

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
FIGURE 1. Libonectes morgani skull in dorsal view. Sq – squamosal; stf – supratemporal fenestra; pt – pterygoid; pc – parietal crest; j – jugal; tb – temporal bar; soc – supraoccipital; p – parietal; po – postorbital; o – orbit; pfr – prefrontal; en – external nares; mx – maxilla; pmx – premaxilla.
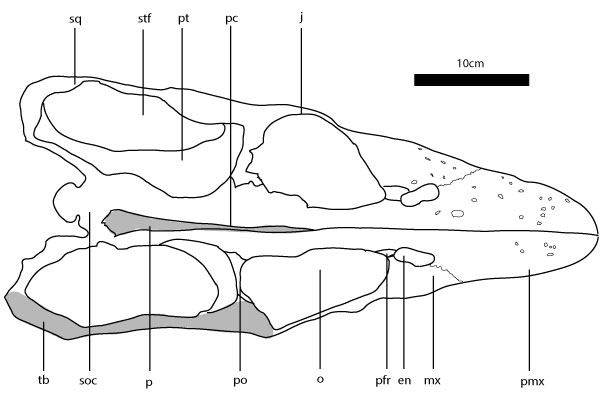
FIGURE 2. Workflow from laser scanning to finite-element analysis. 1 The base model was laser scanned using Scanstudio®. 2 The 3-D modeling used NURBS spline surfaces to conform with the shape of the laser-scanned skull; this is done in Lightwave®. 3 The NURBS surfaces facilitate remodeling according variants intended to be studied. 4 The models are meshed in the FEA software COMSOL Multiphysics®. 5 In COMSOL Multiphysics® the boundary conditions (loadings and constraints) are applied. 6 The results obtained are scaled to the same range in order to facilitate comparison.

FIGURE 3. Base skull model in various views with the boundary conditions. 1 Lateral view. 2 Dorsal view. 3 Ventral view. 4 Anterodorsal view. 5 Laterodorsal view. Boundary conditions: light blue - area for origin of left MAMES origin, dark blue - area for origin of right MAMES origin; light yellow – area for origin of right MPst origin; dark yellow – area for origin of right MPst origin; light green – area for origin of right MAMP origin; dark green – area for origin of right MAMP origin; light orange – area for origin of left MPt origin; dark orange – area for origin of right MPt origin.
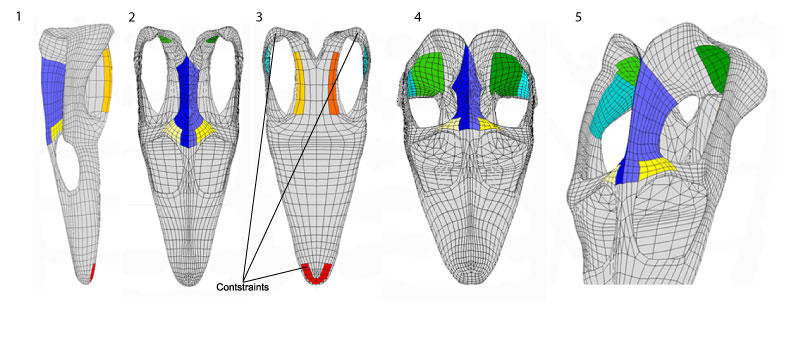
FIGURE 4. Osteological correlates in the cranium of Libonectes morgani. 1 Periosteum texture on the premaxilla-maxilla for textural comparison with other regions of the skull, note the ornamented surface of the bone and the presence of several elongated foramina. 2 Medial view of the temporal bar at the level of the squamosal jugal suture; note the deeply striated and rugose texture of the bone where the mAMES most likely inserted. 3 In anterodorsal view, the posteriormost portion of the temporal fenestra; note excavated form of this section of the skull where the mAMP most likely inserted. 4. Dorsal view of the palate in the orbital region for textural comparison of the periosteum; note the smooth surface of the bone. 5 The highly pitted and rugose texture of the parietal crest is suggestive of the origination of the mAMEM and mAMEP. 6 Finely pitted surface of the posterior surface of the postorbital where the mPST most likely inserted. 7 Finely pitted lateral walls of the pterygoid where most likely the mPT inserted. Scale bar equals 0.5 cm.

FIGURE 5. Osteological correlates in the mandible of Libonectes morgani. 1 Lateral view of the anterior portion of the mandible; the smooth but slightly ornamented texture of this section of the bone contrasts with the areas where muscles insert. 2 In medial view near the coronoid the bone texture is highly but smoothly pitted suggestion the area of insertion of the mPST. 3 Medial view of the mandible posterior to the coronoid; note the finely pitted area, tentatively the insertion of the mAMP. 4 Ventral view of the mandible with fine ornamental striations (contrast with the striations of the mPT insertion). 5 Strongly remodeled bone periosteum presenting a wavy pattern where a portion of the mPT inserted. 6 Dorsolateral view of the mandible near the coronoid; presumable mAMES insertion demonstrating deeply pitted bone texture. 7 Lateral view of the mandible demonstrating deep striations for presumable insertion of the mPT. 8 Dorsal view of the glenoid region with deeply pitted surface possibly for the insertion of the mAME.

FIGURE 6. Muscle attachments in Libonectes morgani, Sphenodon puncatus and Alligator mississipiensis. 1 Libonectes morgani skull in laterodosal view and its adductor chamber musculature origins. 2 Inset of the Libonectes morgani skull in posterodorsal view depicting the supratemporal fenestra and adductor chamber musculature origins. 3 Sphenodonpunctatus skull in laterodorsal view and its adductor chamber musculature origins based on Jones et al. (2009). 4 Alligator mississipiensis skull in ventrolateral view depicting the adductor chamber region and its musculature origins. Grayish dark blue: mAMEM; Dark blue: mAMEP; Light blue: mAMES; Light green: mAMP; Gray: mDM - musculus depressor mandibulae; Orange: mPt; Yellow: mPstP; Pink: mPstS.
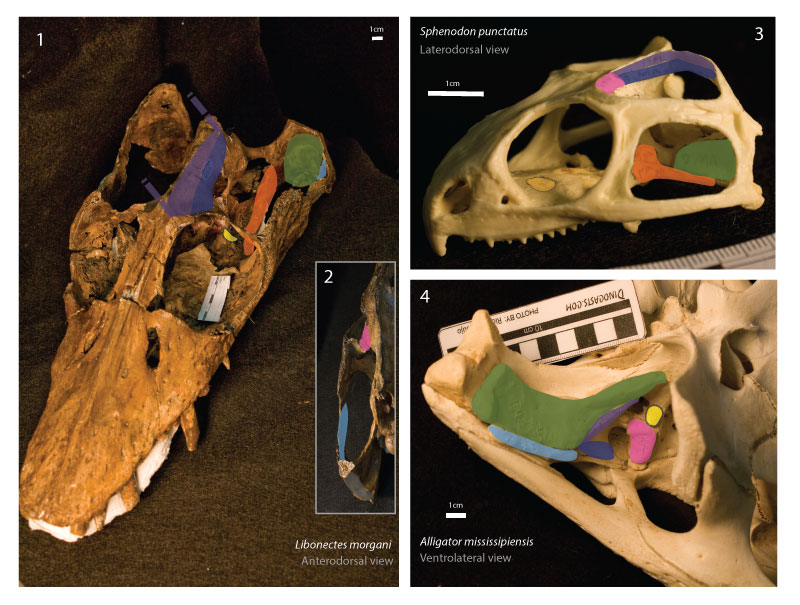
FIGURE 7. Mandible insertions in Libonectes morganiSphenodon puncatus and Alligator mississipiensis. 1 Libonectes morgani medial view of the posterior portion of the mandible. 2 Sphenodonpunctatus medial view of the posterior portion of the mandible. 3 A. mississipiensis medial view of the posterior portion of the mandible. Purple: mAMEM; Dark blue: mAMEP; Light blue: mAMES; Light green: mAMP; Gray: mDM - musculus depressor mandibulae; Orange: mPt; Yellow: mPstP; Pink: mPstS.
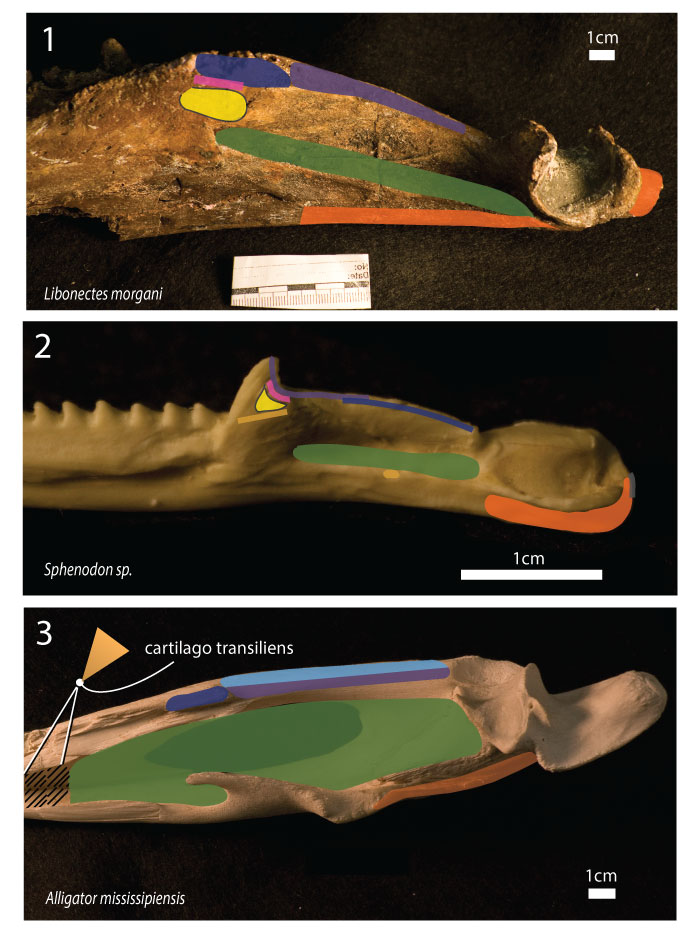
FIGURE 8. Finite-element models results. STB – shallow temporal bar variant model; LPC – low parietal crest variant model; SSF – small supratemporal fenestra variant model. Strain energy density, results in dorsal view: 1 Base model, 2 low parietal crest model, 3 shallow temporal bar model, 4 small supratemporal fenestra model. In lateral view: 5 Base model, 6 low parietal crest model, 7 shallow temporal bar model, 8 small supratemporal fenestra model. Von Mises stresses, results in dorsal view: 9 Base model, 10 low parietal crest model, 11 shallow temporal bar model, 12 small supratemporal fenestra model. In lateral view: 13 Base model, 14 low parietal crest model, 15 shallow temporal bar model, 16 small supratemporal fenestra model. Strain energy density, results in anterodorsal view:n17 Base model, 18 low parietal crest model, 19 shallow temporal bar model, 20 small supratemporal fenestra model. Von Mises stresses, results in anterodorsal view: 21 Base model, 22 low parietal crest model, 23 shallow temporal bar model, 24 small supratemporal fenestra model.

FIGURE 9.Libonectes morgani base model with posterior part of the skull viewed anterodorsally. 1 Results for the model with scaled forces at pterygoid (LC1). 2 Results for the model with no pterygoid (LC2).
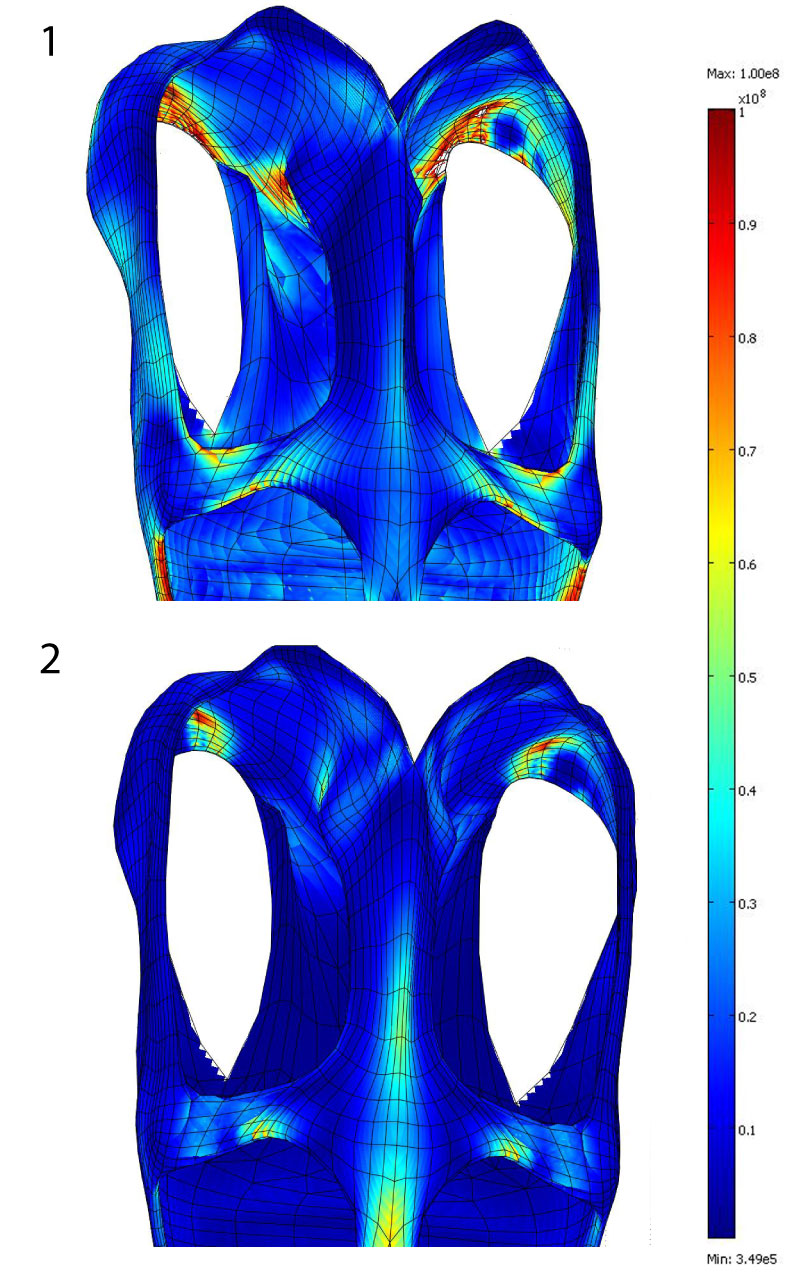
TABLE 1. Areas for the muscles origins in the different variant models.
|
Area (cm^2) |
||||||
|
mPST |
mAME |
mAMP |
mAMES |
mDM |
mPT |
|
|
Initial model |
11.3213 |
58.3881 |
29.4737 |
43.5531 |
13.1584 |
24.3531 |
|
Low parietal |
9.09129 |
34.8869 |
37.6998 |
45.681 |
13.1584 |
21.0722 |
|
Thin temporal |
9.87854 |
52.74 |
37.2582 |
28.9364 |
18.912 |
22.0872 |
|
Small supratemporal |
11.2411 |
31.9708 |
36.9917 |
27.8797 |
23.2992 |
13.3696 |
TABLE 2. Angles for the orientation of the different forces produced by the muscles in all models.
|
Angles between forces components (°) |
||||||
|
Dorsoventral and mediolateral axis |
25 |
23 |
0 |
0 |
0 |
35 |
|
Anteroposterior and mediolateral axis |
0 |
0 |
0 |
0 |
0 |
70 |
|
Dorsoventral and anteroposterior axis |
0 |
0 |
31 |
0 |
0 |
36 |
TABLE 3. Morphometric data of different skull measurements in various eosauropterygians, and extant phylogenetically-relevant taxa used for comparisons and contextualization.
|
Higher-rank clade |
Taxon |
Specimen |
Data Source |
Total length of |
Supratemporal |
Maximum height of the |
Adductor Chamber |
|
Sauropterygia |
Libonectes morgani |
SMUSMP 69120 |
Measured with ruler |
45 |
17 |
8.5 |
19 |
|
Sauropterygia |
Callawayasaurus columbensis |
UCMP 38349 |
O'Keefe 2001 |
15.8 |
5.8 |
2.9 |
4.56 |
|
Sauropterygia |
Rhomaleosaurus zetlandicus |
YORYM G503 |
Taylor 1992 |
91.1 |
27.23 |
4.4 |
17.3 |
|
Sauropterygia |
Nothosaurus winterswijktensis |
NMNHL St 445530, NMNHL St 445913 |
Albers and Rieppel 2003 |
131.8 |
46.7 |
? |
? |
|
Sauropterygia |
Nothosaurus haasi |
HUJ-Pal 2250 |
Rieppel et al 1999 |
119.2 |
38.8 |
4.8 |
13.8 |
|
Sauropterygia |
Placochelys placodonta |
FAFI Ob/2323/Vt.3 |
Rieppel 2001 |
156.8 |
74.4 |
14.9 |
47.8 |
|
Sauropterygia |
Cyamodus rostratus |
UMO BT 748 |
Rieppel 2001 |
118.6 |
57.9 |
14.31 |
64.48 |
|
Sauropterygia |
Henodus chelyops |
Specimen I and II |
Rieppel 2001 |
145.6 |
62.5 |
? |
50.2 |
|
Sauropterygia |
Macroplacus raeticus |
BSP 1967 I 324 |
Rieppel 2001 |
191.1 |
96.9 |
24.6 |
69.8 |
|
Sauropterygia |
Protenodontosaurus italicus |
MFSN 1819GP |
Rieppel 2001 |
156 |
63.6 |
15.5 |
74.5 |
|
Crocodilyformes |
Alligator mississipensis |
SMU Uncatalogued |
Measured with ruler and calliper |
450 |
28.08 |
13.69 |
133.88 |
|
Chelonia |
Chelydra serpentina |
SMU R-160 |
Measured |
106.83 |
64.32 |
13.82 |
39.38 |
|
Chelonia |
Testudo sp. |
SMU Uncatalogued |
Measured from cast |
120.98 |
66.13 |
14.73 |
54.39 |
|
Chelonia |
Chelonia mydas |
SMU Uncatalogued |
Measured |
205.65 |
54.48 |
38.82 |
95.81 |
|
Chelonia |
Lepidochelys olivacea |
SMU Uncatalogued |
Measured from cast |
157.6 |
37.58 |
29.55 |
69.44 |
|
Varanidae |
Varanus |
SMU Uncatalogued |
Measured from cast with ruler and calliper |
226 |
49.34 |
14.06 |
63.85 |
|
Rhyncocephalia |
Sphenodon punctatus |
SMU Uncatalogued |
Measured from cast |
63.96 |
21.13 |
7.04 |
27.52 |
|
Serpentes |
Python |
SMU Uncatalogued |
Measured |
134.52 |
63.52 |
16.92 |
28.93 |
|
Lacertidae |
Cyclura cornuta |
SMU Uncatalogued |
Measured from cast |
132.39 |
38.24 |
21.22 |
57.5 |
|
Mosasauria |
Tylosaurus |
SMU Uncatalogued |
Measured from cast with ruler |
150 |
33 |
8 |
33 |
|
Mosasauria |
Platecarpus |
SMU Uncatalogued |
Measured from cast with ruler and calliper |
370 |
95.86 |
19.48 |
90.69 |
Ricardo Araújo Huffington Department of Earth Sciences
Huffington Department of Earth Sciences
Southern Methodist University
Dallas, Texas 75275
USA
and Museu da Lourinhã
Rua João Luís de Moura, 95
2530-158 Lourinhã
Portugal
Ricardo Araújo is currently completing his doctoral degree at the Southern Methodist University. The doctoral dissertation focuses on Angolan plesiosaurs, functional morphology, and myological reconstruction in plesiosaurs. He is part of the PaleoAngola project, an international collaborative endeavor that aims to study and collect the vast Angolan paleontological heritage. Ricardo is also a co-leader of the PalNiassa project, a Portuguese-Mozambican cooperation that aims to trigger research and increase dynamism in Mozambican academia, by means of an invaluably important, diverse and undiscovered fossil record. Ricardo also actively collaborates with the Museu da Lourinhã, Portugal, on dinosaur eggs and embryos.
![]()
Michael J. Polcyn Huffington Department of Earth Sciences
Huffington Department of Earth Sciences
Southern Methodist University
Dallas, Texas 75275
USA
Michael J. Polcyn is Director of the Digital Earth Laboratory, Lecturer and Research Associate in the Roy M. Huffington Department of Earth Sciences at Southern Methodist University. His current research interests include the systematics of mosasauroids and adaptations in secondarily aquatic tetrapods. Research also includes application of technology to problems in paleontology.
A biomechanical analysis of the skull and adductor chamber muscles in the Late Cretaceous Plesiosaur Libonectes
Plain Language Abstract
The skeletons of animals can be computer modeled and tested to elucidate their mechanical properties. Those mechanical properties can then be used to infer certain function or capabilities. In this paper, we vary the geometry of a computer modeled skull of a well-known plesiosaur species (Libonectes morgani) in order to understand how they affect stress distribution in skull of different structure. As a starting point we describe the bones and how they are connected to one-another, and examine the muscles to determine how they connect to the bones and the strength of the forces they produce. We use the bones to create a geometric computer model and use the data from the muscles to determine the simulated direction of magnitude of the modeled forces. We show that some variations in the skull of elasmosaurid plesiosaurs are to increase strength correlated with changes to accommodate more space for jaw closing muscles, and thus increased bite force. We also tested and reject the hypothesis that the skull in plesiosaurs worked in a similar fashion to crocodiles, using a quick bite for capturing prey and strong muscles for holding the prey. Instead we find that the skull of Libonectes is arranged to accommodate quick bites only.
Abstract in Portuguese
Uma análise biomecânica do crânio e músculos adductors mandibulares no plesiossauro Libonectes do Cretácico superior
Os plesiossauros são um diversificado grupo de répteis marinhos que existiram durante cerca de três quartos do Mesozóico (do Jurássico inferior ao Cretácico superior). Os plesiossauros exibem variações na morfologia do crânio e coluna cervical que presumivelmente estão relacionadas com diferenças funcionais dos hábitos de predação. Contudo, de um ponto de vista biomecânico, estas criaturas marinhas têm um tipo de organização craniana partilhada por alguns répteis: a condição neodiapsídea. Apesar disso, aspectos estruturais básicos nalguns clados, como os elasmossaurídeos, continuam mal compreendidos, nomeadamente a presença de uma ampla fenestra supratemporal, uma arcada temporal alta, e uma crista parietal também alta. Estes aspectos representam compromissos biomecânicos com implicações para os hábitos de predação. Aqui, testamos hipóteses relacionadas com a estrutura e mecânica do crânio do plesiossauro elasmossaurídeo Libonectes morgani do Cretácico superior do Texas (EUA). Usando análise de elementos finitos e cargas baseadas numa reconstrução detalhada dos músculos aductores, estimamos os valores de tensão e extensão para o crânio do Libonectes. Também modelámos digitalmente diversas variantes do crânio do Libonectes, de modo a compreender o papel da morfologia na performance mecânica do crânio (e.g., altura da arcada temporal). Os nossos resultados demonstram que uma secção fisiológica maior dos músculos aductores é conseguida através de uma fenestra supratemporal alargargada apesar disso afectar a performance mecânica do crânio, efeito que é compensado através da resistência adicional dada por uma crista parietal e arcada temporal altos, na situação em que a carga é simétrica, as componentes laterais da força compensam a componente vertical das forças actuantes. Este arranjo craniano também aumenta o comprimento dos músculos aductores e portanto a sua massa muscular total. Propomos ainda que a reduzida crista do pterigóide é indicativa do relativamente insignificante papel do músculo pterigóide, reflectindo uma alteração da maioria da força de aducção da mandíbula para os músculos: aductor mandibular externo, pseudotemporal, e aductor mandibular posterior. A redução dos músculos pterigóides falsifica a hipótese do sistema aductor dual, que postula que o sistema de inércia cinética e de pressão estática coexistem.
PALAVRAS-CHAVE: modelação de elementos finitos; Plesiosauria; Libonectes morgani; mecânica de predação; muscular trade-off; músculos adductores mandibulares
Translator: author
Resumen en Español
Análisis biomecánico del cráneo y los músculos de la cámara aductora en el plesiosauro del Cretácico tardío Libonectes
Los plesiosauros fueron un clado de reptiles marinos que vivieron desde el inicio del Jurásico hasta el final del Cretácico. Muestran una amplia variación en la morfología de la cabeza y el cuello supuestamente relacionada con diferentes hábitos alimentarios. Desde un punto de vista biomecánico estos animales marinos tenían una organización craneal compartida con otros clados de reptiles: la condición neodiápsida. No obstante, todavía no se conoce en detalle la función de algunas características estructurales básicas presentes en algunos clados derivados, como los elasmosáuridos, concretamente las grandes fenestras supratemporales, las barras temporales elevadas y las altas crestas parietales. Estos rasgos representan compromisos biomecánicos con implicaciones paleobiológicas con respecto a los hábitos alimentarios. En este trabajo contrastamos distintas hipótesis específicas relativas a la estructura y mecánica del cráneo en el plesiosauro elasmosáurido Libonectes morgani del Cretácico tardío de Texas (EE. UU.). Mediante la utilización de vectores de cargas y de análisis de elementos finitos basados en una reconstrucción detallada de la musculatura de la cámara aductora, hemos obtenido estimaciones de las distribuciones de esfuerzos y deformaciones en el cráneo de Libonectes. También hemos elaborado digitalmente distintas variantes anatómicas del cráneo de Libonectes con objeto de evaluar el papel de esos rasgos (p. ej., la altura de la barra temporal) en la conducta mecánica del cráneo. Los resultados obtenidos muestran que una fenestra supratemporal agrandada proporciona una mayor área a la sección transversal fisiológica de los músculos aductores y, aunque las prestaciones mecánicas del cráneo quedan reducidas, se compensan con una mayor fuerza debida a la mayor altura de la cresta parietal y la barra temporal, ya que la carga se reparte de forma más simétrica y los componentes laterales dan lugar a un vector de fuerza vertical. Esta disposición aumenta también la longitud de la musculatura aductora y, por tanto, la masa muscular total. Sugerimos que la reducción del reborde pterigoide indicaría un papel menos importante del músculo pterigoideo y un traslado de la mayor parte de la fuerza de la mordida a los músculos pseudotemporales y aductores externos y posteriores de la mandíbula. La reducción del pterigoideo refuta la hipótesis del sistema aductor dual en la que coexisten inercia cinética y presión estática.
PALABRAS CLAVE: modelo de elementos finitos; Plesiosauria; Libonectes morgani; mecanismos de alimentación; compensación muscular; cámara aductora
Traducción: Miguel Company
Résumé en Français
Une analyse biomécanique du crâne et la chambre des muscles adducteurs chez le Plésiosaure crétacé supérieur Libonectes
Les plésiosaures étaient un clade diversifié de reptiles marins qui a subsisté pendant près des trois-quarts de Mésozoïque (du début du Jurassique jusqu'à la fin du Crétacé). Ils montrent des variations dans ma morphologie de la tête et du cou qui sont vraisemblablement liées à des différences fonctionnelles des modes de nutrition. Toutefois, d'un point de vue biomécanique, ces créatures marines ont une organisation crânienne que peu d'autres clades de reptiles possèdent: la condition néodiapside. Néanmoins, les caractéristiques structurales de base chez certains clades dérivés, tels que les elasmosauridés, restent mal compris, notamment la présence de grandes supratemporal fenestrae, de grandes barres temporales, et d'une crête pariétale haute. Ces caractéristiques présentent des compromis biomécaniques avec des implications paléobiologiques concernant le mode de nutrition. Nous testons ici des hypothèses spécifiques concernant la structure et la mécanique du crâne chez le plésiosaure elasmosauridé Libonectes morgani du Crétacé supérieur du Texas (USA). Utilisant une analyse des éléments finis et une reconstruction détaillée de la répartition des charges sur la musculature de la chambre des adducteurs, nous fournissons des estimations de la distribution du stress et de la pression pour le crâne de Libonectes. Nous avons aussi formé numériquement différentes variantes anatomiques du crâne de Libonectes dans le but de comprendre le rôle des ces traits dans la performance mécanique du crane (e.g., hauteur de la barre temporale). Nos résultats montrent qu'une section physiologique plus large des muscles adducteurs est obtenue par un élargissement des fenêtres supra-temporales qui, bien que cela réduise les performances mécaniques du crâne, est contrebalancé par une force accrue due à la grande crête pariétale et la barre temporale, la charge étant principalement symétrique, les composantes latérales fournissent un vecteur de force verticale équilibré. Cet arrangement augmente aussi la longueur de la musculature des adducteurs et donc la masse musculaire totale. Nous proposons que le processus ptérygoïde réduit indique un rôle moindre des muscles ptérygoïdiens, reflétant un transfert de la majeure partie de la force de morsure sur les muscles adducteurs externes, pseudotemporaux, et adducteurs postérieurs de la mandibule. La réduction des ptérygoïdiens facilite l'hypothèse du système d'adducteurs opposés, dans laquelle l'inertie cinétique et la pression statique coexistent.
Mots clés : modèle d'éléments finis ; Plesiosauria ; Libonectes morgani; mécanique alimentaire ; compensation musculaire ; chambre des adducteurs.
Translator: Olivier Maridet
Deutsche Zusammenfassung
Eine biomechanische Analyse des Schädels und der Adduktorenmuskulatur des Plesiosauriers Libonectes aus der späten Kreide
Plesiosaurier waren eine diverse Klade mariner Reptilien, deren Auftreten beinahe drei Viertel des Mesozoikums umfasste (vom frühesten Jura bis zur spätesten Kreide). Sie variieren in der Kopf - und Halsmorphologie, was möglicherweise mit funktionellen Unterschieden bei den Fressgewohnheiten zusammenhängt. Allerdings haben diese marinen Lebewesen vom biomechanischen Standpunkt aus gesehen eine kraniale Organisation, die sie mit ganz wenigen Kladen teilen: die neodiapside Kondition. Nichtsdestotrotz bleiben basale Strukturen in einigen abgeleiteten Kladen wie Elasmosauriden wenig verstanden, nämlich das Vorhandensein eines großen Supratemporalfensters, hohe Temporalbögen und hohe Parietalkämme. Diese Merkmale stellen biomechanische Kompromisse mit paläobiologischen Implikationen in Bezug auf die Fressgewohnheiten dar. Wir testen hier spezifische Hypothesen in Bezug auf Schädelstruktur und Mechanik beim elasmosauriden Plesiosaurier Libonectes morgani aus der späten Kreide von Texas (USA). Indem wir Finite-Element-Analyse und Loading basierend auf einer detaillierten Rekonstruktion der Adduktorenmuskulatur benutzen, liefern wir Schätzwerte der Spannungs- und Dehnungsverteilung für den Schädel von Libonectes. Ebenso morphten wir digital verschiedene anatomische Varianten des Schädels von Libonectes, um die Rolle dieser Eigenschaften in Bezug auf die mechanische Leistung des Schädels einzuschätzen. Unsere Ergebnisse zeigen, dass ein größerer physiologischer Querschnitt der Adduktorenmuskulatur durch ein vergrößertes Supratemporalfenster erreicht wird. Obwohl dadurch die mechanische Leistungsfähigkeit des Schädels reduziert wird, wird dies durch einen stärker belastbaren höheren Parieltalkamm und Temporalbogen kompensiert, vorausgesetzt die Belastung ist weitgehend symmetrisch. Dadurch wirken die lateralen Komponenten ausgleichend und ergeben einen vertikalen Kraftvektor. Dieses Arrangement erhöht ebenso die Länge der Adduktorenmuskulatur und daher die totale Muskelmasse. Wir postulieren, dass der reduzierte Pterygoidflansch eine verkleinerte Rolle des Pterygoidmuskels anzeigt, was eine Verlagerung der Hauptbeisskraft zum M. adductor mandibulae externus, M. pseudotemporalis und M. addctor mandibulae posterior reflektiert. Die Reduktion des Pterygoideus widerlegt die Hypothese eines dualen Adduktorensystems bei der kinetische Trägheit und statischer Druck koexistieren.
SCHLÜSSELWÖRTER: Finite-Element-Modellierung; Plesiosauria; Libonectes morgani; Ernährungsmechanik; muskulärer Ausgleich; Adduktorenkammer
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024