 The Polish sub-fossil chironomids
The Polish sub-fossil chironomids
Isabelle Larocque-Tobler
Article number: 17.1.3A
https://doi.org/10.26879/391
Copyright Palaeontological Association, January 2014
Author biography
Plain-language and multi-lingual abstracts
PDF version
Submission: 3 April 2013. Acceptance: 17 December 2013
{flike id=637}
ABSTRACT
This paper presents some of the chironomid head capsules found in the surface sediment of 50 lakes in northeastern Poland and including taxa found in the sediment (past 1000 years) of Lake Zabinskie. Of the 119 taxa found, 104 are presented here. Many taxa were previously described in Brooks et al. (2007) but some are slightly different. The illustrations and descriptions presented in this paper can be used by chironomists all over the world for comparison with their own material.
Isabelle Larocque-Tobler. L.A.K.E.S Institute, Dreihubelweg 68, 3250 Lyss, Switzerland
and Institute of Geography, University of Bern, Erlachstrasse 9A, 3012 Bern, Switzerland
larocque@giub.unibe.ch
Keywords: Chironomidae; Poland, 50 lakes; Lake Zabinskie
Final citation: Larocque-Tobler, Isabelle. 2014. The Polish sub-fossil chironomids. Palaeontologia Electronica Vol. 17, Issue 1;3A; 28p. https://doi.org/10.26879/391
palaeo-electronica.org/content/2014/637-chironomid-identification
INTRODUCTION
For the past 20 years, sub-fossil chironomids preserved in lake sediments have started to be used in paleolimnology to quantitatively reconstruct various factors such as nutrients (Lotter et al., 1998), macrophytes (Langdon et al., 2010), salinity (Heinrichs et al., 1999), lake depth (Engels and Cwynar, 2011), anoxia (Quinlan et al., 1998) and temperature (Larocque-Tobler et al., 2011). As they are becoming accurate indicators (Brooks et al., 2012), their distribution is being studied in many parts of the world because new training sets are being developed for quantitative reconstruction.
Brooks et al. (2007) presented a complete and comprehensive technical guide of Palearctic Chironomidae larvae. However when working in a new area, such as Poland, illustrating the types associated to a species can be very useful as reference. The taxa in the area might be very similar to the pictures presented in Brooks et al. (2007) but they can also present variations. The goal of the present paper is not to replace the Brooks et al. (2007) guide. The paper presented here should be used as a complement providing a fast and mostly visual identification of the taxa found in Polish lakes. In the author's opinion, it would be very interesting if chironomists working in a part of the world not included in the Brooks et al. (2007) guide would present a visual guide of the chironomids found as presented here and in Larocque and Rolland (2006). This exercise is time-consuming, it is difficult to find head capsules which are well preserved and positioned appropriately on the slide to make pictures, and taxonomic aspects used for many years might need to be revised to either fit the Brooks et al. (2007) nomenclature or better described to explain how different the specimen are. However this work is very valuable for chironomists around the world and definitely worth the effort as one always need to confirm his/her identifications.
PARATAXONOMY
Of the whole chironomid larva, only the head capsule made of chitin preserves in the sediment while the rest of the body is decomposed. To differentiate the taxa, only the preserved parts of the head capsules can be used, and often parts which can easily detach (e.g., mandible, antenna) are not present. Thus, chironomists use a parataxonomy to separate taxa based on the few preserved attributes of the head capsules.
This parataxonomy, firstly based on guides of whole larvae (Cranston et al., 1983; Wiederholm, 1983) has been developed by many chironomists throughout the years. Beginners should be aware that the parataxonomy presented in Brooks et al. (2007) and used by most subfossil chironomists has been developed by scientists dedicated to the fossil work and generous enough to present their work at many workshops. The author presents some of the history based on her experience in learning parataxonomy since 1999 at different chironomid workshops. Ian Walker (UBC in Kelowna, Canada) was one of the pioneers who presented a parataxonomy available early on the web. In 1999, Stephen Brooks (Natural History Museum, London) presented an unpublished key separating the Tanytarsini based on the position and number of teeth on the mandible plus the presence/absence of a spur (and its shape) on the antennal pedestal. In 2000, Maria Rieradevall and Stephen Brooks (2001) presented the use of cephalic setation to separate Tanypodinae. These two keys were later incorporated in the key of Brooks et al. (2007) although some of the Tanytarsini types have been renamed based on extensive work performed by Oliver Heiri (University of Bern, Switzerland), Endre Willassen (University of Bergen, Norway) and Torbjørn Ekrem (University of Trondheim, Norway) on living Tanytarsini larvae. In a workshop in 2002, Klaus Brodersen (University of Copenhagen, Denmark (at that time)) distributed an unpublished key to differentiate Chironominae based on the shape of their plates which the author still uses.
This short history is important for beginners to understand that the parataxonomy presented in Brooks et al. (2007) emerged and evolved through time. With more chironomists working in different parts of the world, this parataxonomy will possibly further evolve thus illustrating the need to present papers on parataxonomy of head capsules found in new study areas.
Most importantly, subfossil chironomists should always remember that the word "type" following a species name is to mention that the head capsule resembles this type but it does not imply that this is this species. The "type" can include many species which can have slight differences. One of the best examples is the use of the name Chironomus anthracinus-type. In Québec (Canada) this type includes eight different species, two which could be partly differentiated by the number of teeth on the mandible (Proulx and Hare, submitted), however the mandible is not always present. The other species cannot be differentiated only with the head capsule/mandibles but they possibly all have the 4th lateral tooth on the mentum reduced at different levels. The ecology of these eight species varies from oligotrophic to eutrophic preferences. Thus, it is important when working in a new area, as in Poland, to illustrate what the types associated to a species name look like.
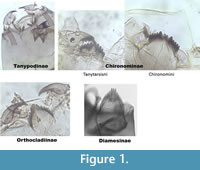 The taxa presented here are divided into sections, representing five of the subfamilies: Tanypodinae, Chironominae, Orthocladiinae, Diamesinae (Figure 1) and Prodiamesinae. The Tanypodinae are elongated or broad head capsules which the author describe as "sleeping bags." They do not have a mentum. The Chironominae include here two different genera: the Tanytarsini and the Chironomini, which can be separated by the shape of ventromental plates. The Tanytarsini have (generally) ventromental plates which are straight and horizontal. The Chironomini have ventromental plates which are longer, wider and often curved. The Orthocladiinae have very narrow ventromental plates, generally on the vertical plan. The Diamesinae generally have more than seven lateral teeth on their mentum.
The taxa presented here are divided into sections, representing five of the subfamilies: Tanypodinae, Chironominae, Orthocladiinae, Diamesinae (Figure 1) and Prodiamesinae. The Tanypodinae are elongated or broad head capsules which the author describe as "sleeping bags." They do not have a mentum. The Chironominae include here two different genera: the Tanytarsini and the Chironomini, which can be separated by the shape of ventromental plates. The Tanytarsini have (generally) ventromental plates which are straight and horizontal. The Chironomini have ventromental plates which are longer, wider and often curved. The Orthocladiinae have very narrow ventromental plates, generally on the vertical plan. The Diamesinae generally have more than seven lateral teeth on their mentum.
The taxa illustrated were found either in one or many lakes of a 50-lake Polish training set (Larocque-Tobler et al., in prep) or in the sediment core of Lake Zabinskie ((54°07'54.5'' N; 21°59'01.1'' E; 120 m a. s. l.), which was analyzed at high resolution (annual to 12 years) for the past 1000 years. This study (training set and analysis of Lake Zabinskie) was part of the CLIMPOL project (www.climpol.ug.edu.pl/pages/idea/general-information.php). Some illustrations are also used to compare different types of head capsules which are similar and can easily be misidentified (e.g., the different types of Heterotrissocladius are also presented in one figure to help differentiation).
SURFACE SAMPLES
The surface sediment of 50 lakes, including Lake Zabinskie, was sampled in summer-autumn of 2011- 2012. A list of these lakes with some of their characteristics is found in Table 1. Most lakes were anoxic (i.e., oxygen level < 4 mg/l in the hypolimnion). Lake depth varied from 3 to 41 m and water temperature at the bottom varied accordingly.
The number of taxa found in the 50 surface sediment samples was 112. The most abundant (i.e., found in the highest number of lakes) are Tanytarsus mendax-type, Tanytarsus no spur on the antenna pedestal, Polypedilum nubeculosum-type, Chironomus anthracinus-type, Dicrotendipes nervosus-type, Cladotanytarsus mancus-type and Microtendipes pedellus-type.
SAMPLES LAKE ZABINSKIE
Lake Zabinskie is a varved lake and the chironomid preserved in its sediment has been used to reconstruct temperature at annual resolution until the 19th century, then at a 6/12-year resolution for the last 1000 years. In those samples, a few taxa have been found which are not in the surface samples or have been found only in one lake at very low percentages (Cryptotendipes, Paratendipes albimanus-type, Micropsectra insignilobus-type, Heterotrissocladius marcidus-type, Heterotrissocladius subpilosus-type, Prodiamesa, Paramerina and Zavrelimyia).
TANYPODINAE
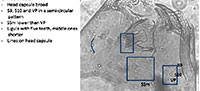
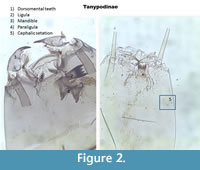 The Tanypodinae are very distinctive from other families. They do not have ventromental plates, but have a ligula, paraligula and some have dorsomental teeth (Figure 2). Before 2005, different types of Tanypodinae were not separated in many of the transfer functions. In Larocque et al. (2001), two categories were used: Procladius and Pentaneurini. The Procladius group included all taxa with a wider head capsule and mandible with dark tips (Figure 2). All other Tanypodinae were classified as Pentaneurini. Since then, Rieradevall and Brooks (2001) developed a taxonomy based on the position cephalic setation (Figure 2). Note that the ventromental pore (VP) is recognizable by its elongated shape and its double lines.
The Tanypodinae are very distinctive from other families. They do not have ventromental plates, but have a ligula, paraligula and some have dorsomental teeth (Figure 2). Before 2005, different types of Tanypodinae were not separated in many of the transfer functions. In Larocque et al. (2001), two categories were used: Procladius and Pentaneurini. The Procladius group included all taxa with a wider head capsule and mandible with dark tips (Figure 2). All other Tanypodinae were classified as Pentaneurini. Since then, Rieradevall and Brooks (2001) developed a taxonomy based on the position cephalic setation (Figure 2). Note that the ventromental pore (VP) is recognizable by its elongated shape and its double lines.
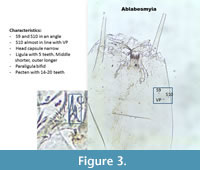
Ablabesmyia (Figure 3) Seta 9 and Seta 10 are in an angle. S10 can be almost aligned with the ventromental pore (VP). If present, the ligula has five teeth with the central one shorter and the outer ones longer. The paraligula is bifid. The pecten hypopharyngis (referred later as pecten) has 14-20 teeth of unequal length.
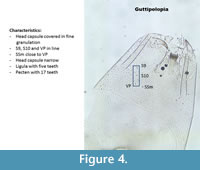
Guttipelopia (Figure 4) is granulated all over the head capsule. This is the only Tanypodinae with this characteristic thus Guttipelopia is easily recognizable. The ligula has five teeth. In Brooks et al. (2007) the teeth are described as follows: with the central one shorter and the outer ones longer. However, in many head capsules from Poland, the teeth have almost all the same length (Figure 4). The pecten has 17 teeth. S9, S10 and VP are almost in one horizontal angle.
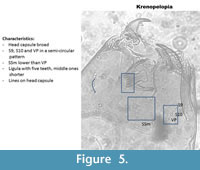
Krenopelopia (Figure 5) has its ventromental setae (S9, S10, VP) in a semi-circular pattern. The ligula has five teeth with the central one shorter and the outer ones longer. The paraligula is paired. The pecten has ten teeth. In the Polish samples, the head capsules seem to have vertical striation. This is not described in Brooks et al. (2007) but seems to be consistent in the head capsules identified.
 |
 |
 |
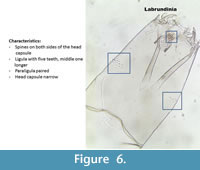
Labrundinia (Figure 6) is very easy to recognize because it has spines on both sides of the head capsule. The ligula has five teeth and the middle one is longer. The paraligula is paired. The pecten has seven teeth of equal length.
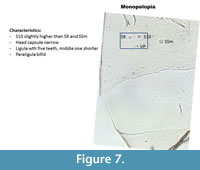
Monopelopia (Figure 7) has a narrow head capsule. S9 and S10 are almost aligned but S10 is slightly higher. SSm and VP are almost aligned. This illustration follows Blakely et al. (2010).
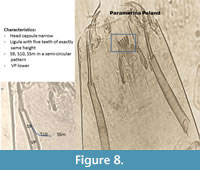
Paramerina (Figure 8) has a ligula with five teeth of exactly the same length. The paraligula is bifid. S9-S10 and SSm are in a curve.
 |
 |
 |
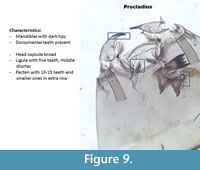
Procladius (Figure 9) has dorsomental teeth. The head capsule is very broad. The mandible has very dark tips. The pecten has 10-15 teeth with a second row of smaller teeth.
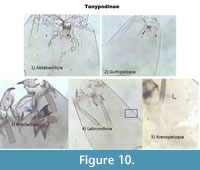
Figure 10 places all Tanypodinae together and illustrates their major differences.
 |
 |
CHIRONOMINAE - TANYTARSINI
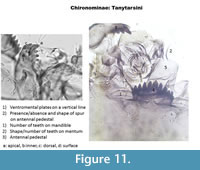
This is the subfamily where the most advances in parataxonomy have been made since 2000. The Tanytarsini have ventromental plates which are horizontal, often with striation (Figure 1,Figure 11). Types can be separated using the presence/absence and shape of a spur on the antennal pedestal, by the shape and number of teeth on the mentum and by the number of teeth on the mandible (Figure 11).
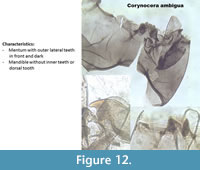
Corynocera ambigua (Figure 12) is unique and easily recognizable with two dark median teeth in front of the mentum and three paler teeth on the back. The mandibles do not have dorsal or inner teeth.
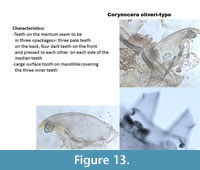
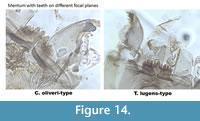
Corynocera oliveri-type (Figure 13) has teeth on the mentum which look like they are in three packages: the three median are paler higher and seem to be on a backward focal plane. There are four darker teeth on each side of the median which are pressed together and seem to be slightly forward. The mandible has three inner teeth, two dorsal teeth and one very large surface tooth covering the inner teeth. Tanytarsus lugens-type (Figure 14) also has a large dorsal tooth on the mandible which can as well cover the inner teeth. However, all the teeth on the mentum are on the same focal plane (Figure 14) sometimes the median tooth is slightly elevated.
 |
 |
 |
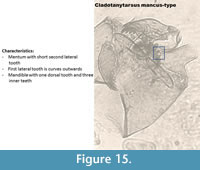
Cladotanytarsus mancus-type (Figure 15) has five lateral teeth on its mentum and the second lateral tooth is smaller. Brooks et al. (2007) described the mandible has having three inner teeth and one dorsal tooth.
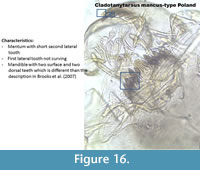
Cladotanytarsus mancus-type Poland (Figure 16). In Poland, specimen had two dorsal teeth and one surface tooth as illustrated in Figure 16 thus we named it Cladotanytarsus mancus-type Poland.
 |
 |
 |
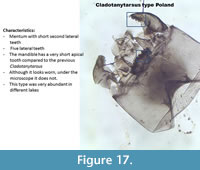
Cladotanytarsus type Poland (Figure 17) also has a very short second lateral tooth but the mentum and the mandible seem worn (teeth on the mentum and mandibles rounder). However, too many of these Cladotanytarsus type were found in different lakes and in the sediment of Lake Zabinskie looking exactly the same thus it cannot be all worn specimen.
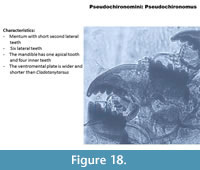
Pseudochironomus (Figure 18) has six lateral teeth on the mentum. The second lateral tooth is very short and slightly pressed to the first lateral tooth. The ventromental plate is shorter than the Tanytarsini. Note that Pseudochironomus is not a Tanytarsini but a Pseudochironomini. The mandible has one apical tooth and four inner teeth.
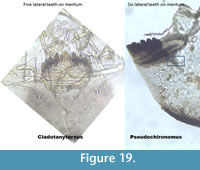
Cladotanytarsus and Pseudochironomus (Figure 19) resemble each other and can at first be misidentified when half capsules are found. The last tooth on Pseudochironomus can be very small looking like the mentum has only five teeth. However, Pseudochironomus has a slender, triangular structure below the ventromental plate.
 |
 |
 |
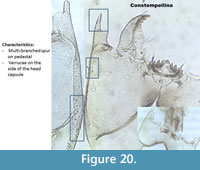
Constempellina (Figure 20) has a long spur on the antennal pedestal, sometimes multi-branched and has verrucae on the head capsules. The ventromental plates are wider than longer, almost like a Chironomini (Figure 1). There are five lateral teeth which sometimes seem a bit curved.
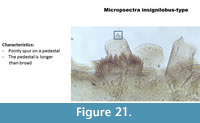
Micropsectra insignilobus-type (Figure 21) has a medium-size pointy spur on a pedestal longer than broad. The mentum is typical of Tanytarsini with five lateral teeth and one median tooth.
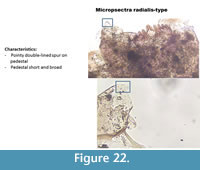
Micropsectra radialis-type (Figure 22) has a short and pointy spur on the antenna pedestal. The pedestal is short and broad. The spur is "double-lined." The mentum is typical of Tanytarsini with five lateral teeth and one median tooth.
 |
 |
 |
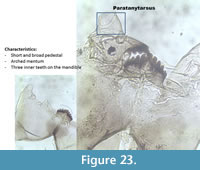
Paratanytarsus (Figure 23) has an arched mentum due to the median tooth and the first lateral teeth on the mentum which are prominent. When the mandible is present, types can be distinguished: P.austriacus-type has three inner teeth on the mandible, P. penicillatus-type has two inner teeth on the mandible while P. type A has a vestigial third inner tooth on the mandible and two dorsal teeth.
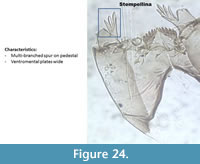
Stempellina (Figure 24) has a unique multi-branched spur on the pedestal. The ventromental plates are wider than longer, looking more like the Chironomini (Figure 1). It has six lateral teeth. Brooks et al. (2007) described the lateral teeth slightly shorter than the median tooth, but this is not visible in the head capsule illustrated here, or in any of those identified in the Polish surface sediment.
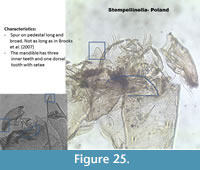
Stempellinella Poland (Figure 25) has a large and long spur on the pedestal. The ventromental plates are wider than longer, looking more like a Chironomini (Figure 1). It has six lateral teeth and one median on the mentum, sometimes with a supplementary tooth. Brooks et al. (2007) describe a mandible with three inner teeth. The specimen in Poland has a mandible with a dorsal tooth and setae on the dorsal tooth. The spur on the antenna is also much longer in Brooks et al. (2007).
 |
 |
 |
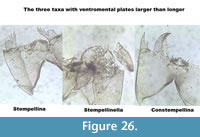
The three taxa with ventromental plates wider than longer are illustrated in Figure 26.
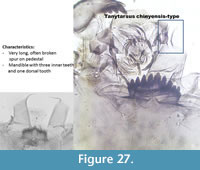
Tanytarsus chinyensis-type (Figure 27) has a very long spur on the pedestal which is often broken or folded. It has five lateral teeth and one median tooth on the mentum. The mandible has three inner teeth.
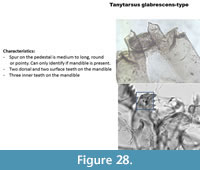
Tanytarsus glabrescens-type (Figure 28) has a mandible with three inner teeth, two surface and two dorsal teeth. The pedestal has a moderately long spur rounded at the tip. In Brooks et al. (2007) this type is described as having the first lateral teeth on the mentum curved-outward, which is not the case in the Polish specimen. However, the mandible does correspond to the description of Brooks et al. (2007) for glabrescens-type.
 |
 |
 |
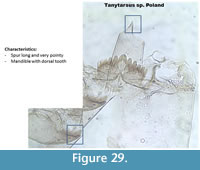
Tanytarsus type Poland (Figure 29) has a very long and pointy spur on the pedestal. The mandible has no dorsal or surface teeth.
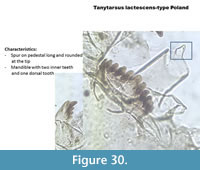
Tanytarsus lactescens-type Poland (Figure 30) has a medium-size rounded spur on the pedestal. The mandible has two inner teeth and one dorsal tooth. The mentum is typical with one median and five lateral teeth all on the same focal plane.
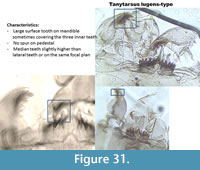
Tanytarsus lugens-type (Figure 31) has a large dorsal tooth on the mandible sometimes covering the three inner teeth. The mandible also has two dorsal teeth. The pedestal has no spur. The mentum has five lateral teeth and one median tooth. The median and the lateral teeth are (sometimes) not on the same focal plan than the other lateral teeth (Figure 30, left picture). This is not always the case in the Polish samples (two other pictures).
 |
 |
 |
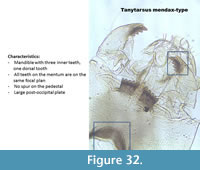
Tanytarsus mendax-type (Figure 32) has a mandible with three inner teeth, one dorsal tooth and no surface tooth. The pedestal has no spur. It has five lateral teeth and one median tooth all on the same focal plan.
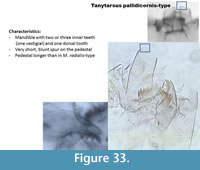
Tanytarsus pallidicornis-type (Figure 33) has a short blunt spur on the pedestal. The mandible has two or three inner teeth and one dorsal tooth.
Tanytarsus special Poland (Figure 34) is not described in Brooks et al. (2007). It has ventromental plates elongated like other Tanytarsus but they are slightly curved at the tip. The second lateral teeth on the mentum are thinner than the four other lateral teeth. The mandible has two inner (one vestigial) teeth and one dorsal tooth. The taxon has been found in a few samples of Lake Zabinskie.
 |
 |
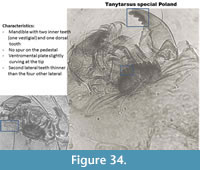 |
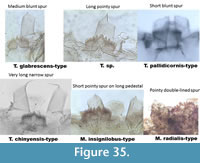
Figure 35 illustrates the types with spur to make visual summary of differences.
 |
CHIRONOMINAE- CHIRONOMINI
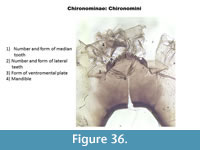
The shape of the ventromental plates and the shape/number of teeth on the mentum are often enough to separate the different taxa of Chironomini (Figure 36).
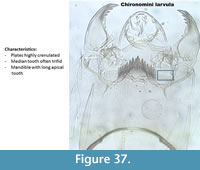
Chironomini (Figure 37) illustrates younger instars of one of the Chironomini taxa, called a larvula. In Brooks et al. (2007) it is written "probably Chironomus" however this larvula was often found in samples with no 4th instar of Chironomus but other Chironomini such as Parachironomus. The larvula is here illustrated because it was found in quite high numbers in many samples.
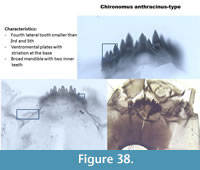
Chironomus anthracinus-type (Figure 38) has six lateral teeth on the mentum with the 4th lateral shorter than the 3rd and the 5th. The median is trifid, which is distinctive for all Chironomus. The ventromental plates have striations at the base. The mandible is broad with two or three inner teeth.
 |
 |
 |
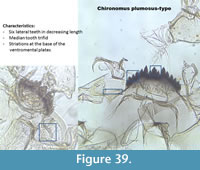
Chironomus plumosus-type (Figure 39) has six lateral teeth on the mentum in decreasing length from the middle to the end of the mentum. The median is trifid. The mandible has three inner teeth. Striations are present at the base of the ventromental plates.
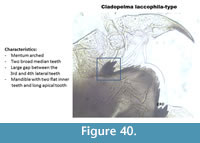
Cladopelma laccophilia-type (Figure 40) has an arched mentum with two median teeth. It has six lateral teeth on the mentum, and there is a large gap between the 3rd and 4th lateral teeth. The last lateral tooth is very small. The inner teeth on the mandible are flat. The apical tooth is very long.
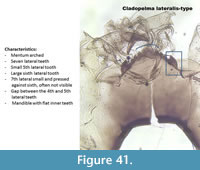
Cladopelma lateralis-type (Figure 41) has a mentum with six lateral teeth. There is a gap between the 4th and the 5th lateral teeth.
 |
 |
 |
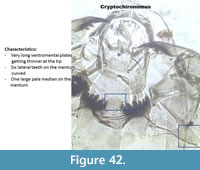
Cryptochironomus (Figure 42) has one rounded and pale median tooth on the mentum. The six lateral teeth on the mentum are dark and curved. The mandible has two inner teeth and a very long apical tooth.
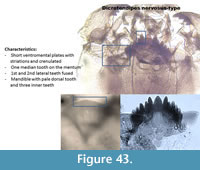
Dicrotendipes nervosus-type (Figure 43) has narrow striation and crenulated ventromental plates. The mentum has one rounded median tooth and six lateral teeth. The first and second lateral teeth are often fused. Sometimes the second lateral is smaller and pressed to the first lateral tooth. The mandible has three inner teeth, a very long apical tooth and a pale dorsal tooth. Smaller teeth can be attached to the dorsal tooth.
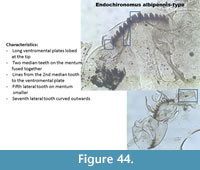
Endochironomus albipennis-type (Figure 44) has a mentum with four median teeth and the two middle ones are fused together. The fifth lateral is smaller than the fourth and sixth6 teeth. The seventh lateral tooth on the mentum is curved outside. The ventromental plates are lobed at the end. The mandible has three inner teeth.
 |
 |
 |
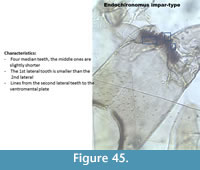
Endochironomus impar-type (Figure 45) has a mentum with four median teeth, and the middle ones are slightly shorter than the outer ones. The first lateral tooth is smaller. There are six lateral teeth on the mentum. The mandible has four inner teeth.
Endochironomus tendens-type (Figure 46) has a mentum with three median and six lateral teeth. The first lateral tooth is the same size as the second lateral tooth. The mandible has three inner teeth.
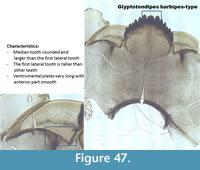
Glyptotendipes barbipes-type (Figure 47) has a mentum with a rounded median tooth which is wider than the first lateral tooth. It has six lateral teeth on the mentum. The first lateral is generally taller than the other five lateral teeth. The mandible has three inner teeth with a pale dorsal tooth. The anterior part of the ventromental plate is smooth.
 |
 |
 |
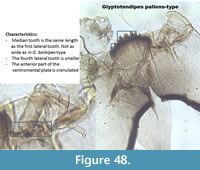
Glyptotendipes pallens-type (Figure 48) has a mentum less arched than G. barbipes-type. The median is the same length as the first lateral tooth. The mandible has three inner teeth with a pale dorsal tooth. The anterior part of the ventromental plate is crenulated.
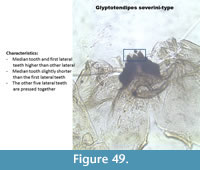
Glyptotendipes severini-type (Figure 49) has a mentum with a median slightly shorter than the first lateral teeth. The five other lateral teeth are pressed together giving the impression of three median teeth. The mandible has three inner teeth with the middle one shorter than the other two. The margin of the mandible has small striation.
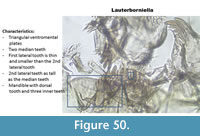
Lauterborniella (Figure 50). The most prominent characteristic is the triangular ventromental plates. The mentum has two median teeth. The first lateral tooth is thin and small. The second lateral is as tall as the median. The mandible has a short apical tooth, a large dorsal tooth and two inner teeth.
 |
 |
 |
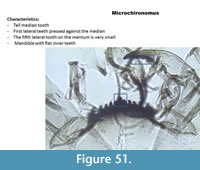
Microchironomus (Figure 51) has a mentum with one median tooth and seven lateral teeth. The first lateral tooth is smaller than the median. The fifth and seventh lateral teeth are very small. The mandible has a long apical tooth and two inner teeth which look almost square.
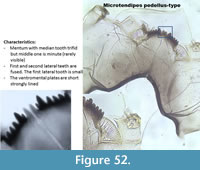
Microtendipes pedellus-type (Figure 52) has a mentum with two pale median teeth and six lateral teeth. The first lateral tooth is small and fused to the second lateral tooth. The mandible has a short apical tooth, three inner teeth and one dorsal tooth.
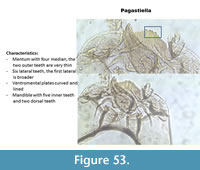
Pagastiella (Figure 53) has very long and curved ventromental plates. The mentum has four median teeth and six lateral teeth. The second median tooth is very thin. The mandible has five inner teeth and two dorsal teeth.
 |
 |
 |
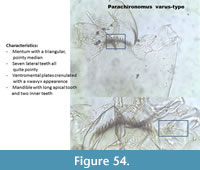
Parachironomus varus-type (Figure 54) has a mentum with a well-defined pointy median and seven lateral teeth. The ventromental plates are "wavy." The mandible has a long apical tooth and two inner teeth.
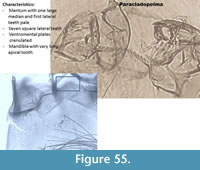
Paracladopelma (Figure 55) has a mentum with a wide and pale median tooth and seven square lateral teeth decreasing in width from the middle to the end of the mentum. The ventromental plates are strongly striated on their whole surface and crenulated. The mandible has a long apical tooth with two or three inner teeth.
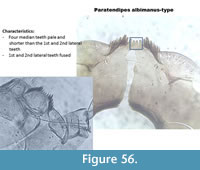
Paratendipes albimanus-type (Figure 56) has a mentum with four pale median teeth. The middle teeth are smaller than the outer two. The first and second lateral teeth are fused. The mandible has two inner teeth, one inner vestigial tooth and one dorsal tooth.
 |
 |
 |
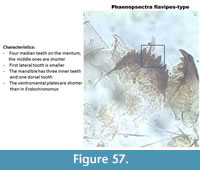
Phaenopsectra flavipes-type (Figure 57) has a mentum with four median teeth and the middle pair is short. It has six lateral teeth but the last two are reduced. The mandible has three inner teeth and the third one is wider than the two middle ones.
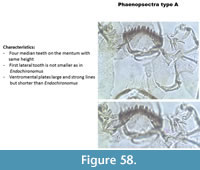
Phaenopsectra type A (Figure 58) has a mentum with four median teeth of equal length. The mentum has six lateral teeth, the outer two are not reduced, as mentioned by Brooks et al. (2007) but not visible in Figure 58. Striations on the ventromental plates are absent in the middle of the plate.
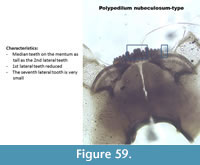
Polypedilum nubeculosum-type (Figure 59) has a mentum with two rounded median teeth. The first lateral tooth is short and the last lateral tooth on the mentum is minute. In the Polish samples, the same configuration of teeth on the mentum has been shown but all teeth were pointy instead of round as illustrated in Figure 60. The mandible had one dorsal tooth and two inner teeth.
 |
 |
 |
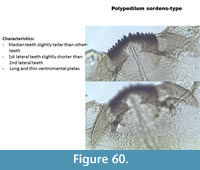
Polypedilum sordens-type (Figure 60) has a mentum with most teeth more or less of equal length. The median are slightly taller than other teeth. The ventromental plates are long and slightly curved. The mandible has three inner teeth.
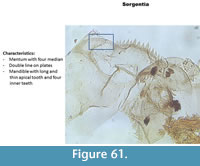
Sergentia (Figure 61) has a mentum with four median teeth. The middle ones are slightly smaller than the outer pair. All four median are taller than the lateral teeth. The ventromental plates have double striations. The mandible has four inner teeth and a long apical tooth.
Stictochironomus rosenschoeldi (Figure 62) has a mentum with four median teeth of which the middle two are smaller than the outer two. The fifth lateral tooth is taller than the fourth and sixth lateral teeth. The ventromental plates have no striation. The mandible has one dorsal tooth and two inner teeth.
 |
 |
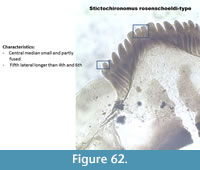 |
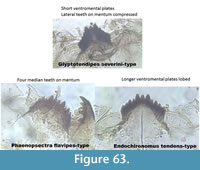
Figure 63 illustrates the differences between the head capsules with three or four median teeth on the mentum and compressed lateral teeth.
 |
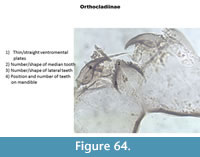
ORTHOCLADIINAE
The shape of the ventromental plates and the number/shape of the teeth on the mentum are used to separate the taxa (Figure 64).
Chaetocladius piger-type (Figure 65) has two pale and broad median teeth on the mentum and five lateral teeth. The ventromental plate is rounded (bulbous) at the tip.
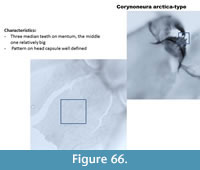
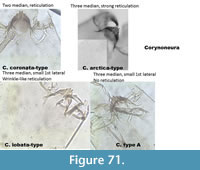
Corynoneura arctica-type (Figure 66) has very thin and long ventromental plates. The mentum has three median teeth with the middle one relatively big (compared to other taxa). There is a well-defined reticulation on the head capsule. The mandible has a short apical tooth, long dorsal tooth and three inner teeth.
 |
 |
 |
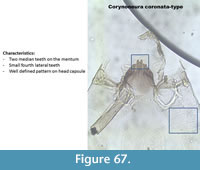
Corynoneura coronata-type (Figure 67) has two rounded median teeth and a defined pattern on the head capsule.
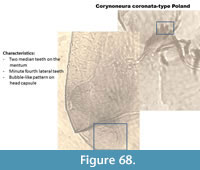
Corynoneura coronata-type Poland (Figure 68) has two median teeth on the mentum and the first lateral teeth are minute. The pattern on the head capsule is very strong and looks like bubbles, which is very different than the pattern of C. coronata-type in Figure 67.
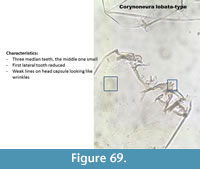
Corynoneura lobata-type (Figure 69) has three median teeth on the mentum with the middle one small. The first lateral tooth is small. It has very weak striation described by Brooks et al. (2007) as "wrinkled sculpturing."
 |
 |
 |
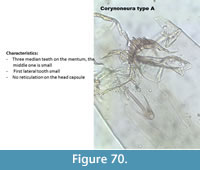
Corynoneura type A (Figure 70) has three median teeth on the mentum with a small middle one. The first lateral tooth is small and thin. There is no pattern on the head capsule.
Figure 71 illustrates the differences between the different types of Corynoneura.
Cricotopus/Orthocladius have not been separated into different morphotypes for a very long time and many transfer functions have them either together or only in Cricotopus and Orthocladius genera. Brooks et al. (2007) mention that this group of genera includes more than 180 species in the Holarctic and that very many variations can occur in the types described below.
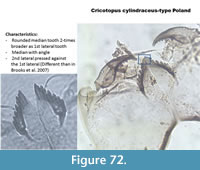
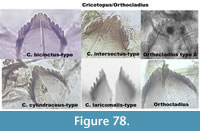
Cricotopuscylindraceus-type Poland (Figure 72) has a mentum with a wide median tooth with angles and the first and second lateral teeth are small. The specimen presented on the right picture has a second lateral pressed against the first lateral, which is not the case in the picture of the half head capsule in Brooks et al. (2007).
 |
 |
 |
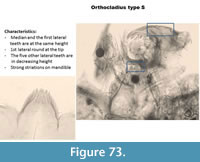
Orthocladius type S (Figure 73) has one rounded median tooth on the mentum. The first lateral teeth are the same height as the median. The five other lateral teeth have heights decreasing from the middle to the end of the mentum. The ventromental plate is typical of Cricotopus. The mandible has strong striations.
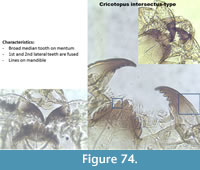
Cricotopus intersectus-type (Figure 74) has a broad median tooth on the mentum. The first and the second lateral teeth are fused.
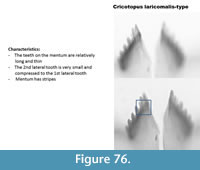
Cricotopus laricomalis-type (Figure 75) has a mentum with long and thin teeth. The second lateral tooth is small and pressed to the first lateral tooth. The example in Brooks et al. (2007) has thinner and longer teeth on the mentum however the presence of stripes on the mentum suggests that this type is "lateralis."
 |
 |
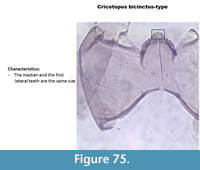 |
Cricotopus bicinctus-type (Figure 76) has a mentum with the median tooth and the first lateral teeth of similar length and width.
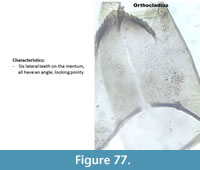
Orthocladius (Figure 77) has a mentum with a broad median tooth and six lateral teeth, which are all flat at the tip, looking kind of rectangular in shape. No picture in Brooks et al. (2007) exactly resembles this type.
Figure 78 summarizes the Cricotopus/Orthocladius and their differences.
 |
 |
 |
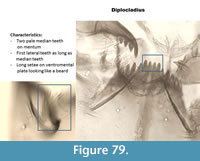
Diplocladius (Figure 79) has very long "beard-like" setae on large ventromental plates. The mentum has two pale median teeth which are of equal length with the first lateral teeth. The mandible has four inner teeth and a very long apical tooth.
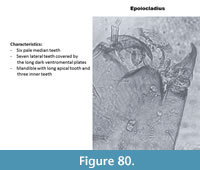
Epoiocladius (Figure 80) has a mentum with six pale median teeth and seven lateral teeth. The ventromental plates are dark and cover the lateral teeth. The mandible has three inner teeth and a long apical tooth.
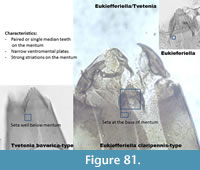
Eukiefferiella/Tvetenia (Figure 81) has strong striations under the mentum. The mentum has single or double median teeth wider than the lateral teeth. The mandible has three inner teeth with a short apical tooth. Spines are present on the margin of the mandible (mola). In Figure 81, three different taxa are present: Eukiefferiella claripennis-type has two median teeth and five lateral teeth. The seta submenti is positioned at the base of the mentum. The position of this seta also identifies the picture on the right corner as Eukiefferiella. The picture on the left corner has a seta much lower on the head capsule suggesting a Tvetenia barvarica-type.
 |
 |
 |
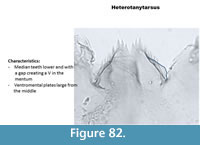
Heterotanytarsus (Figure 82) has a mentum with median and first lateral teeth located below the other six lateral teeth, creating a V shape. The ventromental plate is rounded at the tip. The mandible has three inner teeth and a long apical tooth.
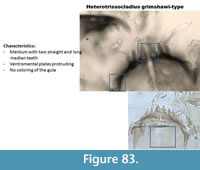
Heterotrissocladius grimshawi-type (Figure 83) has two long and straight median teeth on the mentum. The area under the mentum (gula) is pale. The ventromental plate is protruding at the tip. The mandible has three inner teeth.
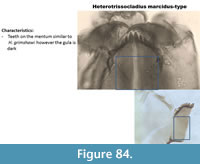
Heterotrissocladius marcidus-type (Figure 84) is similar to H. grimshawi-type with a mentum with two long straight median teeth. However, the area under the mentum (gula) is dark in H. marcidus-type.
 |
 |
 |
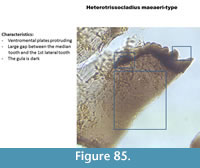
Heterotrissocladius maeaeri-type (Figure 85) has a single broad median tooth on the mentum which appears flat. There is a large gap in a V shape between the median and the first lateral teeth. In the Polish samples, only half head capsules were found and photographed. The picture 8.51 in Brooks et al. (2007) is much more eloquent.
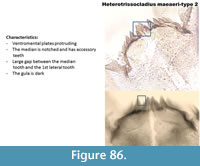
Heterotrissocladius maeaeri-type 2 (Figure 86) has a mentum with a notched median tooth and accessory teeth. There is still a large gap between the median tooth and the first lateral teeth.
Heterotrissocladius subpilosus-type (Figure 87) has a mentum with two broad median teeth with small accessory teeth. The gula is un-pigmented. It can resemble H. maeaeri type-2 but the gap between the median teeth and the first lateral is not present. The gula is also paler.
 |
 |
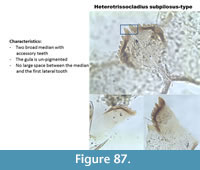 |
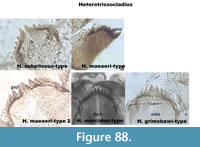
Figure 88 presents the Heterotrissocladius types together.
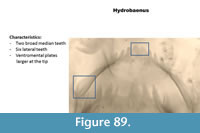
Hydrobaenus (Figure 89) has two broad median teeth on the mentum and six lateral teeth. The ventromental plate is rounded at the tip. The mandible has three inner teeth and a long apical tooth.
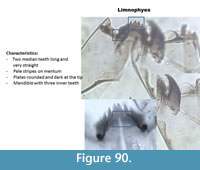
Limnophyes (Figure 90) has two straight and long median teeth on the mentum and five lateral teeth. The mentum has often pale stripes. The ventromental plate is round and dark at the tip. The mandible has three inner teeth and a short apical tooth.
 |
 |
 |
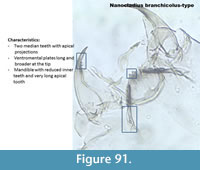
Nanocladius branchicolus-type (Figure 91) has a mentum with two median teeth with apical projections and five lateral teeth. The ventromental plates are long and broader at the tip, similar to Psectrocladius sordidellus-type but they have no seta. The mandible has three reduced inner teeth with a very long apical tooth.
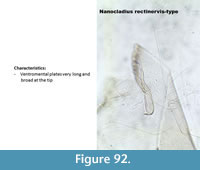
Nanocladius rectinervis-type (Figure 92) has very long and wide ventromental plates. The mandible has three reduced inner teeth with a very long apical tooth.
Paracladius (Figure 93) has a median tooth very broad (half of the mentum) and pale. The mentum has six lateral teeth with a first lateral tooth slightly longer. The ventromental plate is broader at the tip. The mandible has a very long and narrow apical tooth and three inner teeth.
 |
 |
 |
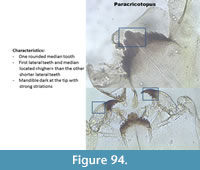
Paracricotopus (Figure 94) has a mentum with a broad median tooth and five lateral teeth. The first lateral teeth are at the same level than the median, the other lateral teeth are lower. The mandible has three inner teeth and a long apical tooth.
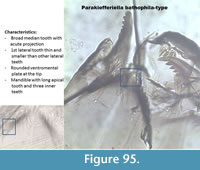
Parakiefferiella bathophila-type (Figure 95) has a mentum with one broad median tooth with apical projection. The first lateral tooth is thin and smaller than the other five lateral teeth. The ventromental plate is rounded at the tip. The mandible has three thin inner teeth and a long apical tooth.
Parakiefferiella triquetra-type (Figure 96) has one broad median tooth with a triangular pale shape. There are six lateral teeth partly covered by the wide ventromental plate. The mandible has three thin inner teeth and a long apical tooth.
 |
 |
 |
Parametriocnemus (Figure 97) has one or two broad median teeth and five lateral teeth. The third and fourth lateral teeth are very small. The ventromental plate is broad and curves outwards at the tip. The mandible has a short apical and three inner teeth.
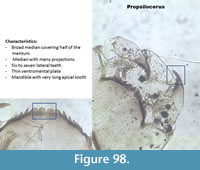
Propsilocerus (Figure 98) has a mentum with broad mentum with projections. There are six to seven lateral teeth. The last lateral tooth is extremely reduced. The ventromental plate is very narrow. The mandible has four inner teeth and the apical tooth is very long and broad.
In the Polish samples, Psectrocladius sordidellus has two distinctive types described below. These two types were found in different lakes, thus it is probably worth to make an effort to separate them.
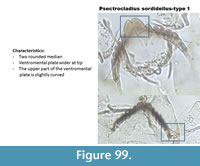
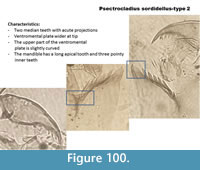
Psectrocladius sordidellus-type 1 (Figure 99) has a mentum with two rounded median teeth. The ventromental plate is wider at the tip. In the Polish samples, there seems to be a distinction between this type and Psectrocladius sordidellus-type 2 (Figure 100), which has also two median teeth but they have apical projections. In both types, the top of the ventromental plate is curved, which helps in distinguishing with half head capsules of P. septentrionalis-type which has a ventromental plate with a straight line at the top.
 |
 |
 |
Psectrocladius calcaratus-type (Figure 101) has one very broad median tooth with an acute projection. The ventromental plates are very long and triangular at the tip. The upper part of the ventromental plate is straight.
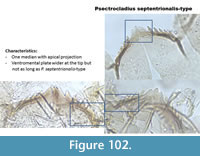
Psectrocladius septentrionalis-type (Figure 102) has one broad median tooth with acute projection. The median is not as broad as in P. calcaratus-type. The ventromental plate is long and triangular at the tip, not as long as in P. calcaratus-type. The upper part of the ventromental plate is straight.
 |
 |
 |
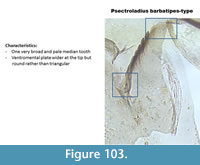
Psectrocladius barbatipes-type (Figure 103) has one broad pale median tooth and lateral teeth are equal in length. The ventromental plates are broad at the tip, not triangular as in P. calcaratus-type or P. septentrionalis-type. The upper part of the ventromental plate is curved but the tip is wider than in P. sordidellus-type.
Figure 104 illustrates all Psectrocladius type together.
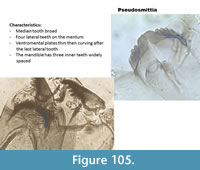
Pseudosmittia (Figure 105) has a mentum with one broad median tooth and four lateral teeth. The ventromental plate is first thin then it curves after the mentum. The mandible has two or three inner teeth with large space in-between.
 |
 |
 |
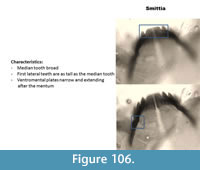
Smittia (Figure 106) has a mentum with a broad median tooth and five lateral teeth. The first lateral teeth are as tall as the median tooth. The ventromental plate is narrow and extends below the mentum. The mandible has three inner teeth and the apical tooth is short.
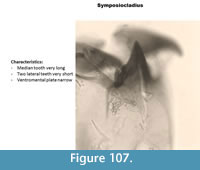
Symposiocladius (Figure 107) has a mentum with one very long median tooth and two very short lateral teeth. The ventromental plate is narrow. The mandible has three short inner teeth and one short apical tooth.
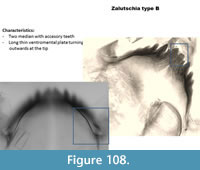
Zalutschia type B (Figure 108) was called lingulata-pauca in many of the earlier training sets. It has a mentum with two median teeth with accessory teeth and six lateral teeth. The ventromental plate is narrow and curving outwards at the tip. Seta can sometimes be seen. The mandible has three inner teeth and a basal inner tooth shorter than the inner teeth.
 |
 |
 |
Zalutschia zalutschicola-type (Figure 109) has a mentum with two broad and pale median teeth and six lateral teeth. The first and the sixth lateral teeth are very small. The ventromental plate is thin becoming wider at the tip. The mandible has three inner teeth and a basal inner tooth shorter than the inner teeth.
 |
DIAMESINAE
The taxa in this family generally have a large number of lateral teeth.
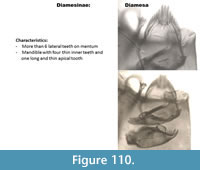
Diamesa (Figure 110) has a mentum with a high number of narrow lateral teeth. The ventromental plates are thin. The mandible has four inner teeth that are long and thin.
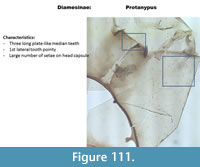
Protanypus (Figure 111) has a mentum with three plate-like pale median and two pointed first lateral teeth. The mandible has five small inner teeth and one long apical tooth. The head capsule has many setae.
 |
 |
PRODIAMESINAE
Monodiamesa (Figure 112) has a mentum with a broad tooth indented in a U shape. Six lateral teeth are decreasing in size. The ventromental plate is long wide and extends beyond the mentum. The mandible has a long apical tooth and two small inner teeth.
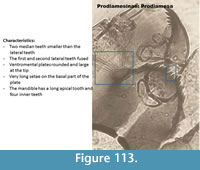
Prodiamesa (Figure 113) has a mentum with the median teeth located lower than the first lateral teeth. The first and second lateral teeth are fused. The ventromental plate is round and large at the base. It has very long setae. The mandible has a long apical tooth and four inner teeth.
 |
 |
ACKNOWLEDGMENTS
This work was possible due to a great sampling team composed of Dr. W, Tylmann, Dr. C. Kamenik, Prof. Dr. M. Grosjean, Dr. I. Hernandez-Almeida, A. Bonk, and B, Amann.This project was part of the CLIMPOL program (www.climpol.ug.edu.pl) funded by the Swiss Contribution to the enlarged European Union, Project No. PSPB-086/2010.
REFERENCES
Blakely, T.J., Cranston, P.S., and Winterbourn, M.J. 2010. Container-inhabiting Monopelopia larvae newly recorded in New Zealand. New Zealand Entomologist, 33:38-42.
Brooks, S.J., Langdon, P.G., and Heiri, O. 2007. The Identification and Use of Palaearctic Chironomidae Larvae in Palaeoecology. Quaternary Research Association, Technical Guide no. 10, London.
Brooks,S.J., Axford,Y., Heiri,O., Langdon,P.G., and Larocque-Tobler, I. 2012. Chironomids can be reliable proxies for Holocene temperatures. A comment on Velleet al., 2010. The Holocene, 22:1482-1494.
Cranston, P.S., Oliver, D.R., and Saether, O.A. 1983. The larvae of Ortocladiinae (Diptera: Chironomidae) of the Holarctic region. Keys and diagnoses. Entomol. Scand. Suppl. 19: 149-291.
Engels, S. and Cwynar, L.C. 2011. Changes in fossil chironomid remains along a depth gradient: evidence for common faunal thresholds within lakes. Hydrobiologia, 665:15-25.
Heinrichs, M.L., Walker, I.R., Mathewes, R.W., and Hebda, R.J. 1999.Holocene chironomid-inferred salinity and paleovegetation reconstruction from Kilpoola Lake, British Columbia. Géographie physique et Quaternaire, 53:211-221.
Langdon, P.G., Ruiz, Z., Wynne, S., Sayer, C.D., and Davidson, T.A. 2010. Ecological influences on larval chironomid communities in shallow lakes: implications for palaeolimnological interpretations. Freshwater Biology, 55:531-545.
Larocque, I., Hall, R.I., and Grahn, E. 2001. Chironomids as indicators of climatic and environmental change: A 100-lake training set from a subarctic region of northern Sweden (Lapland). Journal of Paleolimnology, 26:307-322.
Larocque-Tobler, I., Quinlan, R., Müller Stewart, M., and Grosjean, M., 2011. Chironomid-inferred temperature changes of the last century in meromictic Lake Seebergsee, Switzerland: assessment of two calibration methods. Quaternary Science Reviews, 30:1770-1779.
Larocque, I. and Rolland, N. 2006. Le guide visuel des chironomids sub-fossiles, du Québec à l'île d'Ellesmere, INRS rapport de Recherche R-900, ISBN 2-89146-430-3.
Lotter, A.F., Birks, H.J.B., Hofmann, W., and Marchetto, A. 1998. Modern diatom, cladocera, chironomid, and chrysophyte cyst assemblages as quantitative indicators for the reconstruction of past environmental conditions in the Alps. II. Nutrients. Journal of Paleolimnology, 19:443-463.
Quinlan, R., Smol, J.P., and Hall, R.I. 1998. Quantitative inferences of past hypolimnetic anoxia in south-central Ontario lakes using fossil midges (Diptera: Chironomidae). Canadian Journal of Fisheries and Aquatic Sciences, 55:587-596.
Rieradevall, M. and Brooks, S.J. 2001. An identification guide to subfossil Tanypodinae larvae (Insecta: Diptera: Chironomidae) based on cephalic setation. Journal of Paleolimnology, 25:81-99.
Wiederholm T. 1983. Chironomidae of the Holarctic region. Keys and diagnoses. Part 1. Larvae. Entomol. Scand. Suppl. 19: 1-457.
