Sargis, in press). As in any other methodology axiomatically derived from underlying tested theory, none of this is logically full proof but rather is probability-based.
Therefore, to consider the assessment of causal factors for the features that derive from the obligate activities of organisms to be a method in ecological morphology that is applicable to living species only is unfortunate. It appears to be a monotonic logic-bound consequence that unnecessarily truncates adaptational analysis, which is always probability-based. It also unjustifiably narrows the valid epistemological limits of the testing of a tremendous range of issues in macroevolution. I emphasize again that adaptational assessment of well-known fossils is not any less dependent on probability judgements than is phylogeny estimation in general.
![]()