

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
Joseph P. Botting State Key Laboratory of Palaeobiology and Stratigraphy
State Key Laboratory of Palaeobiology and Stratigraphy
Nanjing Institute of Geology and Palaeontology
Chinese Academy of Science
No. 39 East Beijing Road
Nanjing 210008
China
Joseph Botting is a Young International Scientist at the Nanjing Institute of Geology and Palaeontology. He started working in the Builth Inlier as an undergraduate at Cambridge, and completed his PhD, which was partly on faunas from the area, at the University of Birmingham in 2000. Most of his publications have been on sponges, but he has also worked on echinoderms, worms, trilobites, palaeoecology and several Ordovician Lagerstätten. His research is currently focused on the early evolution of sponges, with forays into exceptionally preserved biotas in China, Morocco, the UK and North America.
Lucy A. Muir. State Key Laboratory of Palaeobiology and Stratigraphy
State Key Laboratory of Palaeobiology and Stratigraphy
Nanjing Institute of Geology and Palaeontology
Chinese Academy of Science
No. 39 East Beijing Road
Nanjing 210008
China
Lucy Muir is primarily a graptolite worker and palaeoecologist, but has published on a variety of other things, including museum collections and modern ecology. She received her undergraduate degree from the University of Cambridge (1997) and then undertook an MSc in Palaeobiology at the University of Bristol (1999). Since gaining her PhD from the University of Edinburgh in 2004, she has worked as a museum curator and a freelance geological consultant. She is currently a postdoctoral researcher at the Nanjing Institute of Geology and Palaeontology, where she is working on a comparison of Ordovician Lagerstätten in Wales, China and Morocco.
TABLE 1. Complete list of the fauna known from the Holothurian Bed at the time of writing; a total of ten sponge species and two dendroids are included, although the exact numbers may increase on further study. Abundance notes are semi-quantitative: "rare": encountered on three or fewer occasions; "occasional": encountered on fewer than half of collecting trips; "frequent": encountered in low numbers on most collecting trips; "common": encountered on almost all collecting trips, often with several individuals; "superabundant": encountered on almost every slab.
|
Group |
Taxon |
Notes |
|
Trilobites |
Ogyginus corndensis |
Common, including as complete juveniles |
|
|
Ogygiacarella debuchii |
Moderately common |
|
|
Cnemidopyge parva |
Occasional complete moults |
|
|
Protolloydolithus reticulatus |
Occasional |
|
|
Platycalymene simulata |
Rare – one complete meraspis |
|
|
Rorringtonia kennedyi |
Occasional |
|
|
Geragnostus mccoyii |
Occasional |
|
|
odontopleurid indet. |
Rare – one rib fragment |
|
|
Microparia lusca |
Rare – few near-complete adults |
|
|
Other cyclopygids (indet.) |
Rare fragments |
|
|
Barrandia cordai |
Rare |
|
Brachiopods |
Apatobolus? micula |
Superabundant |
|
|
Palaeoglossa attentuata |
Common |
|
|
Monobolina ramsayi |
Occasional |
|
|
Opsiconidion nudum |
Occasional |
|
|
Schizocrania cf. salopiensis |
Occasional–only encrusting undetermined substrate |
|
|
Meristopacha granulata? |
Rare |
|
|
Dalmanellid indet. |
Rare |
|
Graptolites |
Cryptograptus tricornis |
Frequent |
|
|
Hustedograptus? teretiusculus |
Occasional |
|
|
Dendrograptus spp. |
Rare; at least two species |
|
Echinoderms |
Cystoid indet. |
Rare – one specimen and one possible |
|
|
Anatifopsis? sp. |
Common |
|
|
Galliaecystis? sp. |
Rare – fragments of four specimens |
|
|
Solute undet. |
Rare – one articulated specimen |
|
|
Carpoid indet. |
Rare – one specimen |
|
|
Oesolcucumaria eostre gen. et sp. nov. |
Occasional |
|
Cnidarians |
Sphenothallus sp. |
rare – few specimens |
|
|
Conulariid indet. |
Rare – one specimen |
|
Ostracodes |
Undetermined smooth form |
Common |
|
|
Bullaeferum sp. |
Occasional |
|
|
Nanopsis sp. |
Occasional |
|
Chordates |
Conodonts |
Occasional as isolated elements; rare as clusters (pellets?) |
|
Molluscs |
Bivalve indet |
Rare |
|
|
Gastropod indet. |
Rare – few specimens |
|
|
Nautiloids indet. |
Rare – few specimens |
|
Sponges |
Pirania? sp. |
Rare |
|
|
Wapkia-like form |
Occasional |
|
|
Xylochos? sp. |
Occasional |
|
|
Reticulosan sp. 1 |
Frequent |
|
|
Reticulosan sp. 2 |
Frequent |
|
|
Hexactinellid indet. |
Rare |
|
|
Sponges undet. (several spp.) |
Rare |
|
|
Eiffeliid indet. |
Isolated spicules only |
|
Worms |
Palaeoscolecid gen. et sp. nov. (Botting et al. in press) |
One confirmed specimen; many too poorly preserved to identify |
|
|
Palaeoscolecid undet. |
One specimen with different plating to Palaeoscolecid gen. et sp. nov. |
|
|
Scolecodonts |
One associated pair of elements |
|
Undet. |
Algal remains? |
Remains on which ostracodes are feeding |
TABLE 2. Approximate proportions of the fauna in terms of collected and life abundance, based on largely non-analytical collecting over 10 years; the figures are therefore not rigorous, but instead estimated values from our observations. The correction factor is based on the ratio of articulated to fragmented scleritomes of Anatifopsis?, representing the proportion of individuals affected by rapid burial, which was then applied to other multi-element scleritomes. Deviations from this factor were employed to allow for ecdysis and additional requirements of soft-tissue preservation; see text for more details.
|
|
complete (c) |
disarticulated (d) |
Correction |
Estimated relative |
|
Apatobolus? micula |
>100 |
Many |
|
>>1000 |
|
sponges |
50 |
Not visible |
20 |
1050 |
|
palaeoscolecids |
17 |
2 fragments |
~20 |
357 |
|
trilobites |
10 (non-moult) 50 (moults) |
? |
20 20/5=4 |
<210 <250 |
|
ostracodes |
30 |
Many |
20/4=5 |
180 |
|
holothurians |
8 |
Not visible |
20+ |
>168 |
|
mitrates |
7 |
~140 |
20 |
~147 |
|
other brachiopods |
? |
100 |
- |
100 |
|
graptolites |
80 |
n/a |
- |
80 |
|
solutes |
1 |
Not visible |
20 |
21 |
|
bivalves |
5 |
n/a |
- |
5 |
|
cornutes |
0 |
4 |
- |
4 |
|
nautiloids |
3 |
n/a |
- |
3 |
|
sphenothallids |
3 |
n/a |
- |
3 |
|
gastropods |
2 |
n/a |
- |
2 |
TABLE 3. Expected taxonomic composition of the preserved fauna under different degrees of preservation. "Poor preservation" implies an entirely disarticulated and washed assemblage of shelly fragments. "Moderate preservation" implies a fauna lacking abrupt burial and soft tissue, but without extended sea-floor exposure or winnowing. The observed fauna is extensive, but will inevitably be increased by further collecting, as taxa from the tail of the hollow curve are discovered. The total diversity of macrobiota in the local area at the time of deposition is impossible to ascertain, but it was likely to have been in excess of 100 species, by comparison with major Lagerstätten such as the Fezouata Biota (Van Roy et al., 2010) and the Burgess Shale (Conway Morris, 1986). In these deposits, many taxa are localised, and were found only after extensive collecting.
|
Complete biota |
Holothurian Bed |
Moderate preservation |
Poor preservation |
|
Trilobites (?11 spp) |
Trilobites (11 spp) |
Trilobites (11 spp) |
Trilobites (<11 spp, frag.) |
|
Brachiopods (?7 spp) |
Brachiopods (7 spp) |
Brachiopods (7 spp) |
Brachiopods (?7 spp) |
|
Other shelly taxa (?8 spp) |
Other shelly taxa (8 spp) |
Other shelly taxa (8 spp) |
Other shelly taxa (<8 spp) |
|
Graptolites (?5 spp) |
Graptolites (5 spp) |
Graptolites (5 spp) |
Graptolites (?5 spp) |
|
Sponges (? 10 spp) |
Sponges (10 spp) |
Sponges (spicules only) |
Sponges (spicules only) |
|
Conodonts (?1 sp.) |
Conodonts (1 sp.) |
Conodonts (1 sp.) |
Conodonts (1 sp.) |
|
Carpoids etc. (?3 spp) |
Carpoids ( 3 spp) |
Carpoids (2 spp) |
Anatifopsis? plates only |
|
Cystoids (?1 sp.) |
Cystoids (1 sp.) |
- |
- |
|
Algae (?1 sp.) |
Algae? (1 sp.) |
- |
- |
|
Holothurians (?1 sp.) |
Holothurians (1 sp.) |
- |
- |
|
Palaeoscolecidans (?2 spp) |
Palaeoscolecidans (2 spp) |
- |
- |
|
Unmineralised arthropods |
- |
- |
- |
|
Unmineralised worms |
- |
- |
- |
|
Other soft-bodied taxa |
- |
- |
- |
|
Estimated total: ? |
51 |
35 |
<30 |
FIGURE 1. Stratigraphic column and map showing the location of the Holothurian Bed locality near Llandrindod, Powys, Wales (inset map shows location within southern UK). Llanfawr Mudstones Lagerstätte refers to Botting et al. (2011), and the Llandegley Rocks Lagerstätte was described by Botting (2005). CMF: Camnant Mudstones Formation. The boundary between the Didymograptus murchisonii and Hustedograptus? teretiusculus biozones is not well constrained, except in the stream itself, where it is somewhat faulted (Sheldon 1987b).
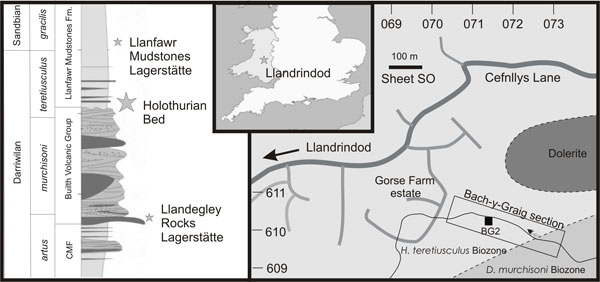
FIGURE 2. Outcrop of the Holothurian Bed, in the upper part of the section; the interval described here overlies a thick tuff bed visible in the upper half of the view. Beds underlying the tuff are blue-grey mudstones with similar fauna to the non-exceptional components found above it. The tuff bed is 27 cm thick.

FIGURE 3. Trilobites from the Holothurian Bed, Bach-y-Graig. 1, Ogygiocarella? sp., recently moulted, wrinkled specimen NMW2011.2G.7, scale bar equals 5 mm. 2,Ogyginus corndensis, articulated moult NMW2011.2G.6, scale bar equals 1 mm. 3, Ogyginus corndensis meraspis, NMW2011.2G.31, scale bar equals 1 mm; 4, Cnemidopyge parva NMW2011.2G.28, scale bar equals 5 mm; 5, Protolloydolithus reticulatus NMW2011.2G.4, scale bar 5mm; 6, Microparia sp. NMW2011.2G.20, scale bar equals 1 mm; 7, Protolloydolithus reticulatus showing replacement of skeleton by pyrite NMW2011.2G.30, scale bar 1 mm; 8, Ogyginus corndensis NMW2011.2G.15, scale bar equals 1 mm; 9, Cluster of trilobite fragments NMW2011.2G.27, scale bar equals 1 mm.

FIGURE 4. Representative fossils from the Holothurian Bed, Bach-y-Graig. 1, undescribed bivalve NMW2011.2G.11, scale bar equals 1 mm; 2, fragment of undescribed hexactinellid sponge, showing typical preservation due to weathered pyrite, NMW2011.2G.14, scale bar equals 1 mm; 3, undescribed palaeoscolecidan worm with dark carbonaceous and/or phosphatic preservation of plates NMW2011.2G.26, scale bar equals 1 mm.

FIGURE 5. Echinoderms from the Holothurian Bed, Bach-y-Graig. 1, articulated mitrate (Anatifopsis?) NMW2011.2G.12, scale bar equals 1 mm; 2, undetermined carpoid(?) echinoderm NMW2011.2G.25, scale bar equals 1 mm; 3, undetermined "cystoid" with arachnoid plates, NMW2011.2G.34, scale bar equals 1 mm. See also Figure 9 for holothurian specimens.

FIGURE 6. Graptolites and brachiopods from the Holothurian Bed, Bach-y-Graig. 1, Cryptograptus tricornis showing typical preservation for graptolites NMW2011.2G.2, scale bar 2 mm; 2, Hustedograptus? teretiusculus with combination of carbonaceous and probably aluminosilicate preservation NMW2011.2G.3, scale bar equals 2 mm; 3, Dendrograptus sp. NMW2011.2G.1, scale bar equals 5 mm; 4, Palaeoglossa attenuata NMW2011.2G.19, scale bar equals 2 mm; 5, Schizotreta sp. encrusting a weakly preserved substrate, NMW2011.2G.9, scale bar equals 5 mm; 6, Small cluster of Apatobolus? micula (see also Figure 8.5) NMW2011.2G.37, scale bar equals 2 mm.

FIGURE 7. Species-abundance curve obtained from in situ sampling exercise over two days, excluding Apatobolus? micula. Note that the curve appears to level out at approximately 48 specimens, but we are aware of a minimum diversity of 52 species at the site, and the true diversity will be substantially higher.
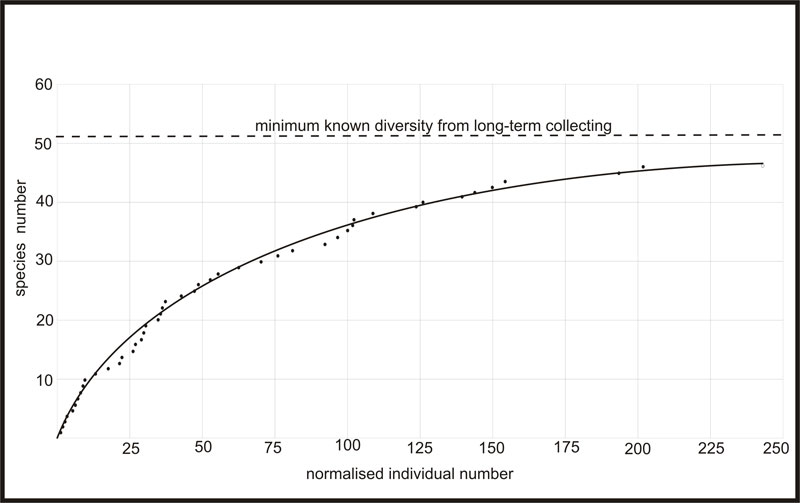
FIGURE 8. Specimens from the Holothurian Bed, Bach-y-Graig showing ecological relationships. 1, organic matter with feeding ostracodes (three examples arrowed) NMW2011.2G.8, scale bar equals 1 mm; 2, Cluster of dissociated Anatifopsis? plates and trilobite sclerites interpreted as possible cephalopod midden, NMW2011.2G.16, scale bar equals 5 mm; 3, pellet composed of fibrous carbonaceous material NMW2011.2G.23, scale bar equals 1 mm; 4, pellet composed of crushed valves of Apatobolus? micula, NMW2011.2G.24, scale bar equals 1 mm; 5, Apatobolus? micula showing repair from probable attempted predation damage, NMW2011.2G.37, scale bar equals 1 mm.

FIGURE 9. Holothurian Oesolcucumaria eostre gen. et sp. nov. 1-3, holotype NHM EE8734; 1, overall view; 2, detail of part of calcareous ring and short ambulacrum; 3; camera lucida drawing of area illustrated in 3; 4, paratype NHM EE8736, large weathered specimen, near-complete; 5, paratype NHM EE8737, anterior part of holothurian showing weakly pyritised oral ring and possibly also ambulacral structures; 6, paratype NMW2011.2G.21, complete weathered specimen with poorly preserved fine detail; 7, paratype NHM EE8739, isolated calcareous ring. Scale bar equals 1 mm.

Fauna and ecology of the Holothurian Bed, Llandrindod, Wales, UK (Darriwilian, Middle Ordovician), and the oldest articulated holothurian
Plain Language Abstract
Trilobites are among the most popular of fossil groups, and during the Ordovician Period, they often dominated the fossil faunas of mudstone beds deposited on the outer continental shelf. This apparent abundance may be misleading, however: because trilobites have a mineral (calcite) skeleton, they preserved easily, biasing the fossil record in their favour. It is much rarer to discover more delicate organisms in fine detail, although offshore muddy habitats today are often dominated by sponges, brittle-stars and sea-cucumbers instead of arthropods. This paper describes a new deposit from the Middle Ordovician (approximately 462 million years ago) of Wales that has yielded not only the trilobites, but a diverse community of organisms including sponges, palaeoscolecidan worms and the extinct carpoid echinoderms. It also preserves the oldest known complete sea-cucumbers, the bodies of which are so soft that they are very rarely fossilized even in sites of exceptional preservation. These small, globular fossils show primitive features of the group that help relate them to other types of echinoderm. Most importantly, the fauna of this site shows that fossil communities that appear to be typical of Ordovician muddy seas can in reality be ecologically very different, and an analysis of the available data suggests that trilobites were a relatively minor component of the fauna, after brachiopods, sponges and armoured worms.
Resumen en Español
Fauna y ecología de la Holothurian Bed, Llandrindod, Gales, Reino Unido (Darriwiliense, Ordovícico medio) y la holoturia articulada más antigua
Al contrario de lo que ocurre en el Cámbrico, los yacimientos con una preservación excepcional (Lagerstätten) del Ordovícico muestran diferencias significativas entre ellos, lo que sugiere una diversidad mucho mayor de las paleocomunidades durante este intervalo. No está claro, sin embargo, qué parte de la diversidad ecológica del Ordovício es debida a lo atípico de las facies o a que las comunidades estén representadas por la mayor parte de las biotas excepcionales conocidas. En este trabajo describimos un nuevo Lagerstätte, preservado gracias a una alta tasa de sedimentación y a una piritización temprana, del Ordovícico medio (Darriwiliense, Biozona de Hustedograptus? teretiusculus) de Gales. Un análisis preliminar de la ecología de la fauna, que incluye un nuevo método para estimar la abundancia en vida a partir de la abundancia preservada de los distintos grupos, ha revelado que, aunque la asociación fósil esté dominada por los trilobites, los organismos dominantes en la comunidad viva eran los braquiópodos, las esponjas de los órdenes Reticulosa y Protomaxonida y los paleoescolécidos, siendo los carpoideos y los holoturoideos tan abundantes como los trilobites. Bajo condiciones normales de preservación, esta comunidad aparecería como la típica fauna de mudstone del Ordovícico, suponiendo que se trate de una comunidad relativamente normal de ambiente marino abierto. La biota no se asemeja mucho a ninguna de las faunas excepcionales descritas previamente en el Ordovícico, lo que sugiere que hay una gran cantidad de diversidad oculta en las comunidades de esa edad. Entre los taxones excepcionales se encuentra el holoturoideo articulado más antiguo conocido, Oesolcucumaria eostre gen. et sp. nov., que es una pequeña forma globular con ambulacros parcialmente esqueletizados.
Palabras clave: Echinodermata; preservación excepcional; nuevo género; nueva especie; paleoecología; piritización
Traducción: Miguel Company
Résumé en Français
Une révision du trilobite dévonien malvinokaffrique dalmanitidé Dalmanitoides Delo, 1935, sur la base de nouvelles données provenant d’Argentine
A la lumière de nouvelles informations sur l’holotype et de matériel additionnel provenant de Dévonien inférieur de la région type d’Argentine, le trilobite dalmanitidé Dalmanitoides Delo, 1935, est re-diagnostiqué et considéré comme un dalmanitidé plutôt qu’un synphoriiné. Des comparaisons suggèrent que Gamonedaspis Braniša et Vaněk, 1973, est un synonyme de Dalmanitoides, de sorte que ce genre dévonien inférieur-moyen inclue au moins cinq espèces : quatre nommées formellement en Amérique du Sud D. drevermanni (Delo, 1935), D. boehmi (Knod, 1908), D. scutata (Braniša et Vaněk, 1973), et D. accola (Clarke, 1913), de même qu’une espèce d’Afrique du Sud traitée ici en nomenclature ouverte (D. sp. A). Bien que le nombre d’espèces de Dalmanitoides suggère que la diversification des dalmanitinés cosmopolites était déjà en place dans les bassins malvinokaffriques, une affinité forte avec le genre boréal et plus ancien Roncellia Lespérance et Bourque, 1971, suggérerait qu’une migration depuis le domaine paléobiogéographique d’Amérique de l’Est pendant le Dévonien inférieur serait à l’origine du clade, ce qui est en accord avec les schémas paléobiogéographiques reconnus sur la base de la distribution des synphoriinés.
MOST CLES : trilobites dalmanitidés ; Dévonien ; Argentine ; malvinokaffrique ; Dalmanitoides; Gamonedaspis; Systématique; Paléobiogéographique
Translator: Olivier Maridet
Deutsche Zusammenfassung
Ökologie und Fauna der Holothurien-Schicht, Llandrindod, Wales, UK (Darriwillium, Mittleres Ordovizium) unddie älteste artikulierte Holothurie
Im Gegensatz zu kambrischen Lagerstätten weisen die exzellent erhaltenen ordovizischen Fundstätten eine taxonomisch deutlich reichhaltigere Fauna auf. Dies weist auf eine größere Diversität der damaligen Lebensgemeinschaft hin. Unklar istallerdings, inwieweit diese hohe ökologische Diversität im Ordovizium auf eine atypische Fazies zurückzuführen ist, bzw. in wieweit diese auf Lebensgemeinschaften basieren, die auf Ausnahme-Biota zurückzuführen sind.
Diese Arbeit beschreibt eine neue Lagerstätte aus dem mittleren Ordovizium von Wales (Darriwilium, Hustedograptus? teretiusculus Biozone), die durch hohe Sedimentationsraten und frühe Pyritisierung entstanden ist. Die vorläufigen Untersuchungen zeigen, dass die Fauna nicht, wie angenommen, von Trilobiten geprägt ist, sondern dass Brachipoden, reticulosane und protomonaxinoide Schwämme und Paläoscoleciden dominieren. Desweiteren treten Carpoiden und Holothurien fast ebenso reichhaltig wie Trilobiten auf. Diese Untersuchungen beinhalten auch eine neuartige Methodik, um die erhaltende Menge der verschiedenen Gruppen in eine geschätzte Menge im Lebendzustand zu übertragen. Unter gewöhnlichen Erhaltungsumständen würde sich diese Lebensgemeinschaft als typische ordovizische Tonstein-Fauna darstellen, was wiederum implizieren würde, dass eine relativ normale Vergesellschaftung des offenen Meeres repräsentiert ist. Die vorliegende Zusammensetzung der Fauna ähnelt den bisher beschriebenen ordovizischen Faunen der Ausnahmefundstätten wenig. Dies kann als Hinweis darauf angesehen werden, dass es in den Vergesellschaftungen dieses Zeitalters eine starke verdeckte Diversität gibt. Unter den besonderen Taxa ist die älteste bekannte artikulierte Holothurie, Oesolcucumaria eostre gen. et sp. nov., eine kleine globulare Form mit teilweise skeletisiertemAmbulakralsystem.
Keywords: Echinodermata; Ausnahmeerhaltung; neue Gattung; neue Art; Paläoökologie; Pyritisierung
Translators: Eva Gebauer and Anke Konietzka
Arabic

Translator: Ashraf M.T. Elewa
Polski Abstrakt
In progress
Translators: Dawid Mazurek and Robert Bronowicz
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024