

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
APPENDIX 1
List and description of 156 characters and states used in our phylogenetic analyses. Ordered characters are indicated with an asterisk (*).
(1) four incisors present (I5 lost)
(0) I5 similar in morphology to I1-4
(1) I5 pointed and strongly canine-like
4. *Degree of development of lingual shelf on P3
(1) small/weakly developed shelf
(2) well-developed lingual shelf
(3) well-developed lingual shelf that extends to the buccal side of P3,
(0) P3 major cusp laterally compressed
(1) P3 major cusp laterally enlarged but not conical
(2) P3 major cusp large and conical, but P3 is narrower than M1
(3) P3 Major cusp large and conical, and P3 is wider than M1,
(0)stylar crest present on StD connects to metastylar tip
(1) StD is a conical cusp not connected to the stylar crest running to the metastylar tip
(2) StD is a conical cusp and no stylar crest present,
(1) small anterior cingulum lingual to anterior tip of tooth, no connection to talon
(2) anterior cingulum enlarged and connects to talon as small shelf,
(1) small anterior cingulum lingual to anterior tip of tooth
(2) small anterior cingulum connected to protocone shelf by a small shelf
(3) large anterior cingulum expanded further up to the protocone,
9. *Direction of preparacrista of M1
(0) preparacrista anterobuccally orientated to connect to anteriorly positioned StB.
(1) preparacrista buccally orientated (perpendicular to tooth row) then posterobuccally orientated
(2) preparacrista posterobuccally orientated to connect to posteriorly located StB
(3) preparacrista posterobuccally orientated to connect to StB and then reconnects with postparacrista posteriorly,
11. *Morphology of the centrocrista on M1 and M2
(0) postparacrista and premetacrista contact each other, forming a complete centrocrista that connects the paracone and metacone (more lingually).
(1) postparacrista and premetacrista contact each other, forming a complete centrocrista that connects the paracone and metacone (more buccally)
(2) postparacrista does not connect to the premetacrista and ends at the base of StB, premetacrista ends at base of StD ,
12. *Morphology of centrocrista on M3
(0) postparacrista and premetacrista contact each other, forming a complete centrocrista that connects the paracone and metacone (more lingually).
(1) postparacrista and premetacrista contact each other, forming a complete centrocrista that connects the paracone and metacone (more buccally)
(2) postparacrista does not connect to the premetacrista and ends at the base of StB, premetacrista ends at base of StD ,
(1) i3 posterior cusp present but small
(2) I3 posterior cusp present and large,
(0) p3 longer or equal in length to p2
15. Presence of preentocristid
16. Direction of preentocristid of m1-3
17. Cusp within the hypoflexid region, between the talonid and trigonid on the buccal side
(0) no median buccal cusp present
(1) median buccal cusp present ,
18. Distinction between lower molar crowns and roots
(1) crown and root indistinct (other than for limit of enamel)
19. Hypoconulid posterior to entoconid
(0) hypoconulid positioned posterobuccal to the entoconid
(1) hypoconulid positioned almost directly posterior to the entoconid ,
(0) entoconid and hypoconid large, talonid relatively wide, crests clearly identifiable.
(1) entoconid and hypoconid smaller than in state 0, talonid smaller
(2) further reduction of entoconid and hypoconid, talonid very small, crest poorly defined,
21. Snout length and premaxilla size
(0) premaxilla taller than it is long; maxilla-nasal contact longer than premaxilla-nasal contact (usually correlated with a relatively short snout)
(1) premaxilla longer than it is tall; premaxilla-nasal contact longer than maxilla-nasal contact (usually correlated with a relatively elongate snout) ,
(1) slender nasals, not markedly expanded posteriorly ,
23. *Position of nasal-frontal suture/maximum posterior extension of nasals
(0) posterior to the anterior rim of the orbit when viewed laterally
(1) nasals terminate just anterior to the orbit when viewed laterally (associated with a wide maxilla-frontal suture)
(0) nasals terminate well anterior to the anterior margin of the orbit,
(0) long infraorbital canal (longer than half the molar row).
(1) short infraorbital canal (shorter or equal to half the molar row).,
(0) jugal invades the maxilla and extends onto the facial region of the skull (not bifid)
(1) maxilla invades the zygomatic arch so that the jugal has two thin wings around the posterior maxillary flange (bifid),
(1) partially developed lacrimal crest
(2) fully developed lacrimal crest,
(1) weak or partial fossa development
(2) very deep antorbital fossa,
(0) orbitosphenoid identifiable in lateral view as a large ossification
(1) orbitosphenoid identifiable in lateral view as a small ossification
(2) orbitosphenoid is very small or absent and not obvious in lateral view,
(0) alisphenoid-parietal contact
(1) squamosal-frontal contact ,
30. *Sphenorbital fissure and foramen rotundum
(0) sphenorbital fissure slightly larger than foramen rotundum. Both canals are tube-like in shape, especially the foramen rotundum
(1) sphenorbital fissure is enlarged and more widely open, and there is a reduction in the length of the tube leading to the foramen rotundum
(2) sphenorbital fissure is further enlarged and open, and there is no tube leading to the foramen rotundum, which instead appears,
31. Presence of Accessory fenestrae
32. Presence of dividing septa in maxillopalatine fenestrae
(0) septum present in maxillopalatine fenestrae
(1) septum absent in maxillopalatine fenestrae,
(0) foramen bound by squamosal or squamosal and alisphenoid
(1) foramen bounded medially by the petrosal,
34. Morphology of the primary foramen ovale
(0) primary foramen ovale is between alisphenoid and petrosal
(1) primary foramen ovale is entirely within the alisphenoid,
35. *Morphology of the secondary foramen ovale
(0) secondary foramen ovale absent
(1) secondary foramen ovale defined by a complete strut or bridge formed by the alisphenoid, but the primary foramen ovale is still visible
(2) secondary foramen ovale present, and extensive ossification of the alisphenoid means that the primary foramen ovale is no longer visible in intact skulls,
36. *Morphology of the ectotympanic
37. *Degree of inflation of the alisphenoid tympanic process
(0) alisphenoid tympanic process small, lateral and medial walls open.
(1) alisphenoid tympanic process largely open but medial wall enclosed greater than in state 0, and hypotympanic sinus is also enlarged relative to state 0
(2) alisphenoid tympanic process and hypotympanic sinus further enlarged.relative to state 1; hypotympanic sinus walled posteriorly by the alisphenoid
(3) alisphenoid tympanic process and hypotympanic sinus greatly hypertrophied,
38. Alisphenoid tympanic process shape
(1) ventrally rounded, anterior boundary rounded does not extend as far as the transverse foramen or foramen ovale
(2) ventrally rounded, anterior boundary pointed and extends as far as the transverse foramen
(3) ventrally angular, anterior boundary pointed and terminates anterior to the transverse foramen,
39. *Morphology of the rostral tympanic process of the petrosal
(0)rostral tympanic process of the petrosal absent or very small
(1) rostral tympanic process of the petrosal forms a distinct projecting process that partially walls the posteromedial margin of the hypotympanic sinus .
(2) rostral tympanic process of the petrosal further enlarged, forming elongate crestlike process that extends the length of the promontorium and forms posteromedial wall of the hypotympanic sinus; medial margin of petrosal overlaps basioccipital.
(3) ventral margin of the rostral tympanic process of the petrosal extends laterally and slightly dorsally, resulting in the formation of a distinct hypotympanic sinus within the rostral tympanic process itself
(4) dorsal extention of the ventral margin of the rostral tympanic process relative to state 3, resulting in lateral wall to the hypotympanic sinus within the process ,
(0) poorly defined and shallow
(1) wider and deeper than the plesiomorphic state but still relatively poorly defined
(2) deep sinus with enclosing walls that is well distinguished from remainder of auditory cavity
(3) wide and deep sinus that extends posterolaterally,
41. Squamosal epitympanic sinus
(3) wide with high posterior wall
(0) supraoccipital about as tall as it is wide
(1) supraoccipital taller than it is wide ,
44. Left and right parietal suture
(1) partially or completely co-ossified suture,
(1) sagittal crest small, not extending to frontals
(0) sagittal crest large and extending to frontals,
(2) three or more mental foramina,
(1) mesiodistally expanded and flat-crowned,
(0) occupies premaxillary-maxillary suture
(1) entirely contained within maxillary,
51. *Relative height of P2 and P3
(1) P2 and P3 subequal in height
(0) well-developed posterior cutting edge
(1) lacking posterior cutting edge,
53. *Relative height of p2 and p3
(1) p2 and p3 subequal in height
(0) present in anterior cingulum of m2-4
55. *Relative position of hypoconid to protoconid on m3
(0) buccally salient to protoconid
(2) lingual to salient protoconid,
57. Shape of narial flange of premaxilla
(1) wing-like narial processes present,
58. Position of lacrimal foramen
(1) within lacrimal-maxillary suture,
59. Supraoccipital contribution to foramen magnum
(0) contributes to superior margin of foramen magnum
(1) exoccipitals contact each other medially, excluding the supraoccipital from the foramen magnum ,
(1) mesiodistally expanded and flat-crowned,
(0) single-rooted unicuspid upper canine
(1) accessory cusps are present on either side of the major cusp of C1,
(0) lower molars are as high on the buccal side as they are on the lingual side
(1) lower molar height is significantly higher on buccal side than lingual side,
(0) metacone is directly posterior to the paracone
(1) metacone is shifted lingually in relation to the paracone,
(1) I3 is larger than I2 and I4,
65. Lingual cusp presence on lower incisors
66. Diastema between i3 and canine
(0) single-rooted unicuspid lower canine
(1) accessory cusps are present on either side of the major cusp of c1,
68. Size of upper or lower canine
(0) large, raised above premolars
(1) small, about as high as premolars,
69. *Diastema between C1 and P1
(1) short, less than the length of P1
(0) long, more than the length of P1,
71. Anterior cusp of P1 and/or P2
72. *Diastema between P1 and P2
(1) short, less than the length of p1
(2) long, more than the length of p1,
73. Morphology of the central cusp of P1 and P2
(0) central cusp is not inflated
(1) central cusp is inflated widening the tooth anteriorly,
74. *Relative length of P2 and P3
(0) P3 almost twice as long as P2
75. *Diastema between canine and p1
(1) short, less than the length of p1
(2) long, more than the length of p1,
(0) p1 is shorter than either p2 or p3
(1) p1 is as long as p2 or p3,
79. *Diastema between p1 and p2
(1) short, less than the length of p1
(2) long, more than the length of p1,
80. *Relative length of p2 and p3
(0) p3 almost twice as long as p2
(0) p3 major cusp laterally compressed
(0) the anterior and posterior roots of p3 are level
(1) the anterior root of p3 is more exposed than the posterior root, reclining p3 toward m1,
(0) StA is large, with a long crest running anteroposteriorly
(1) StA is small, with a short or no crest running anteroposteriorly
85. Morphology of StB/StC on M1
(1) StB and StC are distinct cusps
(2) StB and StC are fused and oval in shape connected by a stylar crest but clearly identifiable as separate cusp
(3) StB and StC are fused and oval in shape connected by a stylar crest but difficult to clearly identify each cusp,
86. *Relative size of StB and StC on M1
87. Connection of StB and StD on M1
(0) StB/C and StD are connected by a series of crests
(1) StB/C and StD are not connected by any crests,
89. *Termination of postprotocrista on M1
(0) postprotocrista ends on anterior flank of metacone
(1) postprotocrista ends on lingual flank of metacone
(2) postprotocrista ends on posterolingual flank of metacone
(3) postprotocrista joins with the posterior cingulum and ends posteriorly to the midpoint of the postmetacrista
(4) postprotocrista joins with the posterior cingulum and ends at the lingual flank of the metastylar tip,
90. *Position of metaconule on M1
(0) metaconule is directly positioned at the base of the metacone
(1) a small shelf is between the base of the metacone and the metaconule
(2) a larger shelf is between the base of the metacone and the metaconule,
(0) StE present as a distinct cusp
(1) remnant of StE as a stylar crest or small cusp
(1) small anterior cingulum lingual to anterior tip of tooth
(2) small anterior cingulum connected to protocone shelf by a small shelf
(3) large anterior cingulum expanded further up to the protocone,
(0) StA is large, with a long crest running anteroposteriorly
(1) StA is small, with a short or no crest running anteroposteriorly
(0) StB is oval with a stylar crest running through it
(1) StB is conical with no stylar crest connection,
96. Connection of StB and StD on M2
(0) StB and StD are connected by a series of crests
(1) StB and StD are not connected by any crests,
97. *Direction of preparacrista of M2
(0) preparacrista terminates at base of StB (no crest connection).
(1) preparacrista terminates at the tip StB.
(2) preparacrista connects to StB with a weak connection to StA
(3) preparacrista connects to parastylar tip/StA,
(0) stylar crest present on StD connects to metastylar tip
(1) StD is a conical cusp not connected to the stylar crest running to the metastylar tip
(2) StD is a conical cusp and no stylar crest present,
100. *Termination of postprotocrista on M2
(0) postprotocrista ends on anterior flank of metacone
(1) postprotocrista ends on lingual flank of metacone
(2) postprotocrista ends on posterolingual flank of metacone
(3) postprotocrista joins with the posterior cingulum and ends posteriorly to the midpoint of the postmetacrista
(4) postprotocrista joins with the posterior cingulum and ends at the lingual flank of the metastylar tip,
101. *Position of metaconule on M2
(0) metaconule is directly positioned at the base of the metacone
(1) a small shelf is between the base of the metacone and the metaconule
(2) a larger shelf is between the base of the metacone and the metaconule,
(0) StE present as a distinct cusp
(1) remnant of StE as a stylar crest
(0) StA is large, with a long crest running anteroposteriorly
(1) StA is small, with a short or no crest running anteroposteriorly
(0) StB is oval with a stylar crest running through it
(1) StB is conical with no stylar crest connection (except by wear),
107. Connection of StB and StD on M3
(0) StB and StD are connected by a series of crests
(1) StB and StD are not connected by any crests,
108. *Direction of preparacrista of M3
(0) preparacrista terminates at base of StB (no crest connection).
(1) preparacrista terminates at the tip StB.
(2) preparacrista connects a crest linking StA and StB
(3) preparacrista connects to parastylar tip/StA,
(0) stylar crest present on StD connects to metastylar tip
(1) StD is a conical cusp not connected to the stylar crest running to the metastylar tip
(2) StD is a conical cusp and no stylar crest present,
110. *Termination of postprotocrista/posthypocrista on M3
(0) postprotocrista ends on anterior flank of metacone
(1) postprotocrista ends on lingual flank of metacone
(2) postprotocrista ends on posterolingual flank of metacone
(3) postprotocrista joins with the posterior cingulum and ends posteriorly to the midpoint of the postmetacrista
(4) postprotocrista joins with the posterior cingulum and ends at the lingual flank of the metastylar tip,
111. *Position of metaconule on M3
(0) metaconule is directly positioned at the base of the metacone
(1) a small shelf is between the base of the metacone and the metaconule
(2) a larger shelf is between the base of the metacone and the metaconule,
(0) StE present as a distinct cusp
(1) remnant of StE as a stylar crest
(1) small anterior cingulum lingual to anterior tip of tooth
(2) small anterior cingulum connected to protocone shelf by a small shelf
(3) large anterior cingulum expanded further up to the protocone,
117. Connection of StA and StB on M4
(1) crest connecting the two cusps,
118. Direction of preparacrista of M4
(0) preparacrista terminates StA
(1) preparacrista terminates at the anterior flank of StB,
(1) postparacrista curves as a small centrocrista and ends just after the centrocrista,
120. *Termination of postprotocrista on M4
(0) postprotocrista ends anterior to the most posterior end of the postparacrista
(1) postprotocrista ends level with the most posterior end of the postparacrista
(2) postprotocrista ends posterior to the most posterior end of the postparacrista ,
(1) small metacone raise above the postparacrista
(2) large metacone expands the length of the tooth,
123. Presence of protocone on M4
(0) paraconid-metaconid distance is longer than metaconid-protoconid distance
(1) paraconid-metaconid distance is as long as metaconid-protoconid distance
(0) paraconid-metaconid distance is shorter than metaconid-protoconid distance
(1) paraconid highly reduced or absent,
127. Position of paraconid on m1
(0) paraconid anterior to metaconid
(1) paraconid anterobuccal to metaconid,
128. Shape of unworn entoconid on m1-3
(1) triangular (wider posteriorly than anteriorly)
129. *Cristid obliqua termination on m1
(0) cristid obliqua terminates buccal to level with the protocone
(1) cristid obliqua terminates lingual to the protocone to the midpoint to the tooth width
(2) cristid obliqua terminates lingual to the midpoint of the tooth width,
130. Posthypocristid direction on m1
(0)oblique to the tooth row axis
(1) perpendicular to the tooth row axis ,
131. Connection of posthypocristid on m1
(0) posthypocristid connects to hypoconulid
(1) posthypocristid connects to base of entoconid when the tooth is unworn, but as the tooth wears down, a crest connects the tip of the entoconid to the posthypocristid.,
132. Size of hypoconulid on m1
(0) paraconid-metaconid distance is longer than metaconid-protoconid distance
(1) paraconid-metaconid distance is as long as metaconid-protoconid distance
(2) paraconid-metaconid distance is shorter than metaconid-protoconid distance
(3) paraconid highly reduced or absent,
134. *Cristid obliqua termination on m2
(0) cristid obliqua terminates buccal to the protocone
(1) cristid obliqua terminates lingual to the protocone about midpoint to the tooth width
(2) cristid obliqua terminates lingual to the midpoint of the tooth width,
135. Posthypocristid direction on m2
(0) oblique to the tooth row axis
(1) perpendicular to the tooth row axis ,
136. Connection of posthypocristid on m2
(0) posthypocristid connects to hypoconulid
(1) posthypocristid connects to entoconid,
137. *Size of hypoconulid on m2
(0) paraconid-metaconid distance is longer than metaconid-protoconid distance
(1) paraconid-metaconid distance is as longer as metaconid-protoconid distance
(2) paraconid-metaconid distance is shorter than metaconid-protoconid distance
(3) paraconid highly reduced or absent,
139. *Cristid obliqua termination on m3
(0) cristid obliqua terminates buccal to the protocone
(1) cristid obliqua terminates lingual to the protocone about midpoint to the tooth width
(2) cristid obliqua terminates lingual to the midpoint of the tooth width,
140. Posthypocristid direction on m3
(0) oblique to the tooth row axis
(1) perpendicular to the tooth row axis ,
141. Connection of posthypocristid on m3
(0) posthypocristid connects to hypoconulid
(1) posthypocristid connects to entoconid,
142. *Size of hypoconulid on m3
(0) paraconid-metaconid distance is longer than metaconid-protoconid distance
(1) paraconid-metaconid distance is as longer as metaconid-protoconid distance
(2) paraconid-metaconid distance is shorter than metaconid-protoconid distance
(3) paraconid highly reduced or absent,
145. Preentocristid orientation on m4
147. Postentocristid orientation on m4
148. Cristid obliqua termination on m4
(0) cristid obliqua terminates lingual to the protocone about midpoint to the tooth width
(1) cristid obliqua terminates lingual to the midpoint of the tooth width,
149. *Posthypocristid direction on m4
(0) oblique to the tooth row axis
(1) perpendicular to the tooth row axis
151. *Size of hypoconulid on m4
(0) buccal shelf end at buccal side of hypoconid
(1) buccal shelf buccally reduced (hypoconid more buccal than shelf),
155. Presence of palatine fenestrae
APPENDIX 2
Nexus-formatted character matrix for the 42 taxa included in our phylogenetic analyses. Outgroup taxa were Djarthia murgonensis, Barinya wangala, Mutpuracinus archibaldi, Dasyurus hallucatus, Dasyuroides byrnei, Phascogale tapoatafa, Antechinus stuartii, Sminthopsis macroura. ? = missing data; - = inapplicable (text file for download).
K. J. Travouillon School of Earth Sciences
School of Earth Sciences
University of Queensland
St. Lucia
Queensland 4072
Australia
and School of Biological, Earth and Environmental Sciences
University of New South Wales
New South Wales 2052
Australia
Kenny J. Travouillon is a vertebrate palaeontologist and palaeoecologist working at University of Queensland, in the School of Earth Sciences. He received a Bachelor of Science (Hons) and PhD from the University of New South Wales, Sydney, Australia.
His early research aimed at clarifying the chronological position of the Riversleigh World Heritage Area (northwestern Queensland) fossil sites amongst the Tertiary of Australia, in absence of absolute dates, using multivariate analyses as a tool to group sites of similar age based on taxonomic information. His research also aimed at identifying the palaeoenvironments of Riversleigh's sites, using cenograms. Cenograms are a graphical representation of the logged body mass of mammals in a fauna. The shape of the cenogram can be use to predict whether the environment in which the fauna lives is open or closed and humid or arid.
He was recently awarded with the Robert Day Postdoctoral Fellowship at the University of Queensland, working on Peramelemorphian (bilbies and bandicoots) phylogeny and describing new taxa from Riversleigh and Etadunna (South Australia).
![]()
R.M.D. Beck School of Biological, Earth and Environmental Sciences
School of Biological, Earth and Environmental Sciences
University of New South Wales
New South Wales 2052
Australia
Robin Beck graduated with a BA (Hons) in Natural Sciences from Pembroke College, University of Cambridge, in 2002 and an MSc in Advanced Methods in Taxonomy and Biodiversity from the Natural History Musem and Imperial College, London, in 2003. He was awarded his PhD from the University of New South Wales in 2008 for his thesis 'Form, function, phylogeny and biogeography of enigmatic Australian metatherians', which was supervised by Mike Archer and Sue Hand. From 2009 to 2011, he was an NSF-funded postdoctoral research fellow at the American Museum of Natural History, working in collaboration with Rob Voss and Sharon Jansa on the morphology and phylogeny of marsupials. Since 2012, he has been a DECRA postdoctoral fellow at UNSW, studying the only known record of early Palaeogene fossil mammals in Australia, from the early Eocene Tingamarra Fauna. His major research interests are the morphology, systematics and biogeography of mammals (particularly marsupials), and combining morphological and molecular data to resolve phylogenetic relationships and divergence times. He has undertaken fieldwork in Australia, New Zealand and Antarctica.
![]()
S.J. Hand School of Biological, Earth and Environmental Sciences
School of Biological, Earth and Environmental Sciences
University of New South Wales
New South Wales 2052
Australia
Suzanne Hand is a vertebrate palaeontologist and Associate Professor at the University of New South Wales where she teaches zoology and geology. Her research interests are largely in the areas of evolutionary biology, functional morphology, phylogenetics, and biogeography, with a special interest in fossil and modern mammals, particularly bats. She co-leads research of the World Heritage-listed Riversleigh fossil deposits of northern Australia, and is also currently researching fossil mammal faunas in Europe, Asia, New Zealand and New Caledonia. She is the author of 100+ publications on Australian fossil mammals including several books.
Photo: R. Morris
![]()
M. Archer School of Biological, Earth and Environmental Sciences
School of Biological, Earth and Environmental Sciences
University of New South Wales
New South Wales 2052
Australia
Mike Archer was born in Sydney but lived in the USA until 1967 when he moved back to Australia. By age 11 he was addicted to fossil and living animals. Degrees: Princeton University BA, University of Western Australia PhD. Key positions: Professor, University of New South Wales; Director of the Australian Museum, Sydney; Dean of Science, University of New South Wales. Current research foci on which I have published (~16 books & 265 refereed journal articles): evolution of mammals, in particular monotremes and marsupials; evolution of Australia's terrestrial environments over the last 100 million years; biocorrelation of Australia's Cenozoic vertebrates; impacts of palaeoclimate change; use of palaeontological datasets to optimise conservation of living species; innovative biofuel company; conservation through sustainable use of native resources; native animals as pets; battling Creationists; dental function and ontogeny; megafaunal extinctions; biogeographic history of New Zealand; early Cenozoic biotas of Argentina; megatsunamis; science communication; lacustrine and karst geology and petrogenesis; etcetera. I started the Thylacine & Lazarus Projects to see how far we could get in efforts to recover, revitalize and enable ancient DNA to construct functional organisms. For more info: http://www.create.unsw.edu.au/team/marcher/.
![]()
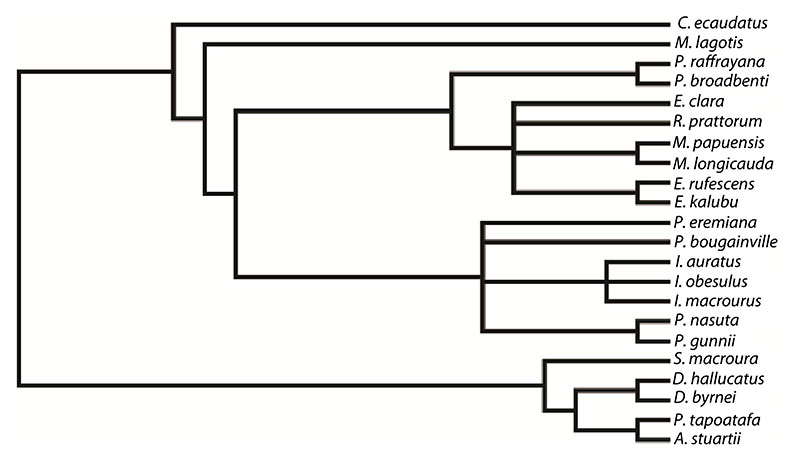
FIGURE 1.Bulungu muirheadae, sp. nov. Holotype QM F10666, 1, occlusal view of left dentary; 2, buccal view of left dentary. Paratype QM F10668, 3, occlusal view of left dentary; 4, buccal view of left dentary. Paratype QM F10669, 5, buccal view of right dentary; 6, lingual view of right dentary. Scale bars for 1-4 equals 1mm, for 5-6 equals 2mm.
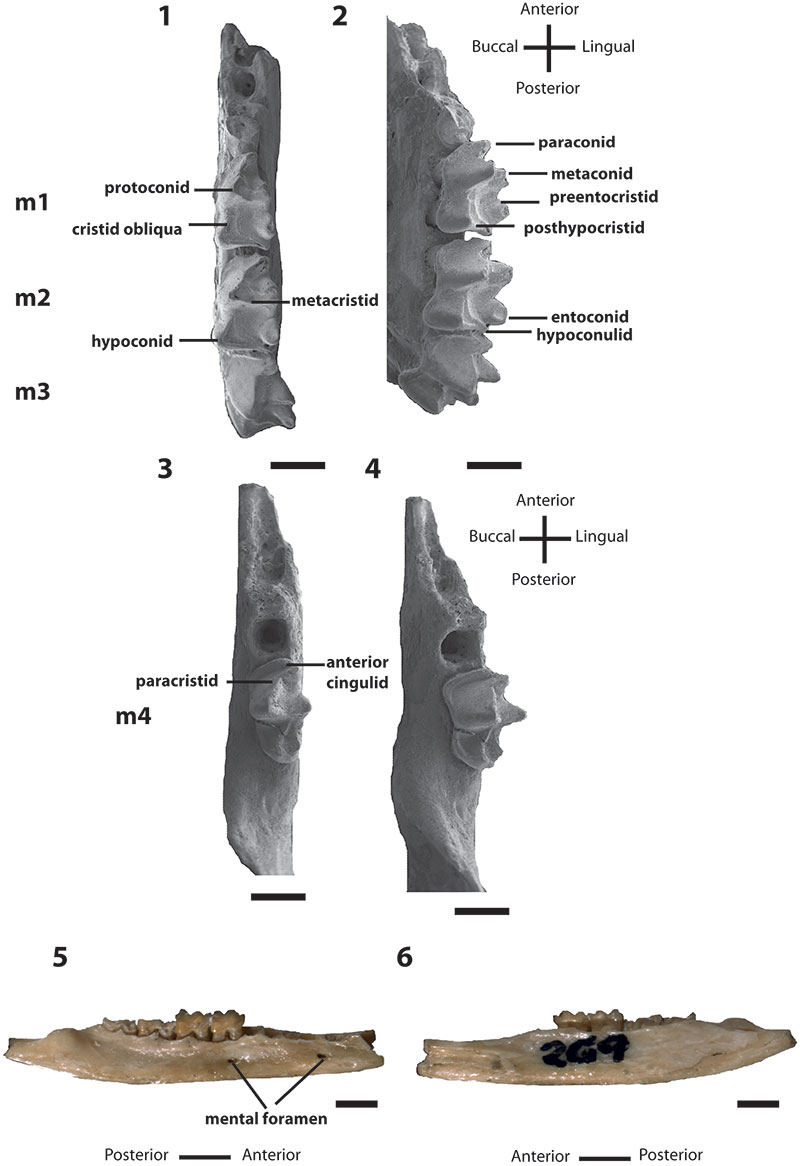
FIGURE 2.Bulungu muirheadae, sp. nov. Paratype UCR 15207, 1, occlusal view stereo pair of M1. Paratype QM F10664, 2, occlusal view stereo pair of M2. Paratype UCR 15275, 3, occlusal view stereo pair of M3. Scale bar equals 1mm.
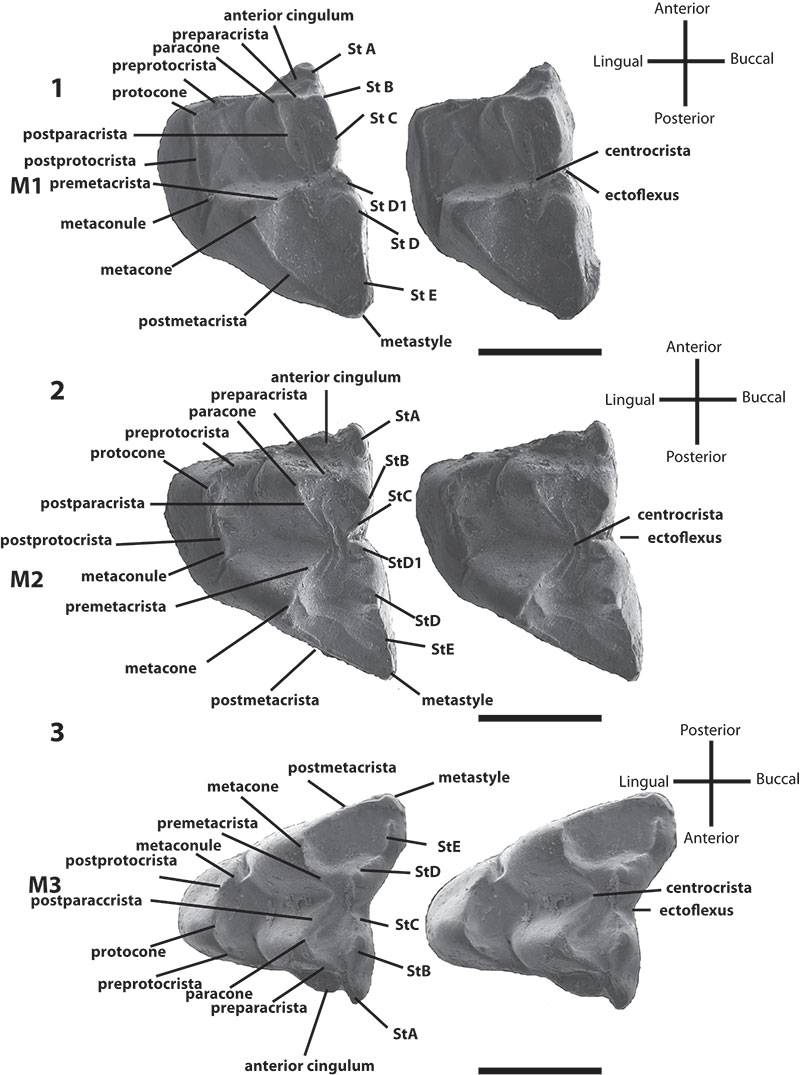
FIGURE 3.Bulungu campbelli, sp. nov. Holotype SAM P13853, 1, lingual view of m1; 2, occlusal view of m1; 3, buccal view of m1; 4, buccal view of right dentary; 5, buccal/occlusal view of right dentary; 6, occlusal view of right dentary; 7, stereo pair of occlusal view of right maxilla. Scale bar equals 1mm.
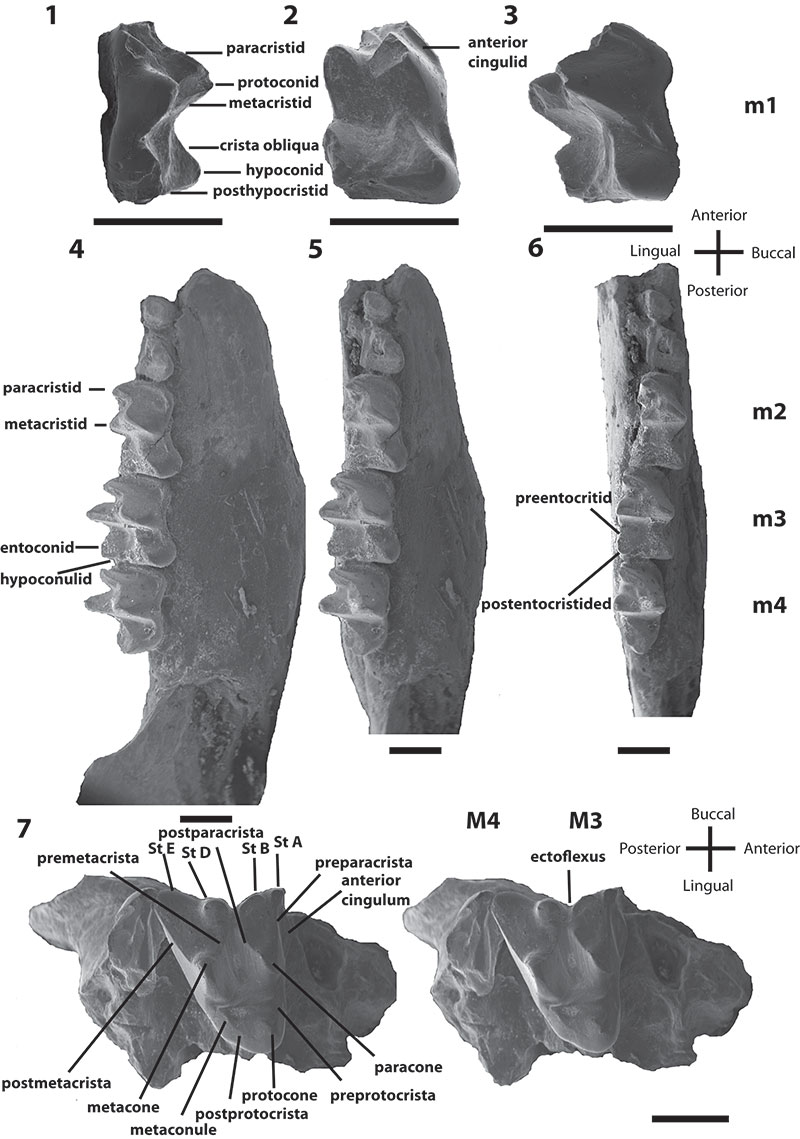
FIGURE 4. Phylogenetic relationships of Bulungu muirheadae and B. campbelli sp. nov. based on our 156 character craniodental matrix. Fossil and recently extinct taxa are indicated by †. Bulungu muirheadae and B. campbelli are highlighted in bold. Crown-group Peramelemorphia is bracketed. 1, strict consensus of 8 most parsimonious trees (tree length = 889; consistency index excluding uninformative characters = 0.3412; retention index = 0.6560) from unconstrained maximum parsimony analysis of the matrix. Numbers above branches represent bootstrap values (1000 replicates); numbers below branches represent decay indices. 2, strict consensus of 24 most parsimonious trees (tree length = 917; consistency index excluding uninformative characters = 0.3308; retention index 0.6395) that result when the matrix is analysed using maximum parsimony and enforcing a "backbone" molecular scaffold based on Westerman et al. (2012).
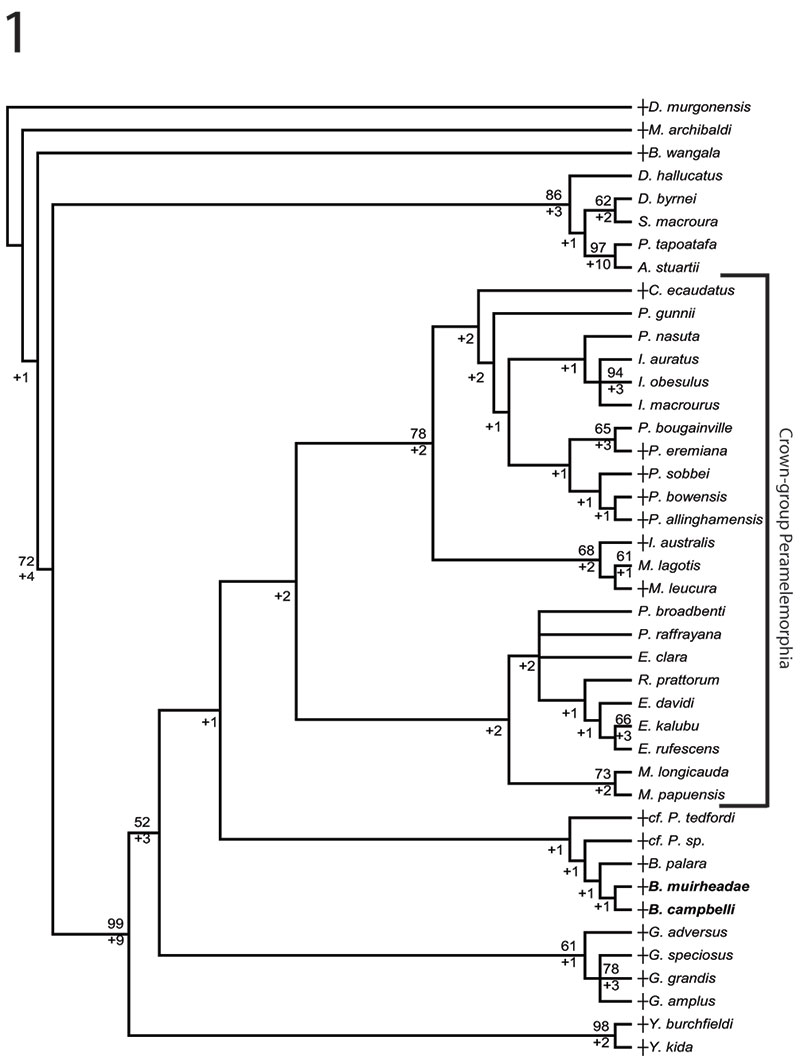
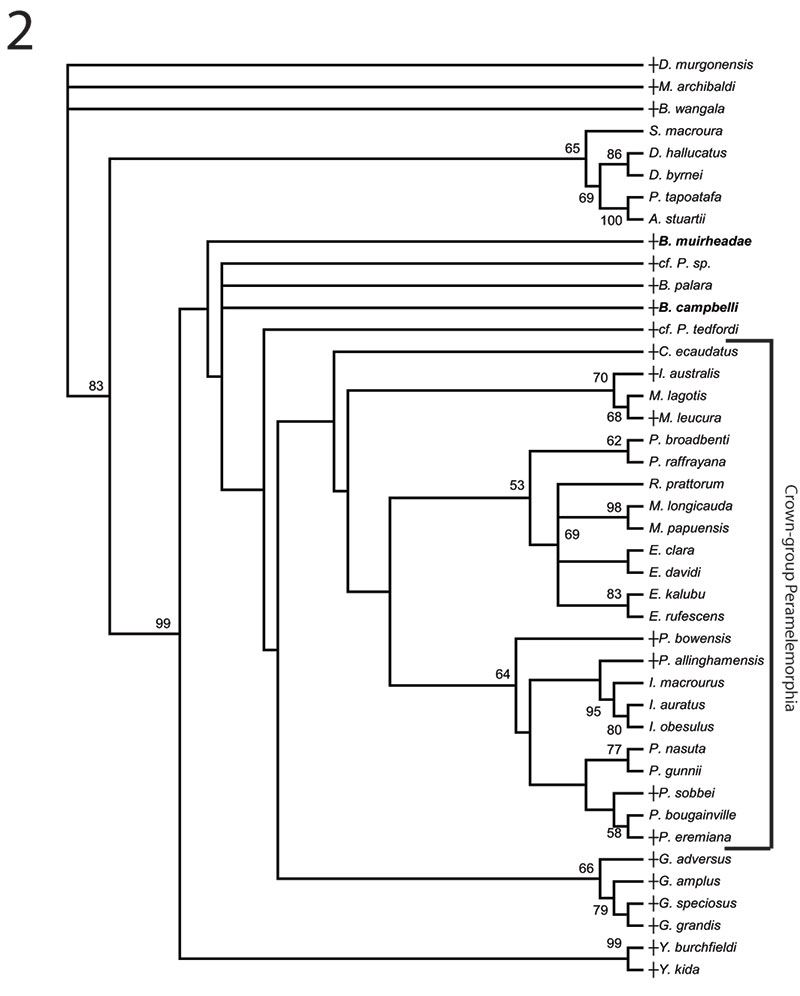
FIGURE 5. Phylogenetic relationships of Bulungu muirheadae and B. campbelli sp. nov. based on our 156 character craniodental matrix. Fossil and recently extinct taxa are indicated by †. Bulungu muirheadae and B. campbelli are highlighted in bold. Crown-group Peramelemorphia is bracketed. 1, 50% majority rule consensus that results after Bayesian analysis of the matrix assuming the Mk + G model (harmonic mean of the log likelihood of the post-burn-in trees = -2898.10). The analysis was run for five million generations, sampling trees every 100 generations, with a "burn-in" period of 1.25 million generations. Numbers above branches represent Bayesian posterior probabilities. 2, 50% majority rule consensus that results after Bayesian analysis of the matrix is analysed enforcing a "backbone" molecular scaffold based on Westerman et al. (2012), assuming the Mk + G model (harmonic mean of the log likelihood of the post-burn-in trees = -2933.45). The analysis was run for five million generations, sampling trees every 100 generations, with a "burn-in" period of 1.25 million generations. Numbers above branches represent Bayesian posterior probabilities.
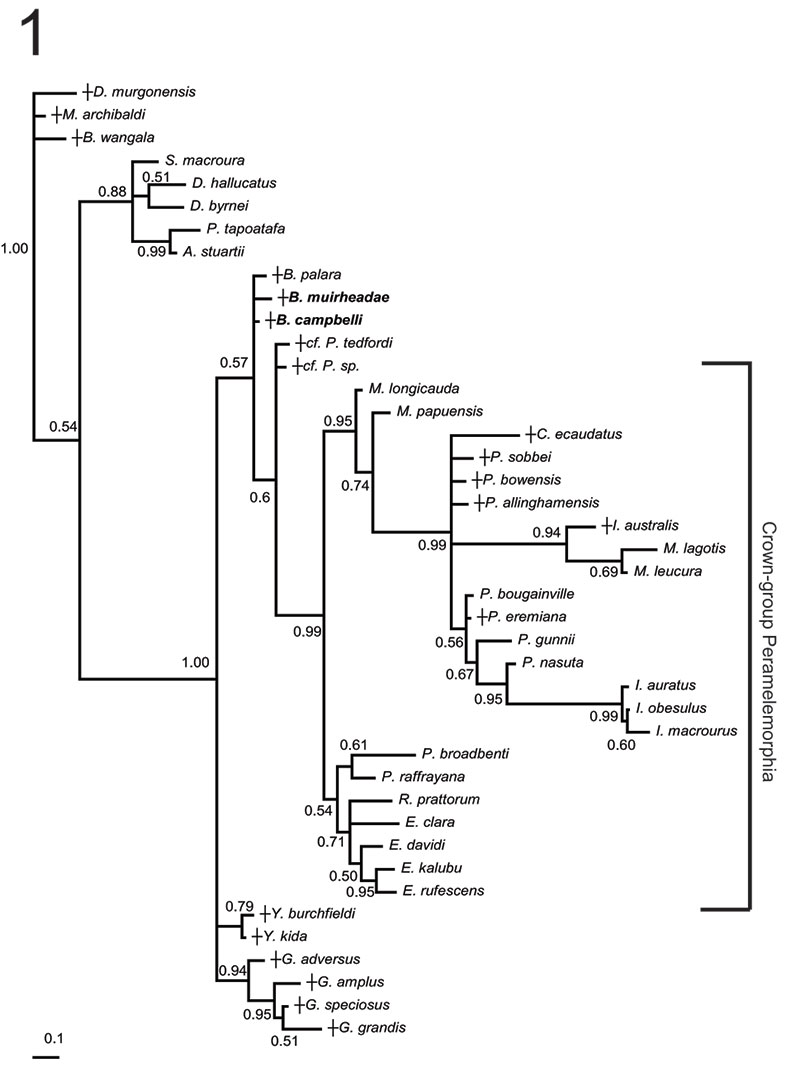
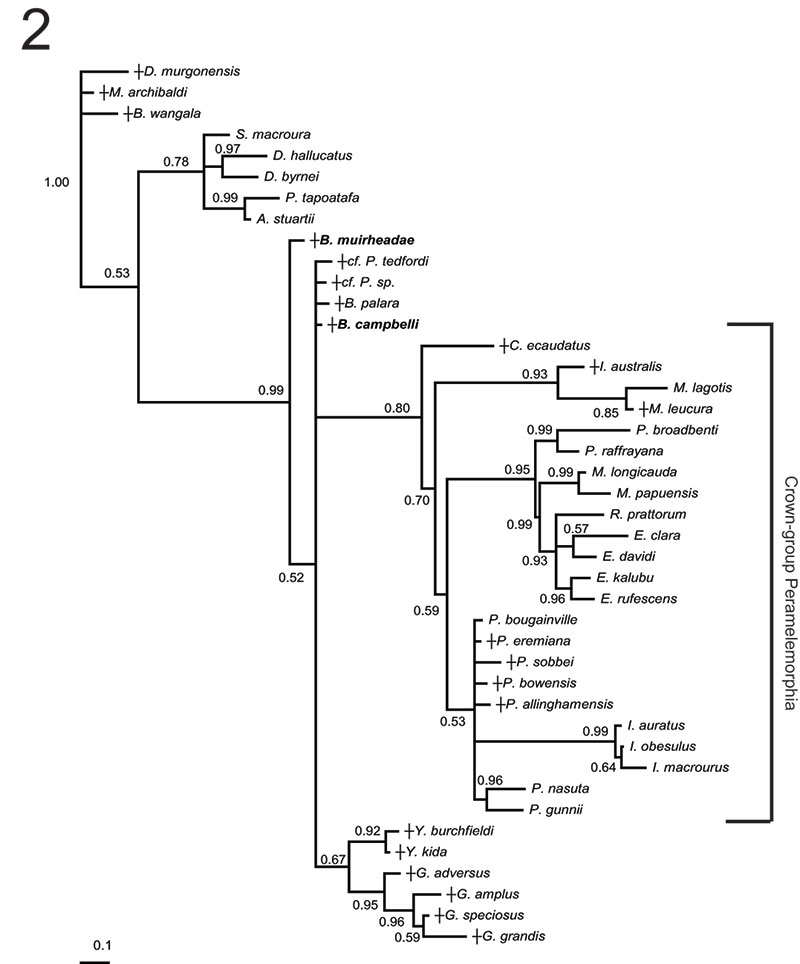
TABLE 1. Measurements of the upper dentition of type and referred material in mm; body mass estimates in grams. L – anteroposterior length, W = lingual-buccal width, M = molar. Measurements of damaged/broken teeth are in brackets.
|
Taxa |
Specimen |
Locality |
M1 |
M2 |
M3 |
Body mass |
|||
|
L |
W |
L |
W |
L |
W |
estimate |
|||
|
Bulungu muirheadae |
QM F10662 |
Tedford Locality |
|
|
|
|
2.21 |
2.12 |
211.45 |
|
Bulungu muirheadae |
QM F10664 |
Tedford Locality |
|
|
2.22 |
1.57 |
|
|
129.49 |
|
Bulungu muirheadae |
QM F10665 |
Tedford Locality |
|
|
|
|
1.88 |
1.83 |
131.95 |
|
Bulungu muirheadae |
QM F12416 |
Tedford Locality |
2.12 |
1.84 |
|
|
|
|
171.8 |
|
Bulungu muirheadae |
QM F12417 |
Tedford Locality |
|
|
|
|
2.05 |
1.86 |
154.38 |
|
Bulungu muirheadae |
QM F12420 |
Tedford Locality |
|
|
1.88 |
1.47 |
|
|
90 |
|
Bulungu muirheadae |
UCMP 108063 |
Tedford Locality |
|
|
|
|
2.12 |
1.98 |
178.78 |
|
Bulungu muirheadae |
UCR 15270 |
Tedford Locality |
|
|
|
|
2.08 |
1.7 |
137.59 |
|
Bulungu muirheadae |
UCR 15275 |
Tedford Locality |
|
|
|
|
1.89 |
1.76 |
125.32 |
|
Bulungu muirheadae |
UCR 15285 |
Tedford Locality |
|
|
|
|
2 |
1.75 |
135.45 |
|
Bulungu muirheadae |
UCR 15299 |
Tedford Locality |
|
|
1.86 |
1.45 |
|
|
86.62 |
|
Bulungu muirheadae |
UCR 15307 |
Tedford Locality |
1.91 |
1.5 |
|
|
|
|
106.22 |
|
Bulungu muirheadae |
UCR15332 |
Tedford Locality |
|
|
(1.83) |
1.57 |
|
|
n/a |
|
Bulungu muirheadae |
UCR 16996 |
Tedford Locality |
1.88 |
(1.24) |
|
|
|
|
n/a |
|
Bulungu campbelli |
SAM P13853 |
Ngapakaldi Quarry |
|
|
|
|
1.7 |
1.75 |
105.68 |
TABLE 2. Measurements of the lower dentition of type and referred material in mms; body mass estimates in grams. L – anteroposterior length, AW = anterior width, PW = posterior width, m = molar.
|
Taxa |
Specimen |
Locality |
m1 |
m2 |
m3 |
m4 |
Body mass estimate |
||||
|
L |
W |
L |
W |
L |
W |
L |
W |
||||
|
Bulungu muirheadae |
QM F10661 |
Tedford Locality |
1.57 |
1.12 |
|
|
|
|
|
|
109.34 |
|
Bulungu muirheadae |
QM F10666 |
Tedford Locality |
1.71 |
1 |
1.84 |
1.14 |
1.95 |
1.19 |
|
|
127.57 |
|
Bulungu muirheadae |
QM F10667 |
Tedford Locality |
|
|
1.67 |
1.25 |
1.89 |
1.26 |
- |
1.02 |
132.31 |
|
Bulungu muirheadae |
QM F10668 |
Tedford Locality |
|
|
|
|
|
|
2.17 |
1.17 |
186.66 |
|
Bulungu muirheadae |
QM F10669 |
Tedford Locality |
1.7 |
1.09 |
1.76 |
1.24 |
|
|
|
|
118.22 |
|
Bulungu muirheadae |
QM F12414 |
Tedford Locality |
|
|
|
|
1.9 |
1.23 |
|
|
128.85 |
|
Bulungu muirheadae |
QM F12415 |
Tedford Locality |
|
|
|
|
2.17 |
1.38 |
|
|
182.68 |
|
Bulungu muirheadae |
QM F12418 |
Tedford Locality |
|
|
|
|
2.1 |
1.28 |
|
|
156.9 |
|
Bulungu muirheadae |
QM F12419 |
Tedford Locality |
1.73 |
1.15 |
|
|
|
|
|
|
124.59 |
|
Bulungu muirheadae |
UCR 15266 |
Tedford Locality |
|
|
|
|
|
|
2 |
1.03 |
142.1 |
|
Bulungu muirheadae |
UCR 15268 |
Tedford Locality |
1.91 |
1.17 |
|
|
|
|
|
|
147.14 |
|
Bulungu muirheadae |
UCR 15279 |
Tedford Locality |
|
|
1.79 |
1.12 |
|
|
|
|
104.66 |
|
Bulungu muirheadae |
UCR 15301 |
Tedford Locality |
|
|
1.76 |
1.18 |
|
|
|
|
110.1 |
|
Bulungu muirheadae |
UCR 15306 |
Tedford Locality |
|
|
|
|
1.89 |
1.21 |
|
|
124.97 |
|
Bulungu muirheadae |
UCR 15318 |
Tedford Locality |
|
|
1.78 |
1.24 |
|
|
|
|
120.16 |
|
Bulungu muirheadae |
UCR 17002 |
Tedford Locality |
|
|
|
|
2.2 |
1.5 |
|
|
209.44 |
|
Bulungu campbelli |
SAMP 13853 |
Ngapakaldi Quarry |
|
|
1.79 |
1.11 |
1.75 |
1.32 |
1.79 |
1.09 |
132.38 |
|
Bulungu campbelli |
SAMP 13853 |
Ngapakaldi Quarry |
1.55 |
1.07 |
|
|
|
|
|
|
100.56 |
The oldest fossil record of bandicoots (Marsupialia; Peramelemorphia) from the late Oligocene of Australia
Plain Language Abstract
Two new species of bandicoots (marsupials) are described from fossil deposits in South Australia, aged between 26.1 and 23.6 million years old. These bandicoots are currently the oldest fossils from this group. The two species are quite small, under 250 grams and principally fed on insects. Our analysis of relationships within this group shows that these two species are related to a species from the Riversleigh World Heritage Area (WHA), Queensland, and the Kutjamarpu Local Fauna in South Australia.
Resumen en Español
El registro fósil más antiguo de bandicuts (Marsupialia; Peramelemorphia), del Oligoceno tardío de Australia
Se describen dos nuevos representantes del Oligoceno tardío de marsupiales del orden Peramelemorphia (bandicuts y bilbis) procedentes de la Formación Etadunna (Australia Meridional). Bulungu muirheadae sp. nov., de la Zona B (Fauna Local [FL] de Ditjimanka) está representada por varios dentarios y molares superiores e inferiores aislados. Bulungu campbelli sp. nov., de la Zona C (FL de Ngapakaldi), está representada únicamente por un dentario y una maxila. Ambas especies constituyen los registros más antiguos de bandicuts descritos hasta al momento. Las dos son de pequeño tamaño (masa corporal estimada <250 gramos) en comparación con la mayoría de especies actuales de bandicuts y, de acuerdo con su morfología dental, probablemente eran insectívoras. Aparentemente se trata de formas congéneres de Bulungu palara de las faunas locales miocenas de Riversleigh (Queensland) y de Kutjamarpu (Formación Wipajiri, Australia Meridional). No obstante, el peramelemorfo de la Zona B parece ser más plesiomórfico que B. palara dado que retiene centrocristas completas en todos los molares superiores.
Palabras clave: Peramelemorfo; Oligoceno; Australia Meridional; bandicut; nueva especie; marsupiales
Traducción: Miguel Company
Résumé en Français
Le plus ancien registre fossile de bandicoots (marsupiaux; Peramelemorphia) de l'Oligocène supérieur d'Australie
Deux nouveaux représentants de l'ordre marsupiale Peramelemorphia (bandicoots et bilbies), provenant de la Formation d'Etadunna dans l'état de l'Australie du Sud, datant de l'Oligocène supérieur, sont décrits ici. Bulungu muirheadae sp. nov., provenant de la zone B (Faune Locale de Ditjimanka [LF]), est représenté par plusieurs dentaires et molaires supérieures et inférieures isolées. Bulungu campbelli sp. nov., provenant de la zone C (Ngapakaldi LF), est représenté par un seul dentaire et maxillaire. Ensemble, ils représentent les plus anciens fossiles de bandicoots décrits à ce jour. Ils sont tous les deux de petite taille (masse corporelle estimée <250 grammes) par rapport à la plupart des espèces vivantes de bandicoot et d'après leur morphologie dentaire, ils étaient probablement insectivores. Ils semblent appartenir au même genre que Bulungu palara, présent dans les faunes de Riversleigh, site du patrimoine mondial (WHA), au Queensland, et dans Kutjamarpu LF (Formation de Wipajiri), en Australie du Sud, datant du Miocene. Cependant, le péramélémorphien de la zone B semble être plus plésiomorphe que B. palara à cause de la rétention de complète centrocristae sur toutes les molaires supérieures.
Mots-clés: Péramélémorphien; Oligocène; Australie du Sud; bandicoot, nouvelles espèces; marsupiaux
Translator: Author
Deutsche Zusammenfassung
Der älteste Fossilnachweis von Beuteldachsen (Marsupialia; Peramelemorphia) aus dem späten Oligozän von Australien
Es werden hier zwei spätoligozäne Vertreter der Beuteltierordnung Peramelemorphia (Beuteldachse und Bilbies) aus der Etadunna Formation von Südaustralien beschrieben. Bulungu muirheadae sp. nov. aus Zone B (Ditjimanka Lokale Fauna [LF]) ist mit einigen Unterkiefern und isolierten unteren und oberen Molaren repräsentiert. Bulungu campbelli sp. nov. aus Zone C (Ngapakaldi LF) ist mit einem einzigen Unterkiefer und einer Maxilla repräsentiert. Zusammen stellen sie die ältesten fossilen beschriebenen Beuteldachse dar. Beide sind klein (angenommenes Körpergewicht <250 Gramm) im Vergleich zu den meisten heute lebenden Beuteldachsen und waren ihrer Gebissmorphologie nach wahrscheinlich Insektenfresser. Sie scheinen mit Bulungu palara aus miozänen Lokalfaunen der Riversleigh World Heritage Area (WHA), Queensland und der Kutjamarpu LF (Wipajiri Formation) von Südaustralien kongenerisch zu sein. Allerdings scheint der Peramelemorphe aus Zone B plesiomorpher zu sein, da er komplette Centrocristae auf allen unteren Molaren beibehalten hat.
SCHLÜSSELWÖRTER: Peramelemorphe; Oligozän; Südaustralien; Beuteldachs; neue Art; Beuteltiere
Translators: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024