SYSTEMATIC
PALEONTOLOGY
DINOSAURIA Owen 1842
SAUROPODA Marsh 1878
TITANOSAURIA
Bonaparte and Coria 1993
MALAWISAURUS
Jacobs, Winkler, Downs, and
Gomani 1993
Type Species.
Malawisaurus dixeyi (Haughton 1928)
Malawisaurus dixeyi
(Haughton 1928)
Figures 4-27, Tables
2-8
Emended Diagnosis
Malawisaurus is
characterized by high-angled premaxillae; posteriorly broad, angled
infratemporal fenestrae; short and divergent basipterygoid processes; dentary
with 15 alveoli that extend two thirds the length of the element; thin splenial
with a spenial foramen; broad but not spoon-shaped teeth with high-angled wear
facets; undivided cervical neural spines and pleurocoels; cervical
prezygapophyses extend beyond the anterior end of the centrum; presence of pre-
and post-spinal laminae in distal cervicals; ribs in proximal and middle
cervicals extend beyond the end of the succeeding centrum; elongate, eye-shaped
pleurocoels on dorsal vertebrae; infradiapophyseal laminae of distal dorsals not
forked; six sacral vertebrae; sacral neural spines fused into a dorsal plate
that overhangs the neural arches; strongly procoelous proximal but platycoelous
middle and distal caudals; proximal caudals with well-developed pre-spinal and
post-spinal laminae; at least five V-shaped middle chevrons; and metacarpals
with distal articular facets.
Taxonomic Note
Malawisaurus
dixeyi was initially named Gigantosaurus dixeyi by
Haughton
(1928) who considered the specimen to be closely related to Tanzanian specimens
that were referred to Gigantosaurus (Fraas 1908). However, this generic
name was preoccupied so the generic name for the Tanzanian specimens was changed
to Tornieria (Sternfield 1911). Without justification, G. dixeyi
was referred to as Tornieria dixeyi. The generic name Tornieria
was later changed to Janenschia by
Wild (1991). The specimens from Malawi
that passed through these generic name changes or were collected subsequently
are from the same area and same rock unit appear specifically identical, but are
distinct from Janenschia from older beds in Tanzania. Therefore,
Jacobs
et al. (1993) erected a new generic name, Malawisaurus, to accommodate
the Malawi taxon.
Geologic Age and
Distribution
Early Cretaceous,
Mwakasyunguti area, Karonga District, Malawi, Africa.
Description
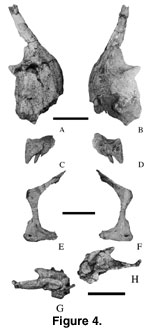 Premaxilla.
Except for the partial braincase that includes several bones, all cranial
material was disarticulated. The right premaxilla (Mal-6; length = 45 mm; height
= 65 mm; Figure 4A-B) with three unerupted teeth and four alveoli was briefly
described by
Jacobs et al. (1993, figure 1a). The body of the premaxilla is
subrectangular in lateral view. The median symphysis is broad and oblique in
medial view. The anterior margin of the premaxillary body rises nearly
vertically from the dental margin in lateral view. The nasal process is narrow,
anteroposteriorly flattened, dorsally oriented, and borders the large external
naris anteriorly (Jacobs et al. 1993). The maxillary suture is tall and is
immediately below the external naris. The premaxilla has a dorsal process and a
dorsoventral groove ventral to the maxillary process that suggest the presence
of a dorsoventral process of the maxilla.
Premaxilla.
Except for the partial braincase that includes several bones, all cranial
material was disarticulated. The right premaxilla (Mal-6; length = 45 mm; height
= 65 mm; Figure 4A-B) with three unerupted teeth and four alveoli was briefly
described by
Jacobs et al. (1993, figure 1a). The body of the premaxilla is
subrectangular in lateral view. The median symphysis is broad and oblique in
medial view. The anterior margin of the premaxillary body rises nearly
vertically from the dental margin in lateral view. The nasal process is narrow,
anteroposteriorly flattened, dorsally oriented, and borders the large external
naris anteriorly (Jacobs et al. 1993). The maxillary suture is tall and is
immediately below the external naris. The premaxilla has a dorsal process and a
dorsoventral groove ventral to the maxillary process that suggest the presence
of a dorsoventral process of the maxilla.
Maxilla. The
maxilla (Mal-106; Figure 4C-D) is represented by a left anterior fragment with two
replacement teeth present. The first tooth alveolus is 60 mm from the premaxilla-maxilla contact. The teeth are aligned nearly parallel to the
anteroposterior axis of the preserved maxilla.
Jugal. A right
jugal (Mal-44; Figure 4E-F) is a small, thin (width = 30 mm; height = 120 mm)
bone, similar to that of Camarasaurus lentus (Madsen et al. 1995,
figures 1e and 6c) in lateral view. The quadratojugal process is mediolaterally
flattened while the postorbital process is mediodorsally curved to accommodate
the jugal process of the postorbital. The posterodorsal curvature of the
postorbital process suggests that the jugal extended along the posterodorsal
margin of the postorbital. The infratemporal fenestra is obtuse-angled
anteriorly as suggested by the shape of the posterior margin of the jugal.
Parietals. The
parietals are small (left, Mal-202-2; length = 70 mm;
Figure 4G-H; right,
Mal-202-3; length = 67 mm).
 The supraoccipital and exoccipital articular
surfaces are shallow depressions. The elements suggest that the supratemporal
fenestrae are elongate mediolaterally. Both were found associated with the
basicranium.
The supraoccipital and exoccipital articular
surfaces are shallow depressions. The elements suggest that the supratemporal
fenestrae are elongate mediolaterally. Both were found associated with the
basicranium.
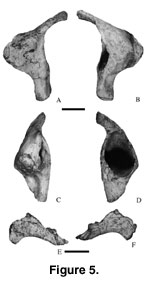
Quadrate. The
right quadrate (Mal-203; Figure 5A-D) is well preserved. The shaft is 160 mm high and
nearly vertical. The head of the quadrate is rounded and is gradually inclined posteriorly. The posterior surface has a tall deep ovoid fossa (height = 70 mm;
width = 50 mm; 45 mm into the pterygoid flange). The medial and lateral walls of
the fossa are 5 mm thick. The fossa is at the dorsoventral midpoint of the
shaft. The pterygoid process is centrally placed on the shaft, thin
mediolaterally, and triangular in lateral view. The articular surface is suboval
in ventral view.
Ectopterygoid.
A left ectopterygoid (Mal-215; Figure 5E-F) is incomplete. It is lunate in
lateral view. The medial surface has a well-developed groove for the palatine
bone while the lateral surface has a well-developed contact surface for the
maxilla.
Basicranium.
The braincase (Mal-202-1; width at paraoccipitals = 140 mm;
Figure 6) is
represented by basioccipital, basisphenoid, exoccipital,
laterosphenoid-orbitosphenoid complex, opisthotic, prootic, and supraoccipital.
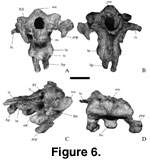 The bones are completely ossified, and their sutures are indistinct. The
pituitary fossa is rounded and large. The canal for the optic nerve (II) is
small, circular, and is directed anterolaterally. Posterior to the canal for the
optic nerve, the rounded canal for the oculomotor nerve (III), and the trochlear
canal for the trochlear nerve (IV) are found in sequence. The opening for the
trigeminal nerve (V) is large and subcircular. A small notch on the
laterosphenoid margin of the trigeminal canal probably carried the ophthalmic
branch of the trigeminal nerve. Anterior and posterior grooves exit ventral to
the canal for the trigeminal nerve. The anterior groove probably corresponds to
the maxillary branch whereas the posterior groove corresponds to the mandibular
branch of the nerve. Posterior to the canal for the trigeminal nerve is a small
dorsoventrally elongate canal for the facial nerve (VII). In Camarasaurus,
the trigeminal nerve and facial nerve openings are on the
basisphenoid-laterosphenoid boundary (Madsen et al. 1995). Thus, these canals
are assumed in Malawisaurus as delineating the
basisphenoid-laterosphenoid boundary. The laterosphenoid is flat and projects
laterally from the braincase wall. Ventral to the fenestra ovalis, in the middle
ear region, is the metotic foramen (called the jugular foramen by
Berman and
McIntosh 1978) for cranial nerves IX to XI (glossopharyngeal, vagus, and spinal
accessory nerves), and probably the jugular vein. The metotic foramen is
mediolaterally narrow and elongate. The hypoglossal canal for cranial nerve XII
is mediolaterally elongate. The foramen magnum is ovoid and higher (28 mm) than
wide (20 mm). It is slightly larger than the occipital condyle. The occipital
condyle is hemispherical but concave dorsally.
The bones are completely ossified, and their sutures are indistinct. The
pituitary fossa is rounded and large. The canal for the optic nerve (II) is
small, circular, and is directed anterolaterally. Posterior to the canal for the
optic nerve, the rounded canal for the oculomotor nerve (III), and the trochlear
canal for the trochlear nerve (IV) are found in sequence. The opening for the
trigeminal nerve (V) is large and subcircular. A small notch on the
laterosphenoid margin of the trigeminal canal probably carried the ophthalmic
branch of the trigeminal nerve. Anterior and posterior grooves exit ventral to
the canal for the trigeminal nerve. The anterior groove probably corresponds to
the maxillary branch whereas the posterior groove corresponds to the mandibular
branch of the nerve. Posterior to the canal for the trigeminal nerve is a small
dorsoventrally elongate canal for the facial nerve (VII). In Camarasaurus,
the trigeminal nerve and facial nerve openings are on the
basisphenoid-laterosphenoid boundary (Madsen et al. 1995). Thus, these canals
are assumed in Malawisaurus as delineating the
basisphenoid-laterosphenoid boundary. The laterosphenoid is flat and projects
laterally from the braincase wall. Ventral to the fenestra ovalis, in the middle
ear region, is the metotic foramen (called the jugular foramen by
Berman and
McIntosh 1978) for cranial nerves IX to XI (glossopharyngeal, vagus, and spinal
accessory nerves), and probably the jugular vein. The metotic foramen is
mediolaterally narrow and elongate. The hypoglossal canal for cranial nerve XII
is mediolaterally elongate. The foramen magnum is ovoid and higher (28 mm) than
wide (20 mm). It is slightly larger than the occipital condyle. The occipital
condyle is hemispherical but concave dorsally.
With the basicranium
(Mal-202-1) oriented with the supraoccipital vertical, which is considered the
normal orientation (Salgado and Calvo 1997), the occipital condyle projects and
faces posteroventrally, the paroccipital processes project anteroventrally, and
the basipterygoid processes project ventrally. The paroccipital processes are
long, anteroposteriorly flattened wing-like structures that curve ventrally,
typical of titanosaurians (Powell 1986;
Chatterjee and Rudra 1996).
Posterolaterally, the paroccipital process has a ventrally directed depression
for the quadrate articulation. The basal tubera are large and separated by a
wide shallow depression.
Salgado and Calvo (1992) interpret the fusion of the
basal tubera in Amargasaurus (MACN-15) as reflecting rigid attachment for
vertebral muscles. By inference, the separation of the basal tubera would
reflect a less extensive muscle attachment in Mal-202-1. The opening for the
internal carotid artery is at the base of the basipterygoid process. The
basipterygoid processes are short (25 mm long) and divergent. In sauropods,
short basipterygoid processes are associated with vertical quadrates, whereas
long basipterygoid processes are associated with highly anteriorly inclined
quadrates (Chatterjee and Rudra 1996).
 Dentary. The
right dentary of Malawisaurus (Mal-174;
Figure 7A-C) was briefly
described by
Jacobs et al. (1993, figure 1b, c). The preserved portion of the dentary is 252 mm long from the symphysis to the surangular notch. The dentary
has a minimum of 15 tooth positions, including unerupted teeth in place. The
tooth row is 159 mm long extending more than half the length of the dentary. The
posteriormost tooth position is 23 mm anterior to the surangular notch.
Replacement teeth are visible through three of the nutrient foramina. The
rostral portion of the dentary appears linear due to crushing.
Dentary. The
right dentary of Malawisaurus (Mal-174;
Figure 7A-C) was briefly
described by
Jacobs et al. (1993, figure 1b, c). The preserved portion of the dentary is 252 mm long from the symphysis to the surangular notch. The dentary
has a minimum of 15 tooth positions, including unerupted teeth in place. The
tooth row is 159 mm long extending more than half the length of the dentary. The
posteriormost tooth position is 23 mm anterior to the surangular notch.
Replacement teeth are visible through three of the nutrient foramina. The
rostral portion of the dentary appears linear due to crushing.
The dentary is slender
and 50 mm deep. The surangular notch is triangular with its apex pointing
anteriorly. The notch divides the dentary posteriorly into a dorsal ramus and a
ventral ramus. The dorsal ramus is small, short, and triangular in lateral view
whereas the ventral ramus is large, long, and quadrangular. The splenial groove
on the medial surface is broadly open posteriorly, tapering forward to close
approximately below the eighth tooth position (Jacobs et al. 1993).
Splenial. The
element (Mal-284; Figure 7D-E) identified as splenial is thin (5 mm wide). A
splenial foramen occurs on the anterior half of the element as in
Mamenchisaurus (Russell and Zheng 1993). The posterior half of the element
is separated into a long, slender ventral ramus and a short, broad dorsal ramus.
Teeth.
Jacobs
et al. (1993, figure 1d) described teeth of Malawisaurus briefly. Some
teeth are present in the jaws (Mal-6, Mal-106, and Mal-174), and others are
isolated (Figure 7F-I; mesiodistal width, range = 06 to 09 mm; average
mesiodistal width = 7 mm; labiolingual widths, range = 4 to 5 mm; average
labiolingual width = 5 mm). The roots of the teeth are nearly cylindrical and
circular in cross section. The crowns are nearly cylindrical at the base and
become lingually flattened towards the apex so that the crown is less convex
lingually than labially. As the crowns become flattened, faint distal and mesial
ridges emerge. The crowns are broadest close to the tip of the teeth. The
surface is rugose. Anterior teeth in the premaxilla and the dentary are similar
in size but broader than more posterior teeth in the maxilla and dentary. The
maxillary teeth (Mal-106) curve anterolingually. Compared with the premaxillary
teeth, the maxillary teeth of Malawisaurus are mesiodistally narrower and
smaller. The maxillary teeth are more cylindrical than the premaxillary teeth
and closely resemble teeth generally associated with titanosaurians (Sanz 1985;
Powell 1986;
Lucas and Hunt 1989;
Scuitto and Martinez 1994; but see
Kues et al.
1980).
Isolated teeth are
assigned to Malawisaurus, because they have more convex labial surfaces
than lingual surfaces, faint distal and mesial ridges, and crowns that are
broader toward the apices of the crowns, and in all respects are similar to the
teeth in the jaws of Malawisaurus. In having convex labial and flattened
lingual surfaces, the teeth of Malawisaurus are like those of
Nemegtosaurus (Nowinski 1971) and Quaesitosaurus (Kurzanov and
Bannikov 1983). Anteroposterior decrease in tooth size and variability of
curvature between anterior and posterior and between upper and lower teeth are
features that are also present in Camarasaurus (Osborn and Mook 1921;
Carey and Madsen 1972), Nemegtosaurus (Nowinski 1971) and
Quaesitosaurus (Kurzanov and Bannikov 1983). In Mamenchisaurus
(Russell and Zheng 1993), teeth diminish in size as crowns become more
compressed.
