 An ameghinornithid-like bird (Aves, Cariamae, ?Ameghinornithidae) from the early Oligocene of Egypt
An ameghinornithid-like bird (Aves, Cariamae, ?Ameghinornithidae) from the early Oligocene of Egypt
Article number: 18.1.5A
https://doi.org/10.26879/470
Copyright Society for Vertebrate Paleontology, February 2015
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 17 March 2014. Acceptance: 5 January 2015
{flike id=1046}
ABSTRACT
The Ameghinornithidae are an enigmatic group of potentially flightless Paleogene birds currently known from three European species. Herein, we report a new fossil that represents the first record of an ameghinornithid-like bird from Africa. The new specimen was collected from the early Oligocene Jebel Qatrani Formation exposed in the Fayum Depression of northern Egypt, and may represent the youngest record of the clade. The new specimen exhibits many similarities with European ameghinornithids, including the absence of an ossified supratendinal bridge on the tibiotarsus, a circular lateral condyle of the tibiotarsus, and a large medial epicondyle. The Fayum specimen is intermediate in size compared to the European ameghinornithids and is characterized by the absence of a projection of the medial condyle cranial to the lateral condyle and the presence of paired ridges (and associated grooves) proximal to the lateral condyle. The occurrence of Ameghinornithidae in the early Oligocene of Africa would have implications regarding the timing and pattern of biogeographic interchanges across the Tethys seaway.
Thomas A. Stidham. Key Laboratory of Vertebrate Evolution and Human Origins, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing 100044, China. presbyornis@gmail.com
N. Adam Smith. The Field Museum of Natural History, 1400 South Lake Shore Drive, Chicago, Illinois 60605, USA
adam_smith@utexas.edu
and
North Carolina Museum of Natural Sciences, 11 West Jones Street, Raleigh, North Carolina 2760, U.S.A.
Keywords: Avian evolution; fossil bird; Jebel Qatrani Formation; Paleogene; Strigogyps
Final citation: Stidham, Thomas A. and Smith, N. Adam. 2015. An ameghinornithid-like bird (Aves, Cariamae, ?Ameghinornithidae) from the early Oligocene of Egypt. Palaeontologia Electronica 18.1.5A: 1-8. https://doi.org/10.26879/470
palaeo-electronica.org/content/2015/1046-egyptian-ameghinornithid
INTRODUCTION
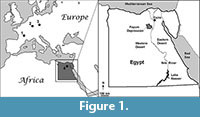 Ameghinornithidae is an enigmatic clade of potentially flightless, extinct birds thought to have had an herbivorous diet (Mayr and Richter, 2011). All previous records of the Ameghinornithidae are from the Middle Eocene of Germany (Messel and Geisel Valley) and late Eocene or early Oligocene of France (Quercy; Mayr, 2005, 2009; Figure 1). No records of the Ameghinornithidae currently are known from outside of Europe and only a single record of a potentially related taxon has been reported from Asia (i.e., Qianshanornithidae; see Mayr et al., 2013). We describe a new avian fossil that represents the first tentative African record of the Ameghinornithidae.
Ameghinornithidae is an enigmatic clade of potentially flightless, extinct birds thought to have had an herbivorous diet (Mayr and Richter, 2011). All previous records of the Ameghinornithidae are from the Middle Eocene of Germany (Messel and Geisel Valley) and late Eocene or early Oligocene of France (Quercy; Mayr, 2005, 2009; Figure 1). No records of the Ameghinornithidae currently are known from outside of Europe and only a single record of a potentially related taxon has been reported from Asia (i.e., Qianshanornithidae; see Mayr et al., 2013). We describe a new avian fossil that represents the first tentative African record of the Ameghinornithidae.
Systematically and taxonomically, the Ameghinornithidae have a convoluted history with respect to the placement of the clade and the taxonomy of referred species (see Mayr, 2005, 2009; Peters, 2007). Ameghinornithidae were once allied to phorusrhacoids, but are now considered to be a clade of uncertain phylogenetic affinities within Cariamae along with Idiornithidae, Bathornithidae, and Phorusrhacidae (i.e., outside of the core Gruiformes sensu Mayr, 2009). No species of Ameghinornithidae has been included in a phylogenetic analysis. The retention of multiple genera in Ameghinornithidae (i.e., Aenigmavis, Ameghinornis, and Strigogyps) was suggested by Peters (2007); however, other authors have placed all species into a single genus-level taxon, Strigogyps (Mayr, 2005, 2009; Mourer-Chauviré, 2006). Three previously recognized ameghinornithid species are known from Paleogene deposits in Europe (Table 1). To this list, we add a new record that may represent the first ameghinornithid from northern Africa. Referral of the Egyptian specimen to the Ameghinornithidae would expand the known geographic range of the group and may also represent the youngest record of the clade.
Geologic Context
The new ameghinornithid-like specimen was collected from the early Oligocene section of the Jebel Qatrani Formation of Egypt, which has an estimated age of~33.0 Ma (i.e., Rupelian; Seiffert, 2006). An Eocene avifauna from the lower section of the Jebel Qatrani Formation awaits description (Smith, personal observation). The fossil fauna and flora from this horizon are consistent with the interpretations of a relatively warm and wet climate (Bown and Kraus, 1988). The Jebel Qatrani Formation is composed of a mixed sequence of lithified siliciclastic and carbonaceous sediments, and has been interpreted as the result of primarily fluvial deposition (see Said, 1990 for further geologic and paleoenvironmental details). Vertebrate remains from this deposit include those of fish, turtles, a diverse mammalian assemblage, and birds (Bown et al., 1982). Previously described avian diversity from the Jebel Qatrani Formation includes many birds that are common in fluvially-dominated terrains today. These birds include cuckoos (Cuculiformes), falcons (Falconiformes), cranes and relatives (Gruiformes), shorebirds (Charadriiformes), flamingos (Phoenicopteriformes), cormorants (Pelecaniformes), herons, and storks (Ciconiiformes; Olson and Rasmussen 1986). Along with systematically enigmatic taxa such as Eremopezus eocaenus, the potential addition of Ameghinornithidae to the avifauna of the Jebel Qatrani Formation complements our knowledge of how the Oligocene aviafauna of northern Africa differs from that of similarly aged and better known avifaunas from Europe.
The sediments of the Fayum Depression in Egypt and the fissure fill deposits in the Quercy region of France hold a particular significance in the fossil history of birds as the major deposits documenting significant avian diversity through the Eocene—Oligocene climatic transition (EOCT; ~33—34 Ma; Rasmussen et al., 1987; Mourer-Chauviré, 2006; Mayr, 2009). That key temporal interval has been correlated with significant turnover in the mammalian fauna of Europe (i.e., the Grande Coupure), global climate change (cooling), and significant biogeographic changes (Holroyd and Maas, 1994; Liu et al., 2009; Mayr, 2011). Furthermore, molecular divergence estimates and the fossil record congruently suggest that the EOCT was a time of global avian radiation and avifaunal turnover (van Tuinen et al., 2006; Brown et al., 2007). From both of these Eocene—Oligocene fossiliferous deposits come some of the oldest records of avian groups (e.g., jacanas and cormorants), as well as fossils of enigmatic extinct groups of uncertain phylogenetic affinities (e.g., ameghinornithids and Eremopezus ; Rasmussen et al., 1987; Mayr, 2009). Paralleling the known mammalian fossil record, the late Eocene and early Oligocene appears to be a transitional period between extinct avian groups restricted to the early Paleogene and the appearance of many modern/extant clades within the later Oligocene and Neogene (Mayr, 2011). Despite the importance of this temporal interval for understanding the origin of the modern avifauna and the distribution of birds in the past, much of the avian record from the Fayum (i.e., Late Eocene and early Oligocene Jebel Qatrani Formation) remains unpublished.
MATERIALS AND METHODS
The taxonomy of Ameghinornithidae follows that suggested by Mayr (2005, 2009; i.e., all species are placed in Strigogyps), and the descriptive terminology utilizes the English equivalents of the Latin osteological nomenclature summarized by Baumel and Witmer (1993). Osteological measurements that follow those of von den Driesch (1976) were taken using digital calipers and were rounded to the nearest 0.1 mm. Ages of geologic time intervals are based on the International Geologic Time Scale (Gradstein et al., 2012).
Institutional Abbreviations
DPC, Duke Lemur Center, Division of Fossil Primates, Duke University, Durham, North Carolina, USA; GMH, Geiseltalmuseum Halle, Halle, Germany; BSP, Bayerische Staatssammlung für Paläontologie und Historische Geologie, München, Germany (Muséum de Munich sensu Gaillard, 1908); PQ, material curated in the collection of the Musée Guimet d'Histoire Naturelle de Lyon, France; SMF, Forschungsinstitut Senckenberg, Frankfurt am Main, Germany.
SYSTEMATIC PALAEONTOLOGY
AVES Linnaeus, 1758
CARIAMAE Fürbringer, 1888
cf. AMEGHINORNITHIDAE Mourer-Chauviré, 1981
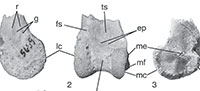
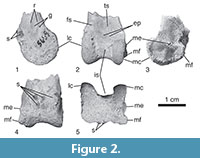 Material. The specimen (DPC 5659; Figure 2, Table 2) is the distal end of a right tibiotarsus collected by T.M. Bown in 1985 during a joint expedition of Duke University and the Egyptian Geological Museum.
Material. The specimen (DPC 5659; Figure 2, Table 2) is the distal end of a right tibiotarsus collected by T.M. Bown in 1985 during a joint expedition of Duke University and the Egyptian Geological Museum.
Locality and horizon. DPC 5659 was collected from Quarry E (sensu Bown and Kraus, 1988; Rasmussen et al., 1987) of the Jebel Qatrani Formation in the Fayum Depression of northern Egypt (Figure 1). Based on data summarized by Seiffert (2006), the estimated age of the Quarry E fauna is ~33.0 Ma (i.e., Rupelian, early Oligocene).
Taxonomic referral. The overall shape, fine morphological details and size of DPC 5659 agree with the species placed in Strigogyps (sensu Mayr, 2007, 2009). The five following apomorphies of the distal tibiotarsus of Ameghinornithidae were proposed by Peters (2007) and are shared by DPC 5659: (1) the absence of an ossified supratendinal bridge; (2) circular (i.e., not ovular) outline of lateral condyle; (3) strongly projected medial epicondyle; (4) proximal extension of the lateral condyle relative to the medial condyle; and (5) asymmetrical intercondylar sulcus. This presence of this unique suite of apomorphies in DPC 5659 supports its tentative referral to Ameghinornithidae (Strigogyps).
An ossified supratendinal bridge is plesiomorphic for crown group birds (i.e., Neornithes), but that structure is absent in the Fayum specimen (Figure 2.2). In addition to ratite paleognaths, several groups of neoavian birds have independently and secondarily lost the ossified supratendinal bridge of the tibiotarsus in adult individuals, and thus that character alone cannot be used to refer an individual fossil to a clade. Birds that lack an ossified supratendinal bridge include owls, hoatzin, some parrots, hornbills, oilbird, and some extinct Paleogene taxa (including Ameghinornithidae; see Mayr and Clarke, 2003; Mayr et al., 2013). DPC 5659 differs from all known owls in the absence of a deeply concave distal margin of the tibiotarsus where the intercondylar sulcus extends fully around the distal end, forming a deep notch or groove. The size of this new taxon is substantially larger than that of most of the other known birds that lack an ossified supratendinal bridge (e.g., oilbird and parrots). Other details of the conformation of the condyles and overall morphology are inconsistent with identification to the other clades that also lack an ossified supratendinal bridge.
Differential diagnosis. DPC 5659 differs from known species of Ameghinornithidae in the following characters: presence of a pair of associated grooves and ridges along the distal shaft of the tibiotarsus proximal to the lateral condyle (Figure 2.1); medial condyle not extending cranial to the lateral condyle; and intermediate in size between the known distal tibiotarsi of the species of Strigogyps (Table 2). See below for more detailed interspecific comparisons.
Specimen description. The shaft of the specimen is broken just proximal to the condyles. The caudal surface of DPC 5659 is partially covered in hardened sediment, and the caudal surfaces of the condyles and intercondylar sulcus are weathered and expose spongy-textured trabecular bone (Figure 2.4-5). The fragmentary and weathered preservation of DPC 5659 is consistent with other avian fossils from the Jebel Qatrani Formation and is not an indication of ontogenetic stage. There is no evidence of an ossified supratendinal bridge even though the astragalus and calcaneum are clearly fused to the tibia (i.e., an adult individual). Overall, the tibiotarsus is medially expanded at its distal end. The medial surface of the shaft proximal to the medial epicondyle is flat to slightly convex (Figure 2.3).
On the cranial surface, the extensor sulcus terminates in a mediolaterally wide and relatively deeply excavated pit (not to be confused with the fossa extensoria sensu Mayr and Rubilar-Rogers, 2010). A secondary, relatively smaller and shallower pit lies just distal to the pit (described above) that denotes the distal extent of the extensor sulcus. The distal edge of the secondary pit forms the proximal margin of the intercondylar sulcus. Putatively, these pits would have provided points of attachment (e.g., for extensor tendons or articular ligaments); however, the exact origin of the secondary pit is uncertain.
In lateral view (Figure 2.1), two grooves (i.e., linear fossae) are positioned at the junction of the lateral condyle and the shaft (i.e., largely proximal to the lateral epicondylar depression). The grooves are paired with two ridges that form the proximal margins of the grooves. The caudal groove is wider than the cranially positioned groove. Similar grooves have not been reported in other ameghinornithid species and the anatomical origin (i.e., muscular or tendinal insertions/passages) of these structures is uncertain.
The lateral condyle is circular in outline, and lacks a notch at the junction with the shaft (Figure 2.1). A lateral epicondyle is not present and the lateral condylar surface is only slightly concave. However, the lateral margin of the condyle is weathered and thus, the lateral surface of the condyle may once have been more concave. In cranial view, the lateral condyle extends more proximally than the medial condyle (i.e., approximately level with the distal extent of the extensor sulcus). The proximal end of the lateral condyle is more pointed than that of the more rounded medial condyle. The cranially facing edges of the condyles within the intercondylar sulcus are damaged. The intercondylar sulcus is a wide somewhat asymmetrical “U-shaped" incisure in distal view, but the damage to the condyles obscures its exact shape (Figure 2.5).
The medial condyle is distinctly semicircular in outline. The distal rim of the medial condyle is slightly damaged. In medial view, the proximal end of the medial condyle has a slight notch at the junction with the shaft (Figure 2.3). The medial epicondyle and the medial edge of the tibial cartilage articulation are projected far enough medially that they are visible in cranial view. The tibial cartilage articulation extends medial to the epicondyle. The medial epicondyle is positioned caudal to the cranial face of the shaft, and the area cranial to the epicondyle is concave.
The tibial cartilage articulation is partially covered by hardened sediment, but is laterally displaced relative to the distal end. The lateral edge of the tibial cartilage articulation is damaged. The distal margin of the medial edge of the tibial cartilage articulation flares medially (much like in diving ducks, e.g., Aythya). There is no evidence for a ridge within the broadly concave articulation surface. In caudal view, the tibial cartilage articulation is concave and appears symmetric (Figure 2.4).
Taxonomic comparisons. The morphology of DPC 5659 differs in many minor details from that of species of Strigogyps. The paired grooves on the lateral shaft of the tibiotarsus proximal to the condyle of DPC 5659 have not been reported previously in the Ameghinornithidae and could be an autapomorphy of the species represented by the Fayum specimen. However, DPC 5659 is not directly comparable with Strigogyps sp. of Mayr (2005) because that specimen comprises wing elements (e.g., humerus, ulna) and does not preserve the tibiotarsus. The same area of the distal shaft where the associated grooves and ridges are located appears damaged in the type specimen of Strigogyps (Aenigmavis) sapea, and this morphology is not present in Strigogyps dubius (Mayr, 2005; Peters, 2007, figure 2). The distal tibiotarsi of Strigogyps robustus are poorly preserved and do not allow the recognition of fine morphological details (Mayr, 2007). However, the dimensions of DPC 5659 are significantly smaller than those of the tibiotarsus of S. robustus (Table 2). While the distal tibiotarsus of this group is fairly well known, the amount of interspecific morphological variability within Ameghinornithidae is still poorly constrained.
The absence of a medial condyle that projects cranially (anteriorly) beyond the lateral condyle in DPC 5659 may suggest that it is not (very) closely related to species of Strigogyps (i.e., Ameghinornithidae sensu stricto). That putative ameghinornithid trait is only clearly present in S. dubius (Peters, 2007), and the relative projection of the condyles may be variable within the clade. Compared with S. dubius, the flattened area proximal to the lateral condyle on the cranial surface is not as distinct in the new Fayum specimen. The extensor sulcus is less deeply excavated in DPC 5659 than in S. dubius and S. sapea. In distal view, the intercondylar sulcus is more symmetrical than that of S. dubius (Figure 2.5), but given that the lateral surface of the medial condyle is somewhat damaged in DPC 5659, the exact shape of the intercondylar sulcus cannot be determined. The medial epicondyle is more caudally positioned in DPC 5659 than the condition in S. dubius (as illustrated in Peters, 2007, figure 3). The lateral condylar rim is worn in DPC 5659, and it cannot be discerned if it was inflated as those of S. sapea and S. dubius (sensu Peters, 2007). However, because the preserved rim in DPC 5659 is flush with the adjacent condylar surface, it probably extended further medially before it was damaged and would likely have looked similar to the inflated state attributed to other species of Strigogyps. The size of DPC 5659 is intermediate between that of S. dubius and S. sapea (Table 2), indicating that DPC 5659 is consistent in body size with the known ameghinornithids. The Fayum specimen lacks the deep medial epicondylar pit present in S. sapea (Figure 2; also see Mayr, 2005, figure 5).
DPC 5659 exhibits similarities with Qianshanornis rapax (Qianshanornithidae; Mayr et al., 2013), a taxon from the Paleocene of China that may have affinities with Ameghinornithidae. The systematic position of Qianshanornithidae in Aves has not been established unambiguously. The Fayum specimen and Q. rapax are both characterized by the presence of a flattened spot or embossment proximal to the lateral condyle. However, the near equal proximal extension of the condyles (i.e., the medial and lateral condyles extend nearly equidistant proximally) in Q. rapax differs from DPC 5659 and the species of Strigogyps. The condition in which the lateral condyle extends proximal to the level of the medial condyle is plesiomorphic for Cariamae and much of traditional Gruiformes. Additionally, the cranial projection of the lateral condyle in comparison to the medial condyle is greater in Q. rapax and DPC 5659 than that of S. dubius (Mayr et al., 2013; Figure 2).
DISCUSSION
The occurrence of an ameghinornithid-like bird in the early Oligocene of Africa (~33.0 Ma) may represent a geographic range extension for ameghinornithids (or their close relatives), and depending on the absolute age of the Quercy fossil material attributed to S. dubius, this Egyptian fossil also may represent a temporal extension for the clade as well (i.e., the youngest record of the group). The bulk of research on dispersal to and from Africa in the Paleogene has focused on mammals rather than birds, and the presence of an ameghinornithid-like bird in Africa provides new biogeographic data regarding distribution of birds along the Tethys Sea. Despite the much richer record of mammals from the Eocene and Oligocene of Asia, Europe, and Africa, there is no consistent biogeographic pattern clearly indicated as to dispersal among those continents (Holroyd and Maas, 1994; Grohé et al., 2012). However, the presence of related avian and mammalian taxa on both sides of the Tethys Sea (i.e., in Europe and North Africa), in addition to the occurrence of a potential ameghinornithid relative in the Paleocene of China (i.e., Q. rapax), indicates that dispersal did occur, even though the timing and directionality of those events cannot be constrained at present (Holroyd and Maas, 1994). At first glance, the older records of ameghinornithids in Europe would seem to suggest dispersal to Africa from Europe, but the paucity of early Paleogene terrestrial bird fossils in Africa could obscure the actual temporal sequence and dispersal pattern. Furthermore, the presence of a potential ameghinornithid relative in the Paleocene of China (Mayr et al., 2013) could be indicative of an Asian center of origin for ameghinornithid-like birds (i.e., Qianshanornithidae and Ameghinornithidae) with later dispersal to other parts of the Old World.
Just as the previous systematic grouping of Ameghinornithidae and the flightless Phorusrhacidae stirred up a great deal of biogeographic discussion regarding potential dispersal routes and modes between Europe and South America in the Paleogene (reviewed by Mayr, 2005; Mourer-Chauviré et al., 2011), the presence of an ameghinornithid-like bird in northern Africa raises questions about avian dispersal across or around the Tethys Sea during or prior to the early Oligocene. If the interpretation of ameghinornithids as flightless is correct, then their presence in Africa would seem unexpected. In contrast, the increased geographic distribution may actually indicate that ameghinornithids (and their close relatives) had at least limited flight capability (allowing for their distribution on both sides of the Tethys Sea). Mayr (2007) suggested that they might have had limited flight capability based on the no longer discernible impressions of primary feathers in Strigogyps robustus. Clearly more complete specimens are needed to adequately address these locomotive and biogeographic questions.
ACKNOWLEDGMENTS
We thank G. Gunnell and C. Riddle for access to the Duke University Lemur Center Fossil Primate Center collections and for assistance during TAS’s visit. We thank N. Zelenkov for general discussion on Paleogene avian taxonomy, P. Holroyd for information on Fayum mammalian biogeography, G. Mayr, C. Mourer-Chauviré, and anonymous reviewers for comments on an earlier draft of the manuscript. TAS is supported by the Chinese Academy of Science Professorship for Senior International Scientists (Grant No. 2011Y2ZA01), and NAS is supported by a National Evolutionary Synthesis Center Postdoctoral Fellowship (NESCent; NSF EF-0905606). This is Duke Lemur Center paper #1278.
REFERENCES
Baumel, J.J. and Witmer, L. 1993. Osteologia. p. 45-132. In Baumel, J.J., King, A.S., Breazile, J.E., Evans, H.E., and Vanden Berge J.C. (eds.), Handbook of avian anatomy: Nomina Anatomica Avium (Second Edition). Publications of the Nuttall Ornithological Club 23, Cambridge, MA, USA.
Bown, T.M., Kraus, M.J., Wing, S.L., Fleagle, J.G., Tiffney, B.H., Simons, E.L., and Vondra, C.F. 1982. The Fayum primate forest revisited. Journal of Human Evolution, 11:603-632.
Bown, T.M. and Kraus, M.J. 1988. Geology and paleoenvironment of the Oligocene Jebel Qatrani Formation and adjacent rocks, Fayum Depression, Egypt. United States Geological Survey Professional Paper, 1452:1-60.
Brown, J.W., Payn, R.B., and Mindell, D.P. 2007. Nuclear DNA does not reconcile ‘rocks’ and ‘clocks’ in Neoaves: a comment on Ericson et al . Biology Letters, 3:257-259.
Fürbringer, M. 1888. Untersuchungen zur Morphologie und Systematic der Vögel. T.J. Van Holkema, Amsterdam.
Gaillard, C. 1908. Les oiseaux des phosphorites du Quercy. Annales de l'Université de Lyon (Nouvelle Série), 23:1-178.
Gradstein, F.M., Ogg, J.G., Schmitz, M.D., and Ogg, G. M. 2012. The Geologic Time Scale (Volume 2). Elsevier, Boston.
Grohé, C., Morlo, M., Chaimanee, Y., Blondel, C., Coster, P., Valentin, X., Salem, M., Bilal, A.A., Jaeger, J.-J., and Brunet, M. 2012. New Apterodontinae (Hyaenodontida) from the Eocene locality of Dur At-Talah (Libya): Systematic, Paleoecological, and Phylogenetical Implications. Plos One, 7:e49054:1-19.
Holroyd, P.A. and Maas, M.C. 1994. Paleogeography, paleobiogeography, and anthropoid origins, p. 297-334. In Fleagle, J.G. and Kay, R.F. (eds.), Anthropoid Origins. Plenum Press, New York.
Lambrecht, K. 1935. Drei neue Vogelformen aus dem Lutétian des Geiseltales. Nova Acta Leopoldina (Neue Folge), 3:361-367.
Linnaeus, C. von. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima, reformata. Holmiae, Laurentii Salvii. [1-4].
Liu, Z., Pagani, M., Zinniker, D., DeConto, R., Huber, M., Brinkhuis, H., Shah, S.H., Leckie, R.M., and Pearson, A. 2009. Global cooling during the Eocene—Oligocene climate transition. Science, 323:1187-1190.
Mayr, G. 2005. ‘‘Old World phorusrhacids’’ (Aves, Phorusrhacidae); a new look at Strigogyps (‘‘Aenigmavis’’) sapea (Peters 1987). PaleoBios, 25:11-16.
Mayr, G. 2007. Synonymy and actual affinities of the putative Middle Eocene “new world vulture” Eocathartes Lambrecht, 1935 and “hornbill” Geiseloceros Lambrecht, 1935 (Aves, Ameghinornithidae). Paläontologische Zeitschrift, 81:457-462.
Mayr, G. 2009. Paleogene Fossil Birds. Springer-Verlag, Berlin.
Mayr, G. 2011. Two-phase extinction of “Southern Hemispheric” birds in the Cenozoic of Europe and the origin of the Neotropic avifauna. Palaeobiology and Palaeoenvironment, 91:325-333.
Mayr, G. and Clarke, J.A. 2003. The deep divergences of neornithine birds: a phylogenetic analysis of morphological characters. Cladistics, 19:527-553.
Mayr, G. and Richter, G. 2011. Exceptionally preserved plant parenchyma in the digestive tract indicates a herbivorous diet in the Middle Eocene bird Strigogyps sapea (Ameghinornithidae). Palaontologische Zeitschrift, 85:303-307.
Mayr, G. and Rubilar-Rogers, D. 2010. Osteology of a new giant bony-toothed bird from the Miocene of Chile, with a revision of the taxonomy of Neogene Pelagornithidae. Journal of Vertebrate Paleontology, 30:1313-1330.
Mayr, G., Yang, J., de Bast, E., Li, C., and Smith, T. 2013. A Strigogyps -like bird from the Middle Paleocene of China with an unusual grasping foot. Journal of Vertebrate Paleontology, 33:895-901.
Mourer-Chauviré, C. 1981. Premiére indication de la presence de Phorusrhacidés, Famile d'oiseaux géants d'Amérique du Sud, dans le Tertiaire Européén: Ameghinornis nov. gen. (Aves, Ralliformes) des Phosphorites du Quercy, France. Géobios, 14:637-647.
Mourer-Chauviré, C. 2006. The avifauna of the Eocene and Oligocene phosphorites du Quercy (France): an updated list. Strata (Ser. 1), 13:135-149.
Mourer-Chauviré, C., Tabuce, R., Mahboubi, M., Adaci, M., and Bensalah, M. 2011. A Phororhacoid bird from the Eocene of Africa. Naturwissenschaften, 98:815-823.
Olson, S.L. and Rasmussen, D.T. 1986. Paleoenvironment of the earliest hominoids: new evidence from the Oligocene avifauna of Egypt. Science, 233:1202-1204.
Peters, D.S. 1987. Ein ‘‘Phorusrhacide’’ aus dem Mittel-Eozän von Messel (Aves: Gruiformes: Cariamae). Documents des Laboratoires de Géologie de Lyon, 99:71-87.
Peters, D.S. 2007. The fossil family Ameghinornithidae (Mourer-Chauviré 1981): a short synopsis. Journal of Ornithology, 148:25-28.
Rasmussen, D.T., Olson, S.L., and Simons, E.L. 1987. Fossil birds from the Oligocene Jebel Qatrani Formation, Fayum Province, Egypt. Smithsonian Contributions to Paleobiology, 62:1-20.
Said, R. 1990. Cenozoic, p. 451-486. In Said, R. (ed.), The Geology of Egypt. A.A. Balkema, Brookfield.
Seiffert, E.R. 2006. Revised age estimates for the later Paleogene mammal faunas of Egypt and Oman. Proceedings of the National Academy of Sciences, 103:5000-5005.
van Tuinen, M., Stidham, T.A., and Hadly, L. 2006. Tempo and mode of modern bird evolution observed with large-scale taxonomic sampling. Historical Biology, 18:205-221.
von den Driesch, A. 1976. A guide to the measurement of animal bones from archaeological sites. Peabody Museum Bulletin, 1:103-129.
