

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
 Enrico Borghi
Enrico Borghi
Società Reggiana di Scienze Naturali
Via Tosti
1 42100 Reggio Emilia
Italy
e.borghi@corghi.com
Enrico Borghi graduated (engineering) from the Bologna University, Italy, in 1978 and became member of the Società Reggiana di Scienze Naturali in 1980. His primary interests are the fossil echinoids from the Mediterranean area, their palaeoecology and relationships with extant populations.

 Vittorio Garilli (corresponding author)
Vittorio Garilli (corresponding author)
Paleosofia-APEMA Research & Educational Service
Via Alla Falconara 34 90136 Palermo
Italy
vittoriogarilli@apema.eu, vittorio.garilli@paleosofia.it
Vittorio is a PhD palaeontologist with special interest in Cenozoic molluscs. He is always open to various research fields (from foraminifers to dinosaurs!) when involving palaeoecology and palaeo(bio)geography. His main work area is the Mediterranean Sea, trying to understand the Quaternary climate and biodiversity changes occurred in this basin. After working at the University of Palermo and at the Muséum d’Histoire Naturelle (Paris), he co-founded Paleosofia-APEMA, which is engaged in the fields of research, conservancy and education.
 Sergio Bonomo
Sergio Bonomo
Istituto per l’Ambiente Marino Costiero
Consiglio Nazionale delle Ricerche
Calata Porta di Massa
Interno Porto di Napoli
80133 Naples
Italy
sergio.bonomo@cnr.it
Since 2011, Sergio Bonomo is a researcher at the Institute for Coastal Marine Environment of National Research Council, where he studies living and fossil Coccolithophyceae in water samples and marine sediments. Sergio graduated from the Università degli Studi di Palermo and PhD from the Università degli Studi di Napoli “Federico II”. Most of his research deals with various aspects of the biostratigraphy, palaeoecology and palaeoceanography in the Mediterranean region using calcareous nannofossils.
FIGURE 1. Location map (1.1) of the Plio-Pleistocene echinoid-bearing deposits cited in the text and simplified geo-structural sketch (1.2) of NE Sicily (from Antonioli et al., 2006 and Arisco et al., 2006, modified). Capo Milazzo sites: 1, Punta Lazzi; 2, Cala S. Antonio; 3-4, Lighthouse area; 5, Punta Mazza; 6, Bacedasco (Piacenza) and Salsomaggiore (Parma); 7, Quattro Castella and Castellarano (Reggio Emilia); 8, Cianca and Fossetta (Modena); 9, Vescona quarry near Asciano (Siena); 10, Chiusi (Arezzo); 11, Anzio (Rome); 12, Ponte Calderaro and Monte Torre (Catanzaro); 13, Lazzaro and Croce Valanidi (Reggio Calabria); 14, Salice, Coilare and Contrada Petrazza (Messina); 15, Ficarazzi (Palermo).
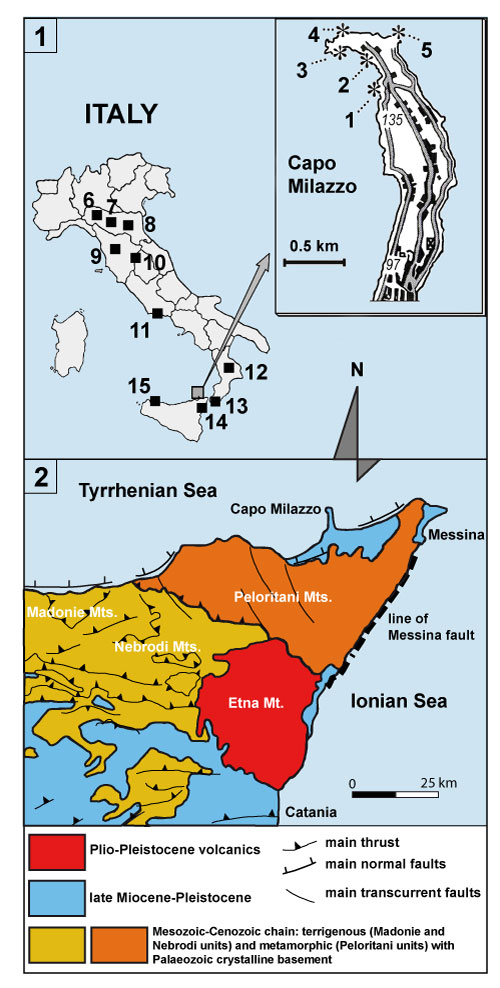
FIGURE 2. Simplified evolution of the Punta Mazza section (Capo Milazzo, NE Sicily, Italy) with stratigraphy by means of calcareous nannofossils (this study) and Violanti (1988).
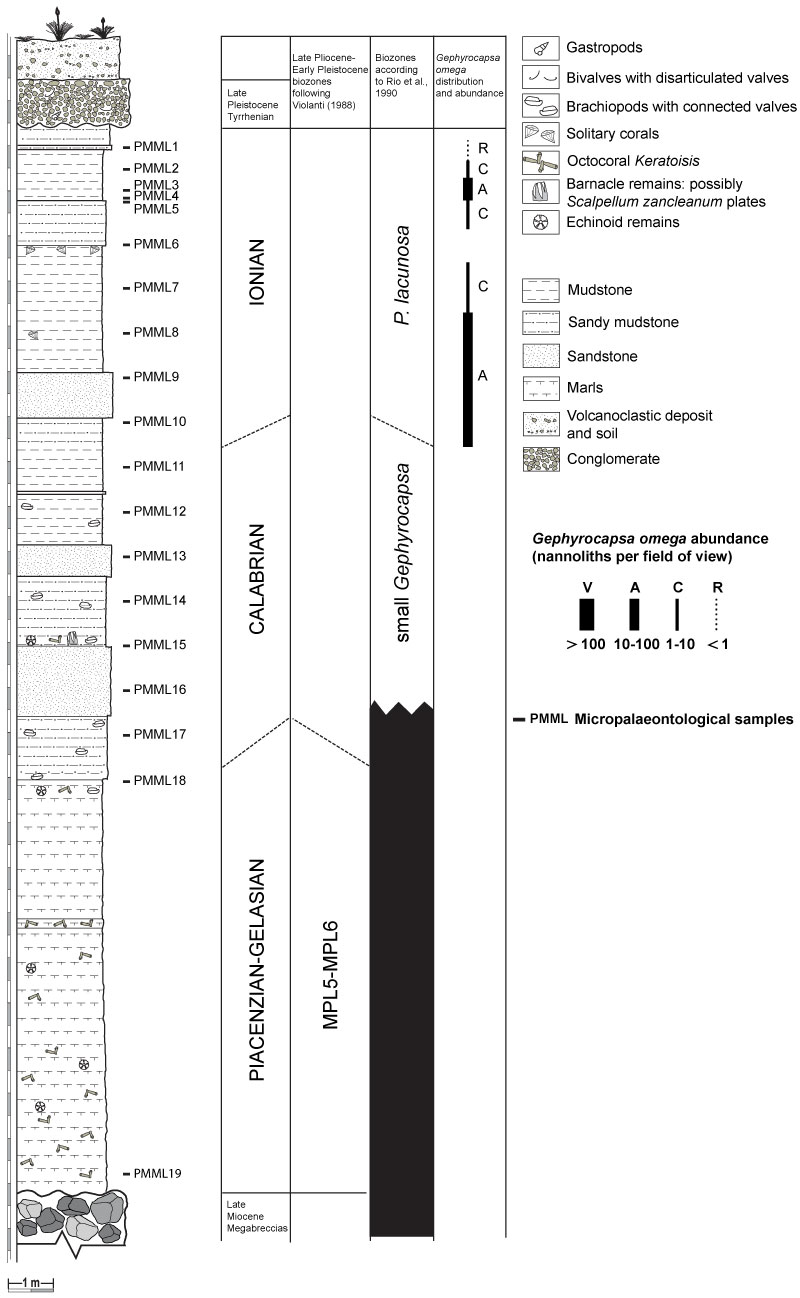
FIGURE 3. The Punta Mazza section at Capo Milazzo (NE Sicily, Italy). MM, late Miocene Megabreccias; PP, Pliocene-Early Pleistocene (Gelasian) marls, following Violanti (1988); PC, Pleistocene, Calabrian; PI, Pleistocene, Ionian; PT, Late Pleistocene (Tyrrhenian) conglomerate; PTVS, post Tyrrhenian volcaniclastic deposit and soil.

FIGURE 4. Species of the genus Cidaris Leske, 1778 from the Plio-Pleistocene of Capo Milazzo (4.1-7, 4.10-16, 4.19, 4.24, 4.27-28, 4.30-32) and Italy mainland (4.17-18, 4.20-23, 4.25-26, 4.29), and from the Recent site off Gallipoli, Ionian Sea (4.8-9). Cidaris margaritifera (Meneghini, 1862) from the uppermost Piacenzian-Gelasian and Calabrian of Capo Milazzo. 4.1, lateral view of a complete segment (MG.1033.39) with only the first fully developed adapical tubercles bearing crenulation, W=13 mm; Cala S. Antonio; 4.2, oblique view of a complete segment (MG.1033.41) with only the two last adoral plates confluent; W=25 mm, Punta Lazzi; 4.3, complete, fusiform spine (MG.1035.31) with almost smooth ridges, L=43 mm, Punta Lazzi; 4.4, complete, fusiform, non-crenulate spine (MG.1035.32) with fine spinules, L=52 mm, S. Antonio; 4.5, non-crenulate spine (MG.1032.68) with small granules, L=22 mm, Punta Lazzi; 4.6, cylindrical spine (MG.1032.69) with almost smooth ridges, L=24 mm, S. Antonio; 4.7, cylindrical spine (MG.1035.33) with low ridges bearing fine spinules; base partially crenulate, L=50 mm, S. Antonio; 4.10, close up of a spine (MG.1032.53) with smooth ridges, Ds=1.7 mm, S. Antonio; 4.11, close up of a spine (MG.1032.54) with low and sharp spinules, Ds=2.4 mm, S. Antonio; 4.12, spine (MG.1032.55) with granules, Ds=2.5 mm, S. Antonio; 4.13, crenulate spine (MG.1032.57) with ridges bearing small granules, Ds=4.2 mm, S. Antonio; 4.14, spine (MG.1032.58) with prominent spinules, Ds=4.8 mm, S. Antonio; 4.15, spine (MG.1032.62) with low spinules and space between rows covered by spongy coat, and close up view (4.16) of the non-crenulate acetabulum rim, Ds=4.8 mm, S. Antonio; 4.19 , close up view of a poriferous area (MG.1032.06), Punta Lazzi; 4.24, section of a primary spine (MG.1032.38), Ds=4.6 mm, Punta Lazzi; 4.27, fusiform, partially crenulate spine (MG1032.51) with spongy coat still visible between ridges; Ds=5.8 mm, Punta Lazzi; 4.28, short and stout primary spine (MG.1033.38); L=17 mm, Punta Lazzi. Cidaris cidaris (Linnaeus, 1778), complete test, D=42 mm (MG.1033.36); Recent, Gallipoli (Puglia), depth: 55 m. 4.8, adoral view; 4.9, close up of a crenulate adapical tubercle. Cidaris margaritifera (Meneghini, 1862) from Italy mainland. 4.17, frontal view of a complete segment, H=46 mm, Tmax=4 mm at ambitus (MG.1032.01), Calabrian of Lazzaro (Reggio Calabria), 4.18, lateral view; 4.20, primary spine; Ds=3.9 mm; Pliocene, Fossetta (Modena), Landi collection (IPUM); 4.21 , primary spine with rounded grains, Ds= 4.2 mm, Pliocene, Cianca (Modena), Landi collection (IPUM), and close up view of the smooth acetabulum rim (4.22) ; 4.23, close up of a spine with prominent spinules, Ds=4.3 mm (MG.1078.02), Piacenzian, Quattro Castella (Reggio Emilia); 4.25, primary spine with thorns and close up view of the partially crenulate acetabulum (4.26), Ds=3.7 mm (MG.1034.04), early Pliocene, Vescona quarry near Asciano (Siena); 4.29, interambulacral plate, W=14.8 mm, T=4 mm (MG.1034.05), early Pliocene, Chiusi (Arezzo), with lateral view (4.30). Cidaris cerullii (Checchia Rispoli, 1923), (MG.1033.34), Gelasian of Punta Lazzi. 4.31, test fragment, H=36 mm; with 4.32, close up of the ambulacral area. Scale bars equal 2 mm.
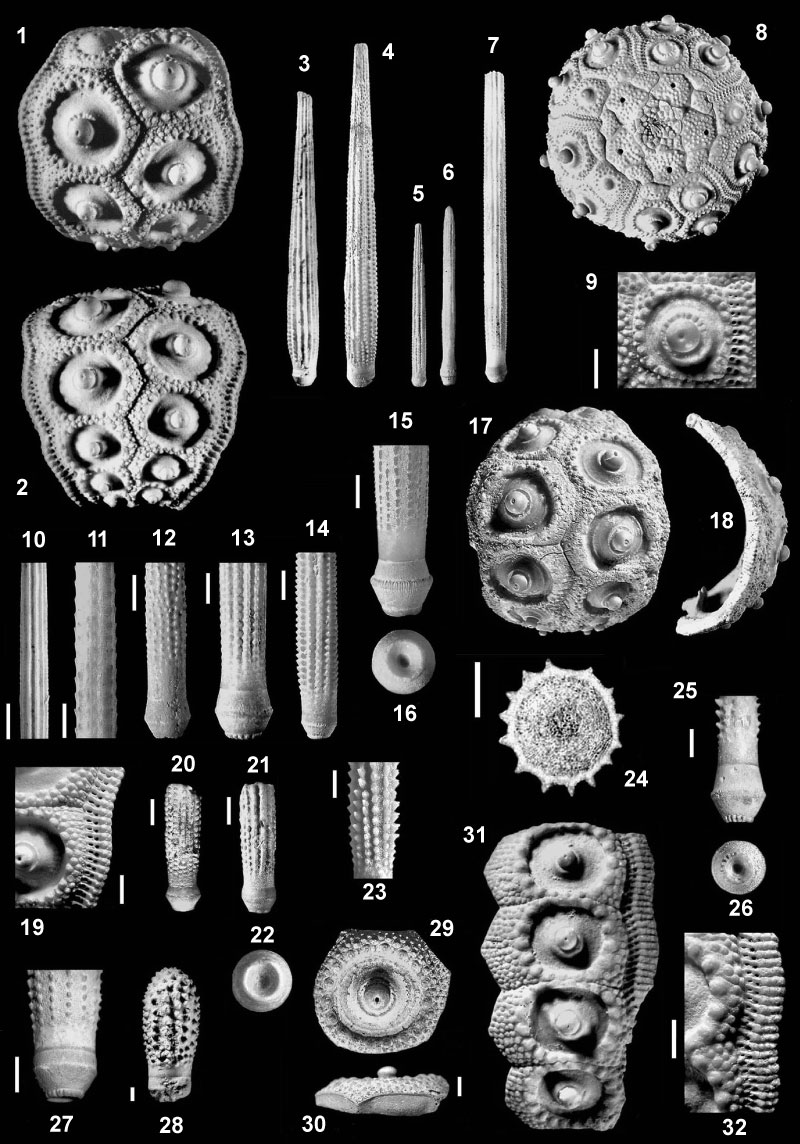
FIGURE 5. Plio-Pleistocene species of the genus Histocidaris Mortensen, 1903 from Capo Milazzo (5.1-3, 5.6-13, 5.16) and from Italy mainland (5.4-5, 5.14-15). Histocidaris sicula Borghi, 1999; uppermost Piacenzian-Gelasian of Cala S. Antonio. 5.1, holotype (MG.1073), W=27 mm; 5.2, interambulacral plate with the adjoining ambulacral plates, (MG.1033.42), W=12 mm; 5.3, cross section of a primary spine (MG.1033.06), Ds=2.6 mm; 5.6, close up view of the hollow tip of a primary spine (MG.1033.33), Ds=2.7 mm; 5.7, primary spine (MG.1035.08) with smooth shaft, the arrow marks the edge of the collar, 2 mm long, Ds=2.5 mm; 5.8, primary spine (MG.1074) with low ridges and crenulate acetabulum rim (5.9), the arrow marks the edge of the collar, which is 5.8 mm long, Ds=2.8 mm; 5.10 , adoral primary spine (MG.1033.09), L=12 mm; 5.11, adoral primary spine (MG.1077), L=11 mm, with lateral view (5.12); 5.13 , complete primary spine (MG.1035.01), L=128 mm; 5.16, adapical interambulacral plates (MG.1033.32), W=22 mm. Histocidaris rosaria (Bronn, 1831). 5.4, primary spine with sharp spinules (MG.1034.02), Ds=3 mm, Piacenzian, Campore quarry, Salsomaggiore (Parma); 5.5, primary spine with prominent thorns (MG.1034.03), Ds=4 mm, Piacenzian, Campore quarry, Salsomaggiore; 5.14, thorny adoral primary spine (MG.1034.04), L=14 mm, Piacenzian, Campore quarry, Salsomaggiore; 5.15 , tip of a primary spine, Ds=2.5 mm (MG.1034.01), early Pliocene, Vescona quarry, Asciano (Siena). Scale bars equal 2 mm.

FIGURE 6. Stirechinus scillae Desor, 1856 from the Plio-Pleistocene of Sicily. 6.1-3, complete test (MG.1034.10), D=33.5 mm, Early Pleistocene of Contrada Coilare (Messina), showing apical (6.1), lateral (6.2) and oral (6.3) views. 6.4, close up of the ridges linking the primary tubercles (MG.1034.18); 6.5, close up view of a pore-zone (MG.1034.19); 6.6-7, primary spines, L=20 mm (MG.1086.01-02); 6.8, close up view of an interambulacral scrobiculate primary tubercle, lacking ridges. All from the very late Piacenzian-Gelasian of Punta Lazzi. 6.9-10, apical (6.9) and lateral (6.10) views of a complete juvenile test (MG.1035.22), D=6.4 mm, very late Piacenzian-Gelasian of Punta Mazza; 6.11, lateral view of a complete corona, D=54 mm (MG.1034.18), very late Piacenzian-Gelasian of Punta Lazzi; 6.12, scheme of the apical system taken from a specimen of the Checchia Rispoli collection (MGUP); figured in Checchia Rispoli, 1916, plate 27, figure 6. Scale bars equal 2 mm.
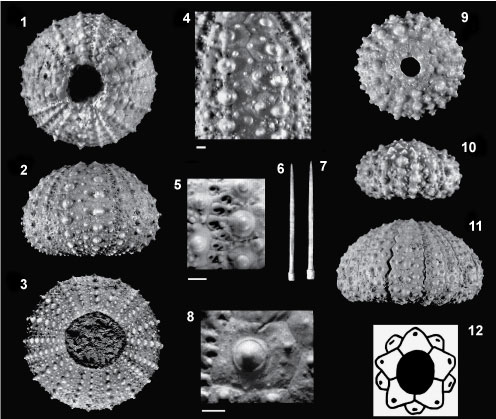
FIGURE 7. Spatangoid species from the Pliocene of north Italy. 7.1, aboral view of Schizaster braidensis Botto Micca, 1896, L=29 mm (Dip.Te.Ris IV.G60), Zanclean-Piacenzian of Genova. 7.2-3, aboral view of Holanthus ovatus (Sismonda, 1842), Pliocene of San Ruffino (Reggio Emilia), with the enlarged scheme of the plating structure (7.2) from a compete test, L=21 mm, MG.1034.06 (7.3). Scale bar equals 1 cm.
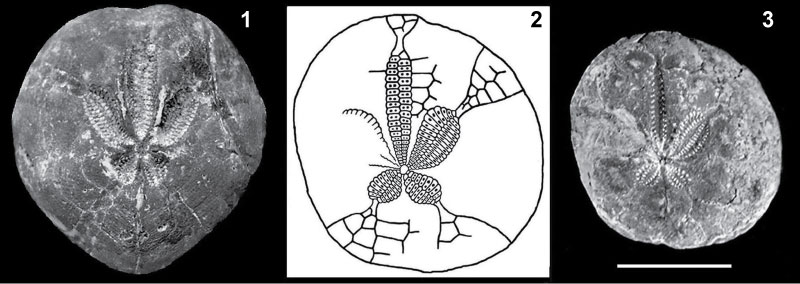
TABLE 1. Echinoids recorded in the Mediterranean Pliocene bathyal deposits and their occurrence throughout Mediterranean Miocene to Early Pleistocene. †, extinct; ?, doubtful records. Bathymetric ranges of extant populations are from Mortensen (1928, 1943), Tortonese (1965, 1977) and Phelan (1970).
TABLE 2. Echinoids recorded in the present Mediterranean deep waters (below 250/300 m depth), their bathymetric and Atlantic geographical distribution (Kohler, 1927; Tortonese, 1965, 1977).
TABLE 3. Stirechinus scillae, Desor, 1856: measurements of the examined complete specimens.
TABLE 4. Echinoid species recorded at Capo Milazzo.
TABLE 5. Strictly bathyal echinoids inhabiting muddy bottoms of the present Eastern Atlantic (Kohler, 1927; Mortensen, 1928).
Plio-Pleistocene Mediterranean bathyal echinoids: evidence of adaptation to psychrospheric conditions and affinities with Atlantic assemblages
Plain Language Abstract
Although fossil echinoids are generally well represented in the late Cenozoic of the Mediterranean area, well-preserved specimens of species limited to deep-sea bottoms are exceedingly rare. Consequently the evolution of the Miocene to Recent Mediterranean deep-water echinoids is still scantily known. Here we studied the Plio-Pleistocene deep-water echinoid faunas from the Argille Azzurre Formation of north Italy, commonly represented only by isolated spines and plates, and from Capo Milazzo, in north-eastern Sicily. The latter site gave us a rare opportunity of studying much better-preserved and articulate specimens, providing with further evidence of the nature of the deep-Mediterranean fauna that was established after the late Miocene (late Messinian) salinity crisis. The late Piacenzian-Calabrian assemblages studied mainly consist of few regular echinoid species, which can be regarded as typical of cold deep waters in psychrospheric conditions. This (palae)oceanographical setting is different from the current Mediterranean one, which displays a homothermic regime, a strongly stratified (oligotrophic) deep water body, where the echinoid assemblage is represented by few eurythermal species, with a biodiversity influenced by the eastern Atlantic. Our results also indicate that the studied bathyal regular-echinoid assemblages show strong affinities with the western Atlantic, the Caribbean area in particular. This suggests that during the late Cenozoic, at least until the Calabrian, the deep-water Mediterranean-Atlantic faunal exchange within echinoids was settled by biogeographic dynamics different from that occurring today.
Riassunto in italiano
Echinoidi batiali del Plio-Pleistocene mediterraneo: adattamento a condizioni psicrosferiche e affinità con le associazioni atlantiche.
Nonostante gli echinoidi fossili siano generalmente frequenti nei depositi tardo cenozoici dell’area mediterranea, gli esemplari ben conservati appartenenti a specie esclusive di acque profonde sono così rari che l’evoluzione di questo gruppo, in ambienti profondi di tale bacino, è scarsamente conosciuta per ciò che attiene all’intervallo Miocene-Recente. In questo lavoro vengono esaminati gli echinoidi plio-pleistocenici della Formazione delle Argille Azzurre (nord Italia), che sono normalmente rappresentati solo da piastre e spine disarticolate, e quelli di Capo Milazzo (Sicilia nord orientale), località che ha fornito materiale più completo e meglio preservato. Questi fossili ci hanno consentito di approfondire temi paleoambientali e paleogeografici relativi alla fauna profonda degli echinoidi del Mediterraneo nel periodo seguente la crisi di salinità del tardo Miocene (Messiniano superiore). Le associazioni studiate, riferibili al tardo Piacenziano-Calabriano, mostrano una stretta affinità con quelle oggi viventi nelle fredde acque del piano batiale dell’Atlantico occidentale, l’area caraibica in particolare, in condizioni psicrosferiche. Tale quadro paleoambientale è assai diverso da quello presente nell’attuale Mediterraneo profondo, dove l’assetto oceanografico è caratterizzato da un regime di omotermia, in cui le associazioni profonde di echinoidi sono rappresentate da specie euriterme con spiccata affinità est-atlantica. Lo studio mostra quindi che durante il tardo Cenozoico, almeno sino al Calabriano, lo scambio faunistico tra Mediterraneo e Atlantico, con riguardo agli echinoidi, era regolato da dinamiche biogeografiche differenti da quelle attuali.
Parole chiave: Echinoidi batiali; Psicrosfera; Atlantico; Mediterraneo; Plio-Pleistocene; Sicilia.
Traduttori: gli autori
Resumen en Español
Los equinoideos batiales del Plio-Pleistoceno en el área Mediterránea: evidencia de adaptación a condiciones psicrosféricas y afinidad con las asociaciones del Atlántico.
Aunque los equinoideos fósiles generalmente están bien representados en el Cenozoico tardío del área Mediterránea, los ejemplares bien preservados de especies exclusivas de fondos de aguas profundas son extremadamente raros. Como consecuencia, la evolución de los equinoideos de aguas profundas en el Mediterráneo todavía es poco conocida para el intervalo Mioceno-Actual. En el presente trabajo se estudian las faunas plio-pleistocénicas de equinoideos de aguas profundas procedentes de la Formación Argille Azzure, del norte de Italia (normalmente presentándose como radiolas y placas sueltas), y de Capo Milazzo, en el noreste de Sicilia. Esta última localidad nos concedió una oportunidad única de estudiar ejemplares articulados mucho mejor preservados. Estos fósiles aportan nuevas evidencias sobre la naturaleza de la fauna marina profunda del Mediterráneo que se estableció con posterioridad a la crisis de salinidad del Mioceno superior (Messiniense superior). Las asociaciones de equinoideos de edad Piacenziense-Calabriense estudiadas en este trabajo consisten fundamentalmente en algunas especies que se consideran típicas de aguas frías y profundas propias de condiciones psicrosféricas. Este contexto (paleo)oceanográfico es diferente del que impera actualmente en el Mediterráneo, caracterizado por un régimen homotérmico y por un cuerpo de aguas profundas oligotróficas fuertemente estratificado, en el que las asociaciones de equinoideos están representadas por algunas especies euritérmicas, y cuya biodiversidad denota influencia del Atlántico oriental. Nuestros resultados indican asimismo que las asociaciones de equinoideos regulares batiales estudiadas muestran una fuerte afinidad con aquellas propias del Atlántico occidental, en particular de la zona Caribeña. Estos indicios sugieren que durante el Cenozoico tardío, al menos hasta el Calabriense, la interrelación migratoria de las faunas de equinoideos de aguas profundas del Atlántico y del Mediterráneo se estableció debido a una dinámica biogeográfica diferente a la actual.
Palabras clave: Equinoideos batiales; Psicrosféra; Atlántico; Mediterráneo; plio-pleistoceno; Sicilia.
Traductor: Diego A. García-Ramos
Résumé en Français
Échinides bathyales du Plio-Pléistocène de la Méditerranée: adaptation aux conditions psychrosphériques et affinités avec les assemblages de l'Atlantique.
Bien que les échinides fossiles soient bien représentés dans le Cénozoïque supérieur de la région méditerranéenne, les espèces limitées aux fonds profonds sont extrêmement rares. En fait, l'évolution de échinides méditerranéens profonds dans le Miocène-Actuel est encore très peu connue.
Nous avons étudié les échinides profonds du Pliocène-Pléistocène de la Formation Argille Azzurre (nord de l'Italie), souvent représentés par des épines et des plaques isolées, et de Capo Milazzo (Sicile nord orientale). Le dernier site nous a fourni une rare occasion d'étudier des spécimens beaucoup mieux préservés et articulés. Ces fossiles fournissent une évidence supplémentaire de la nature de la faune profonde méditerranéenne qui se plaça après la crise de salinité du Miocène supérieur (Messinien supérieur). Les associations du Plaisancien-Calabrien étudiées sont principalement constituées de quelques espèces d'échinides réguliers, qui peuvent être considérées comme typiques des eaux froides profondes dans des conditions psychrosphériques. Ce cadre (palé)océanographique est différent de celui de la Méditerranée actuelle, qui montre un régime d'homothermie et relative oligotrophie, où l'association des échinides est représentée par peu d'espèces eurythermes, avec une biodiversité influencée par l'Atlantique oriental. Nos résultats indiquent également que les échinides régulièrs méditerranéens étudiés montrent de fortes affinités avec les associations de l'Atlantique occidental, les Caraïbes en particulier. Ça suggère que pendant le Cénozoïque supérieur, au moins jusqu'à le Calabrien, l'échange des associations des échinides profonds entre la Méditerranée et l'Atlantique était réglé par des dynamiques biogéographiques différentes.
Mots cles: Échinides bathyales; Psychrosphére; Atlantique; Méditerranée; Plio-Pléistocène; Sicile.
Traducteurs: Maria Luisa Runfola et Vittorio Garilli
Deutsche Zusammenfassung
Bathyale Echinoide aus dem mediterranen Plio-Pleistozän: Nachweis auf Adaption auf psychrospherische Bedingungen und Ähnlichkeiten mit atlantischen Assemblagen.
Paläontologische Nachweise von autochthonen Tiefwasser-Echinoiden sind so selten, dass die hier beschriebene gut erhaltene Assemblage aus dem Plio-Pleistozän von Capo Milazzo (Nordost-Sizilien) eine wichtige Möglichkeit darstellt, die Biodiversität der bathyalen Echinoiden des spätkänozoischen Mittelmeeres zu untersuchen. Die hier untersuchte diversitätenarme Fauna wird von Cidaris margaritifera, Histocidaris sicula und Stirechinus scillae dominiert, Arten, die eng verwandt sind mit den rezenten Echinoiden, die heute auf die tiefen Abhänge des westlichen Atlantiks beschränkt sind. Die Echinoiden Assemblagen von Capo Milazzo und der Plio-Pleistozänen Argille Azzurre Formation (Italien) teilen eine Anzahl von Arten von denen die meisten auch aus den seichten Bereichen der Plio-Pleistozänen Ablagerungen und dem heutigen Mittelmeer bekannt sind. C. margaritifera ist die einzige strikt bathyale Echinoide, die in beiden Formationen vorkommt. Die paläoökologische Untersuchung dieser Echinoiden weist auf eine epibenthische Lebensweise in trüben Tiefenwässern mit psychrospherischen Konditionen hin. Die folgenden Arten aus der Argille Azzurre Formation werden als strikt bathyal interpretiert: Histocidaris rosaria, Schizaster braidensis und Schizaster ovatus (transferiert in die Gattung Holanthus). Die moderne mediterrane (verarmte) Tiefwasser-Echinoiden Assemblage zeigt Ähnlichkeiten mit derjenigen aus dem nord-östlichen Atlantik und, mit Ausnahme von Holanthus expergitus, sind alle mediterranen Arten aus bathyalen Tiefen eurybathisch, da sie ebenso in Küstenregionen leben. Im Gegensatz dazu zeigen die bathyalen Echinoiden von Capo Milazzo starke Ähnlichkeit mit strikten Tiefwasser-Arten aus dem westlichen Atlantik, besonders mit denen aus der Karibik. Sie verschwanden während des Quartärs wegen des Verlusts der psychrospherischen Bedingungen aus dem Mittelmeer. Basierend auf der Punta Mazza Sektion und datiert mit Nanofossilien und aus der Literatur ist ihre stratigrafische Reichweite in Capo Milazzo spätes Piacenzium-Calabrium.
Keywords: bathyale Echinoide; psychrospherisch; Atlantik; Mittelmeer; Plio-Pleistozän; Sizilien
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024