

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
 Yasuyuki Nakamura
Yasuyuki Nakamura
Tropical Biosphere Research Center
University of the Ryukyus
Senbaru 1, Nishihara
Okinawa 903-0213
Japan
ynaka.riukiaria@gmail.com
Yasuyuki Nakamura is a postdoctoral researcher of the Tropical Biosphere Research Center, University of the Ryukyus. He received his degree (PhD) from University of the Ryukyus in historical biology. His current research focuses on the biogeography of amphibians and squamate reptiles of the Ryukyus and other Japanese islands with special reference to human impacts on those animals as inferred from Late Quaternary fossils and archeological remains.

 Hidetoshi Ota
Hidetoshi Ota
Institute of Natural and Environmental Sciences
University of Hyogo, and the Museum of Nature and Human Activities
Yayoi-gaoka 6, Sanda
Hyogo 669-1546
Japan
ohta@hitohaku.jp
Hidetoshi Ota is a professor of the Institute of Natural and Environmental Sciences, University of Hyogo, and a curator of the Museum of Nature and Human Activities. He previously served as a faculty member of the University of the Ryukyus for 20 years, during which he studied various aspects of vertebrate fauna of the Ryukyu Archipelago. His current interests include systematics, biogeography, and natural history of reptiles and amphibians in the East Asian and western Pacific islands. He received his PhD from Kyoto University.
FIGURE 1. Maps of the Ryukyu Archipelago (1, 2) and Okinawajima Island (3). The map of Okinawajima shows topography, distribution of the Pleistocene limestone, and study sites. Geological data were obtained from Kizaki (1985).
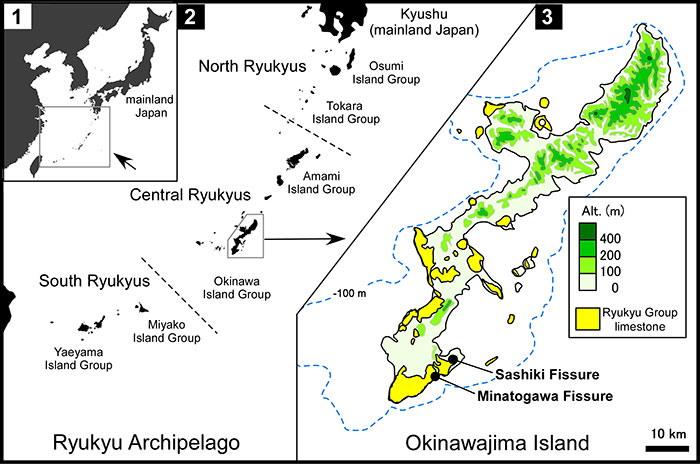
FIGURE 2. Photograph of the Sashiki Fissure (left) and schematic figure showing the structure of the fissure (right). In the right figure, broken lines represent the outline of the fissure behind rock, shaded areas represent studied sediments, and open circles represent the locations of the dating samples.
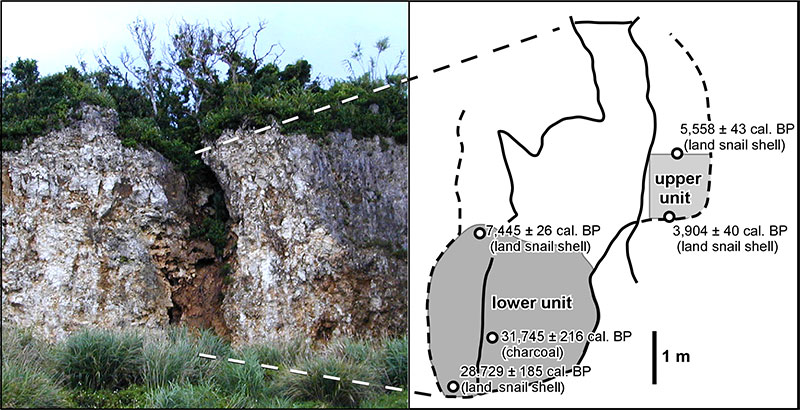
FIGURE 3. Fossils referred to Limnonectes namiyei (1-4) and Babina holsti (5-11). 1-3, left female humerus lacking the proximal and distal parts and the crista ventralis (YMHF-MA 001) in ventral (1), medial (2) and dorsal (3) views; 4, left ilium lacking the anterior part (RUMF-GF-04000) in lateral view; 5-7, right female humerus (one of six registered as RUMF-GF-04003) in ventral (5), medial (6) and dorsal (7) views; 8-10, right male humerus lacking the proximal part (one of two registered as RUMF-GF-04004) in ventral (8), medial (9), and dorsal (10) views; and 11, right ilium lacking the anterior part (one of nine registered as RUMF-GF-04005) in lateral view. Abbreviations: acet, acetabulum; acet.m, acetabular margin; cr.dors, crista dorsalis; cr.lat, crista lateralis; cr.med, crista medialis; cr.par, crista paraventralis; cr.ven, crista ventralis; e.cap, eminentia capitata; ep.rad, epicondylus radialis; ep.ul, epicondylus ulnaris; fo.div, fossula dividens; il.sh, ilial shaft; ol.sc, olecranon scar; p.asc, pars ascendens; stm, spina tuberculi medialis; supr.fo, supracetabular fossa; tub.sup, tuber superior. Scale bars equal 5 mm.
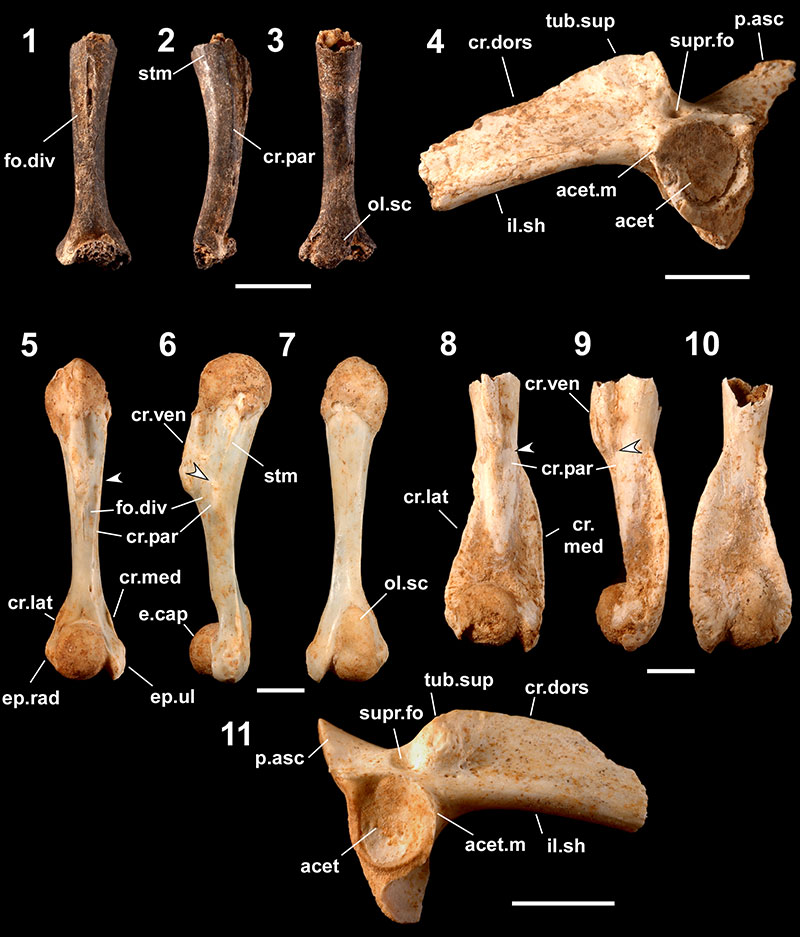
FIGURE 4. Fossils referred to Odorrana ishikawae (1-7) and Odorrana narina (8-14), 1-3, right female humerus (one of 10 registered as RUMF-GF-04009) in ventral (1), medial (2), and dorsal (3) views; 4-6, right male humerus (one of three registered as RUMF-GF-04010) in ventral (4), medial (5), and dorsal (6) views; 7, right ilium lacking the anterior part with part of the ischium (one of five registered as RUMF-GF-04011) in lateral view; 8-10, right female humerus (one of eight registered as RUMF-GF-04014) in ventral (8), medial (9), and dorsal (10) views; 11-13, right male humerus lacking the proximal part of the shaft and the distal part of the epicondylus ulnaris (RUMF-GF-04015) in ventral (11), medial (12), and dorsal (13) views; and 14, pelvic girdle (fused right and left ilia [lacking anterior parts] with the ischium and the pubis: RUMF-GF-04016) in right lateral view. Abbreviations are the same as Figure 3. Arrows indicate the proximal ends of the crista paraventralis. Scale bars equal 5 mm.
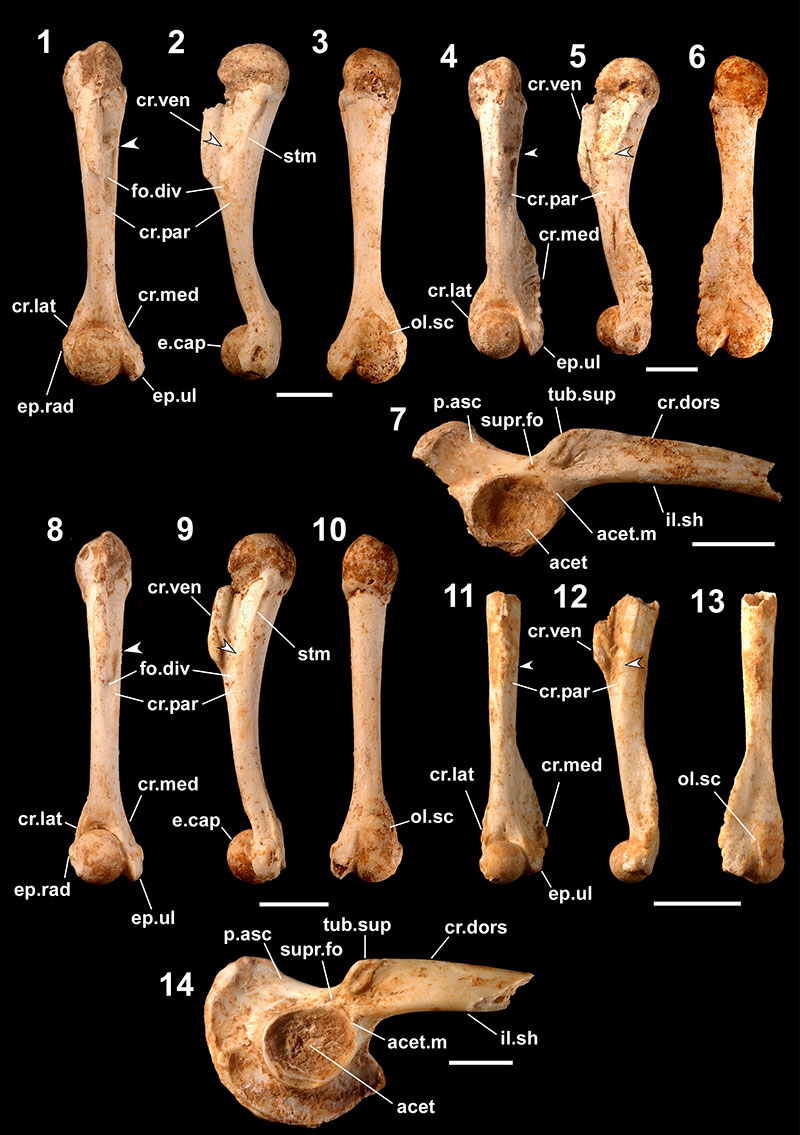
FIGURE 5. Fossils referred to Rana ulma (1-7) and Microhyla okinavensis (8). 1-3, right female humerus (one of 176 registered as RUMF-GF-04019) in ventral (1), medial (2), and dorsal (3) views; 4-6, right male humerus (one of 87 registered as RUMF-GF-04020) in ventral (4), medial (5), and dorsal (6) views; 7, right ilium (one of 93 registered as RUMF-GF-04021) in lateral view; and 8, right ilium (one of two registered as RUMF-GF-04023) in lateral view. Abbreviations are the same as Figure 3. Arrows indicate the proximal ends of the crista paraventralis. Scale bars equal 1 mm.
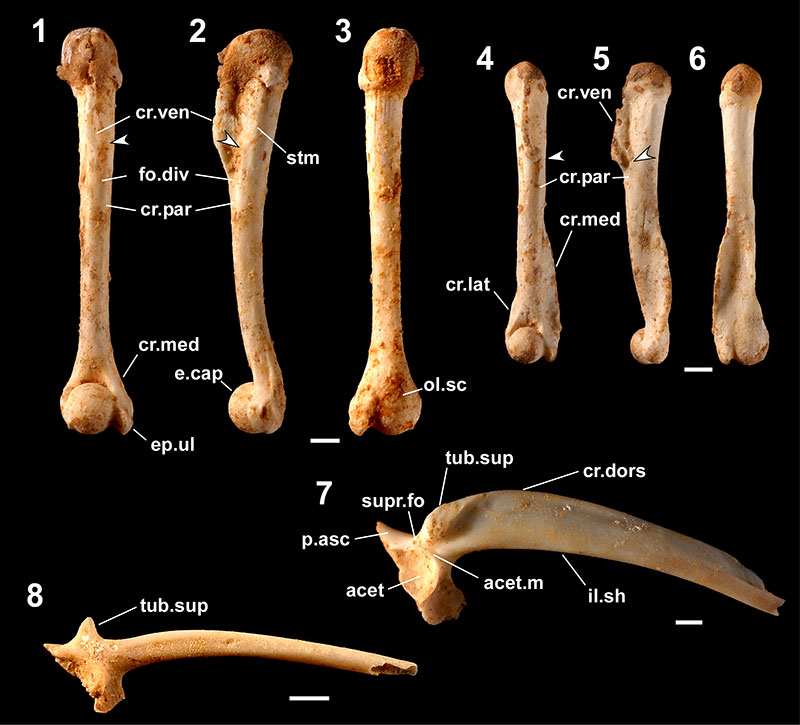
FIGURE 6. Fossils referred to Buergeria japonica (1-4) and Rhacophorus viridis viridis (5-11). 1-3, right female humerus lacking the proximal part (YMHF-MA 010) in ventral (1), medial (2), and dorsal (3) views; 4, left ilium lacking most part of the crista dorsalis (RUMF-GF-04024) in lateral view; 5-7, left female humerus (one of seven registered as RUMF-GF-04025) in ventral (5), medial (6), and dorsal (7) views; 8-10, left male humerus (RUMF-GF-04027) in ventral (8), medial (9), and dorsal (10) views; and 11, pelvic girdle (fused right and left ilia [lacking anterior parts] with the ischium: RUMF-GF-04029) in left lateral view. Abbreviations: pre.acet, preacetabular zone; the others are the same as Figure 3. Arrows indicate the proximal ends of the crista paraventralis. Scale bars equal 1 mm.
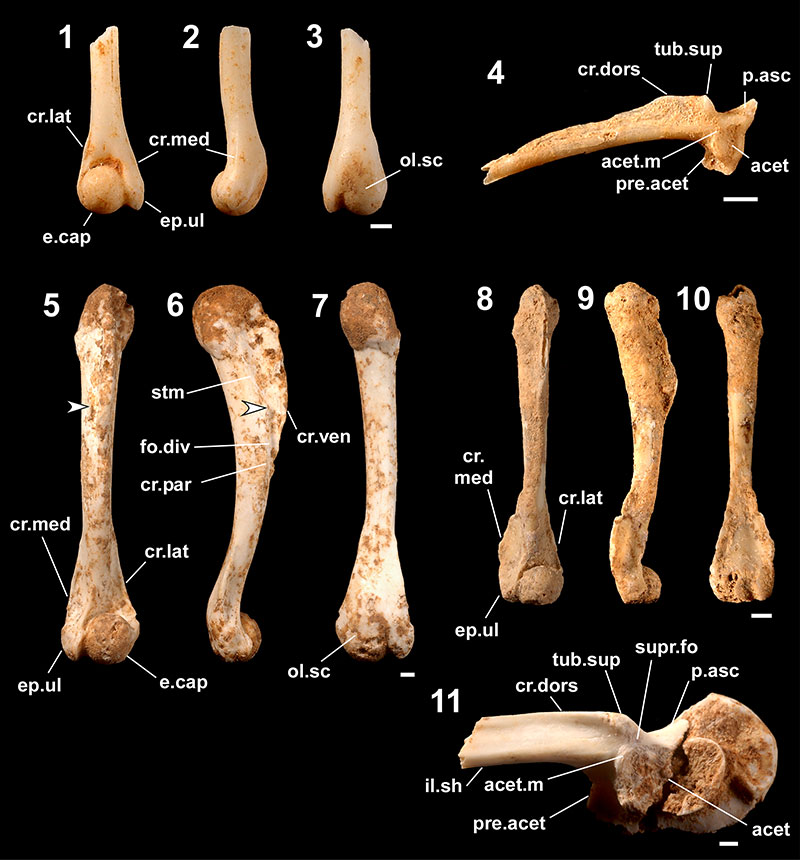
FIGURE 7. Fossils referred to Cynops ensicauda. 1-4, postatlantal precaudal vertebra (one of 108 registered as RUMF-GF-04039 ) in anterior (1), left lateral (2), dorsal (3), and ventral (4) views; 5 and 6, atlas (one of five registered as RUMF-GF-04038) in anterior (5) and left lateral (6) views; 7, parietal-prootic-exoccipital (RUMF-GF-04033) in dorsal view; 8, right maxilla (one of three registered as RUMF-GF-04032) in lateral view; 9, right dentary (one of nine registered as RUMF-GF-04037) in medial view; 10, right rib (one of 25 registered as RUMF-GF-04040) in posterior view; 11, right humerus (one of 85 registered as RUMF-GF-04041) in lateral view; and 12, right femur (one of 92 registered as RUMF-GF-04042) in posterior view. Abbreviations: con, condyle; diap, diapophyses; epi.pr, epipleural processes; neu.sp, neural spine; n.prep, notch for prearticular; parap, parapophyses; pos.pr, posterior process; subd.d, subdental ditch; zygap, zygapophyses. The arrow in 12 indicates the concavity (see text). Scale bars equal 1 mm.

FIGURE 8. Fossils referred to Echinotriton andersoni. 1-4, postatlantal precaudal vertebra (one of 290 registered as RUMF-GF-04052) in anterior (1), left lateral (2), dorsal (3), and ventral (4) views; 5 and 6, atlas (one of 11 registered as RUMF-GF-04051) in anterior (5) and left lateral (6) views; 7, parietal-prootic-exoccipital (RUMF-GF-04047) in dorsal view; 8, right maxilla (one of 30 registered as RUMF-GF-04045) in lateral view; 9, right quadrate (one of five registered as RUMF-GF-04049) in dorsal view; 10, right dentary (one of 70 registered as RUMF-GF-04050) in medial view; 11, right rib (one of 181 registered as RUMF-GF-04053) in posterior view; 12, right humerus (one of 144 registered as RUMF-GF-04054) in lateral view; and 13, right femur (one of 163 registered as RUMF-GF-04055) in posterior view. Abbreviations: tro, trochanter; the others are the same as Figure 7. Scale bars equal 1 mm.

FIGURE 9. Maps showing current distributions (shaded areas) of five forest frogs (Limnonectes namiyei a, b, d, f-h, k, n [ 1], Babina holsti a, b, d, f-k, m [ 2 ], Odorrana ishikawae a, b, d, f-h, j-l, o [ 3 ], O. narina a, b, f-h [ 4 ], and Rana ulma a, b, d, f-k, o [5]) and the newt Echinotriton andersoni a-j (6) on Okinawajima Island and adjacent islets. Superscripts after the species names indicate the data sources—a and b: Chigira (1977, 2003); c: Honda et al. (2012); d: Koba (1957); e: Matayoshi et al. (1977); f-h: Okinawa General Bureau, North Dam Construction Office (1997, 1998, 2002); i and j: Sato (1989, 1993); k: Tanaka (1993); l: Toyama (1988); m: Tominaga et al. (2014); n: Utsunomiya (1996b); and o: Y.N. personal observations at Oku. “?” denotes a population lacking a record or observation in the past 30 years.
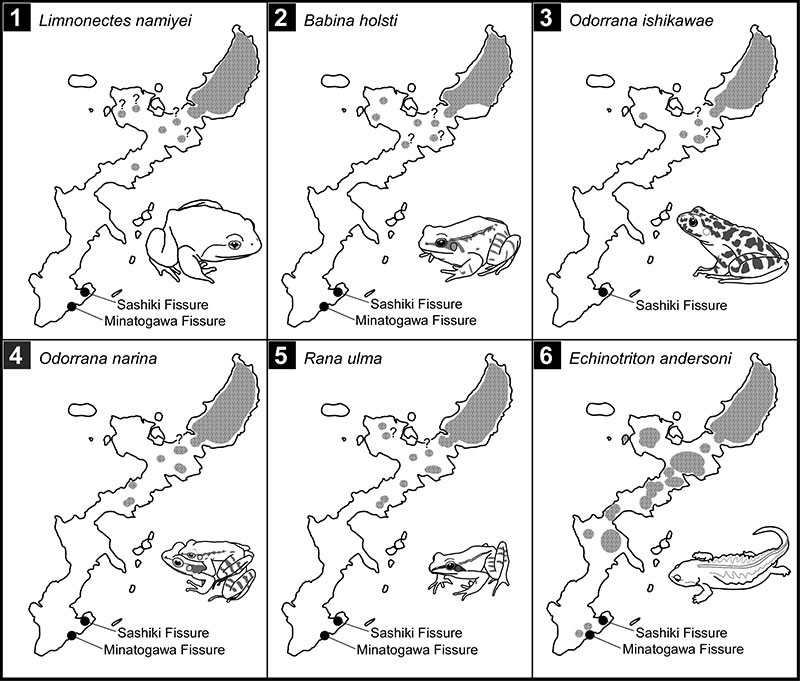
TABLE. Amphibian taxa and abundance (in minimum number of individuals [MNI]) from two Late Pleistocene-Holocene fissures in Okinawajima, and a comparison with the results of Hasegawa (1980). - indicates absence of the fossil, * denotes extralimital taxa (mainland Japanese species: Glandirana rugosa and Rana tagoi ; Southeast Asian species: Polypedates leucomystax), and ** denotes a taxon mentioned by Nokariya (1983b).
List of comparative skeletal specimens examined (including partially skeletonized specimens). Institutional and collection abbreviations are as follows: collection number of University Museum of the Kagoshima University, Kagoshima, Japan (OT, IK, HT); Kyoto University Museum Zoological collection, Kyoto University, Kyoto, Japan (KUZ); Yasuyuki Nakamura private collection (NPN); and National Science Museum Tokyo collection, Tokyo, Japan (NSMT).
Bombina orientalis: NPN 178, 179, Gangwong-do, South Korea (one male and one female).
Bufo bankorensis: NPN 105, Yilan Co., Taiwan; NPN 106-110, Wulai, Taipei Co., Taiwan (two males and four females).
Bufo gargarizans miyakonis: NPN 097, 098, four un-catalogued specimens, Miyakojima Is., Okinawa Pref., Japan (four males and two females).
Bufo japonicus formosus: NPN 065, 066, 068, 069, Kanagawa Pref., Japan (three males and one female).
Bufo japonicus japonicus: NPN 082-084, 089, 090, Kagoshima Pref., Japan (three males and two females).
Bufo torrenticola: NPN 070-072, 076, 077, Nara Pref., Japan (three males and two females).
Duttaphrynus melanostictus: NPN 100, 101, Keelung, Taiwan; NPN 099, Taoyuan Co., Taiwan; NPN 102, Ranong, Thailand (two males and two females).
Hyla japonica: KUZ R56877, Nagano Prf., Japan; NPN 664, 665, Oita Prf., Japan; NPN 1109, Shizuoka Prf., Japan (one male and three females).
Hyla hallowellii: NPN 308, 309, 311, 312, Okinawajima Is., Okinawa Prf., Japan (two males and two females).
Fejervarya kawamurai: NPN 030-034, 1127, 1128, 1134, 1135, Okinawajima Is., Okinawa Prf., Japan; KUZ R56879, R56880, Wakayama Prf., Japan (four males and seven females).
Fejervarya sakishimensis: NPN 114, 116, 117, Ishigakijima Is., Okinawa Prf., Japan (one male and two females).
Limnonectes fujianensis: NPN 325, 327, Taipei Co., Taiwan (one male and one female).
Limnonectes namiyei: KUZ R65738, R71534, R72180, R72422, R72423, Okinawajima Is., Okinawa Prf., Japan (two males and three females).
Babina holsti: KUZ R62704, R62978, R72031, NSMT 00855, Okinawajima Is., Okinawa Prf., Japan (three males and one female).
Babina subaspera: OT-880815-01, OT-940723-03, OT-940723-04, OT-940725-06, OT-940726-07, KUZ R28159, Amamioshima Is., Kagoshima Prf., Japan (two males and four females).
Glandirana emeljanovi: NPN 192, 193, Gangwong-do, South Korea (two females).
Glandirana rugosa: KUZ R56881, R 56882, R56884, Wakayama Prf., Japan; NPN 046, 047, 722-725, Oita Prf., Japan (four males and five females).
Hylarana guentheri: KUZ R38877, Matsu Is., Taiwan (one female).
Hylarana latouchii: NPN 160, 164, Taipei Co., Taiwan (one male and one female).
Lithobates catesbeianus: NPN 1012, Okinawajima Is., Okinawa Prf., Japan; NPN 1122, Iheyajima Is., Okinawa Prf., Japan (two males).
Nidirana adenopleura: NPN 155, 156, Taipei Co., Taiwan (one male and one female).
Nidirana (or Babina) okinavana : NPN 125, 126, 128, 131, Ishigakijima Is., Okinawa Prf., Japan (two males and two females).
Odorrana amamiensis: NPN 132, 133, 135, 136, 294, Amamioshima Is., Kagoshima Prf., Japan (three males and two females).
Odorrana ishikawae: KUZ R57893, R62749, R65737, Okinawajima Is., Okinawa Prf., Japan (two males and one female).
Odorrana splendida: KUZ R28147, R28155, R28156, R28159, Amamioshima Is., Kagoshima Prf., Japan (two males and two females).
Odorrana narina: NPN 003, 014-018, 269, 271, 276-278, 284, 372-376, Okinawajima Is., Okinawa Prf., Japan (nine males and eight females).
Odorrana supranarina: NPN 355-358, 360, 361, Ishigakijima Is., Okinawa Prf., Japan (three males and three females).
Odorrana utsunomiyaorum: KUZ R62289, Ishigakijima Is., Okinawa Prf., Japan; KUZ R62458, R62459, R62461, Iriomotejima Is., Okinawa Prf., Japan (two males and two females).
Pelophylax nigromaculatus: KUZ R56872, R56874, NPN 662, 663, Nagano Prf., Japan; NPN 667-670, Oita Prf., Japan; NPN 813, 814, Nara Prf., Japan (three males and seven females).
Pelophylax porosus porosus: NPN 211, 214, 216, 217, Nagano Prf., Japan (two males and two females).
Pelophylax porosus brevipodus: NPN 049-055, Nagano Prf., Japan; NPN 1114, 1116, Shiga Prf., Japan (three males and five females).
Rana dybowskii: NPN 245-247, 249, Tsushima Is., Nagasaki Prf., Japan (two males and two femals).
Rana huanrensis: NPN 170, Gangwong-do, South Korea (one male).
Rana japonica: NPN 330-332, 044, 045, 661, 666, Oita Prf., Japan (three males and four females).
Rana kobai: NPN 827, 828, Kakeromajima Is., Kagoshima Prf., Japan; NPN 1106-1108, 1111, Amamioshima Is., Kagoshima Prf., Japan (four males and two females).
Rana longicrus: KUHE 50594, Taipei Co., Taiwan (one female).
Rana ornativentris: KUZ R56888, R56889, Nagano Prf., Japan; NPN 321, 322, 324, Kanagawa Prf., Japan (two males and three females).
Rana pirica: NPN 316, 317, Hokkaido Prf., Japan (one male and one female).
Rana sakuraii: NPN 336-339, Kanagawa Prf., Japan (two males and two females).
Rana sauteri: KUHE 50596, Tainan, Taiwan (one female).
Rana tagoi tagoi: KUZ R56878, Hyogo Prf., Japan; NPN 021, 022, 048, 334, 335, 342-344, Kanagawa Prf., Japan; NPN 1113, Nagano Prf., Japan; NPN 1123, 1124, Nara Prf., Japan (five males and seven females).
Rana tsushimensis: NPN 297-302, Tsushima Is., Nagasaki Prf., Japan (three males and three females).
Rana ulma: NPN 001, 002, 006, 008, 010-012, 023, 112, 119, 121, 122, 243, Okinawajima Is., Okinawa Prf., Japan (six males and seven females).
Microhyla okinavensis: NPN 035-038, 1136-1139, Okinawajima Is., Okinawa Prf., Japan (three males and five females).
Buergeria buergeri: KUZ R56887, Wakayama Prf., Japan; NPN 345, 346, 353, Oita Prf., Japan (two males and two females).
Buergeria japonica: NPN 024, 026-029, 1131, 1132, Okinawajima Is., Okinawa Prf., Japan (three males and four females).
Buergeria robusta: NPN 146, 550, Taipei Co., Taiwan (one male and one female).
Polypedates leucomystax: NPN 019, 020, 042, 043, 1125, 1126, 1129, 1130, 1133, Okinawajima Is., Okinawa Prf., Japan (three males and six females).
Rhacophorus arboreus: NPN 204, 205, Kanagawa Prf., Japan (two males).
Rhacophorus schlegelii: NPN 079, Nara Prf., Japan; NPN 196, 203, Shizuoka Prf., Japan (one male and two females).
Rhacophorus owstoni: NPN 113, 320, 387, Iriomotejima Is., Okinawa Prf., Japan (two males and one female).
Rhacophorus viridis amamiensis: NPN 136, 137, 139, Amamioshima Is., Kagoshima Prf., Japan (two males and one female).
Rhacophorus viridis viridis: NPN 004, 005, 039, 040, 222, 225-227, 367, 371, Okinawajima Is., Okinawa Prf., Japan (five males and five females).
Kurixalus eiffingeri: NPN 319, Ishigakijima Is., Okinawa Prf., Japan; NPN 318, Iriomotejima Is., Okinawa Prf., Japan (one male and one female).
Hynobius nigrescens: NPN 1140, Locality and gender unknown.
Hynobius lichenatus: NPN 1141, Locality and gender unknown.
Cynops ensicauda: NPN 025, 314, 315, Okinawajima Is., Okinawa Prf., Japan (one male and two females).
Cynops pyrrhogaster: NPN 303, 304, Kagoshima Prf., Japan (one male and one female).
Echinotriton andersoni: KUZ R72424, R72425, Okinawajima Is., Okinawa Prf., Japan (two females).
Late Pleistocene-Holocene amphibians from Okinawajima Island in the Ryukyu Archipelago, Japan: Reconfirmed faunal endemicity and the Holocene range collapse of forest-dwelling species
Plain Language Abstract
In this paper, we report amphibian fossils excavated from two Late Pleistocene-Holocene sites on Okinawajima Island of the Ryukyu Archipelago, southwestern Japan. The identified species (eight frogs and two newts) were mostly those currently confined to this island or to this and adjacent islands of the Central Ryukyus. Two mainland Japanese frog species and one Southeast Asian frog species, previously reported from one of the study sites (Minatogawa Fissure) are considered misidentifications. This study also revealed occurrences of five forest-dwelling frogs in the southern part of Okinawajima during the Late Pleistocene that are currently strictly confined to the northern forested area of the island, suggesting that the former area was also largely covered by humid forests with abundant lotic habitats in the Late Pleistocene. Such an environment seems to have been lost in the southern part of the island by the middle Holocene, because amphibian species identified from the middle Holocene deposits there did not include those five species at all.
Abstract in Japanese

Translation by author
Resumen en Español
Los anfibios del Pleistoceno tardío-Holoceno de la isla de Okinawajima en el archipiélago de Ryukyu, Japón: endemicidad faunística reconfirmada y disminución del rango de hábitats de las especies de bosque durante el Holoceno
Los fósiles de anfibios obtenidos de dos fisuras del Pleistoceno tardío-Holoceno en la parte sur de la isla de Okinawajima, del archipiélago de Ryukyu, fueron identificados por la comparación detallada con especímenes óseos de taxones existentes. Las especies identificadas (ocho ranas y dos tritones) se limitaron a ciertas especies actuales de la isla, las cuales eran en su mayoría endémicas de la isla o de la parte central del archipiélago con inclusión de la misma. Como consecuencia, se consideran errores de identificación los registros anteriores del Pleistoceno de una de las localidades de estudio (la Fisura Minatogawa) de especies de ranas de otras áreas de distribución (dos especies continentales japonesas [Glandirana rugosa and Rana tagoi] y una rana del sudeste asiático actualmente naturalizada en el archipiélago Ryukyu [Polypedates leucomystax]). Este también puede ser el caso de Fejervarya kawamurai, una rana sinantrópica actualmente con una distribución amplia en el este de Asia, incluyendo las islas de Ryukyu Central. De las especies obtenidas de los depósitos del Pleistoceno tardío en estas localidades, cinco ranas (Limnonectes namiyei, Babina holsti, Odorrana ishikawae, O. narina, y Rana ulma) son habitantes de los bosques y están actualmente confinadas en la zona boscosa del norte de Okinawajima. Además, ninguna de estas especies fue hallada en los depósitos del Holoceno medio de una de las localidades, la Fisura Sashiki. Nuestros resultados indican que, en el Pleistoceno tardío, el área sur de terreno calizo de Okinawajima albergaba un bosque denso y húmedo con diversos hábitats lóticos, similar al de la zona norte boscosa de la isla en la actualidad, y que estas condiciones ambientales se perdieron alrededor del punto medio del periodo Holoceno.
Palabras clave: Biogeografía; ranas; caliza; tritones; paleoambiente
Traducción: Enrique Peñalver
Résumé en Français
Amphibiens du Pléistocène supérieur-Holocène de l'île Okinawajima dans l'archipel de Ryukyu, au Japon: reconfirmé l'endémicité de la faune et l' effondrement de la gamme d'espèces vivantes dans les forêts de l'Holocène
Des fossiles d'amphibiens découverts dans deux fissures Pléistocène-Holocène dans la partie sud de l'île Okinawajima de l'archipel de Ryukyu ont été identifiés grâce à des comparaisons détaillées avec des spécimens squelettiques de taxons actuels. Les espèces identifiées (huit grenouilles et deux tritons) ont été confinées à des éléments existants sur l'île et étaient pour la plupart endémiques à l'île ou à la partie centrale de l'archipel qui l'inclut. Les registres précédents du Pléistocène supérieur de grenouilles hors-limites (deux espèces japonaises continentales [Glandirana rugosa et Rana tagoi] et une grenouille d'Asie du Sud actuellement naturalisés dans les Ryukyus [Polypedates leucomystax]) provenant de l'un des sites d'étude (la Fissure Minatogawa) sont donc considérés comme des erreurs d'identification. Cela peut également être le cas avec Fejervarya kawamurai, une grenouille synanthropique actuellement distribué largement en Asie de l'Est, y compris les Ryukyus centraux. Parmi les espèces obtenues à partir des dépôts du Pléistocène supérieur à ces sites, cinq grenouilles (Limnonectes namiyei, Babina holsti, Odorrana ishikawae, O. narina, et Rana ulma) sont des espèces vivantes dans les forêts, actuellement confinées dans la zone boisée du nord de Okinawajima. En outre, aucune de ces espèces ont été détectés dans les dépôts Holocène moyen à l'un des sites, la Fissure Sashiki. Nos résultats indiquent que la zone calcaire sud de Okinawajima dans le Pléistocène supérieur abritait une forêt dense et humide avec divers habitats lotiques, similaires à la zone boisée nord de l'île à l'heure actuelle, et que ces conditions environnementales ont été perdus au milieu de l'Holocène.
Mots-clés: Biogéographie; grenouilles; calcaire; tritons; paléoenvironnement
Translator: Kenny J. Travouillon
Deutsche Zusammenfassung
Spätpleistozän-holozäne Amphibien der Okinawajima Insel im Ryukyu Archipelago, Japan: eine Wiederbestätigung der Faunenendemität und der holozäne Kollaps der waldbewohnenden Arten
Es werden fossile Amphibien von zwei spätpleistozänen-holozänen Spalten im südlichen Teil der Okinawajima Insel im Ryukyu Archipelago durch detaillierten Vergleich mit heutigen Taxa identifiziert. Die identifizierten Stücke (acht Frösche und zwei Molche) wurden auf heute vorkommende Elemente auf der Insel beschränkt und waren meist endemisch, entweder auf der Insel oder im zentralen Teil des die Insel umfassenden Archipelagos. Vorherige Nachweise aus dem späten Pleistozän über Frösche außerhalb der Grenze (zwei Arten von den japanischen Hauptinseln [Glandirana rugosa and Rana tagoi] und ein südostasiatischer Frosch der zurzeit in den Ryukyus eingebürgert wird [Polypedates leucomystax]) aus einem der untersuchten Gebiete (die Minatogawa-Spalte) werden daher als Misidentifikationen angesehen. Dies könnte auch mit Fejervarya kawamurai der Fall sein, ein synanthropischer Frosch der momentan in Ostasien weit verbreitet ist, einschließlich der zentralen Ryukyus. Von den Arten aus den spätpleistozänen Ablagerungen dieser Fundstellen sind fünf Frösche (Limnonectes namiyei, Babina holsti, Odorrana ishikawae, O. narina und Rana ulma) waldbewohnend und momentan auf das nordöstliche bewaldete Gebiet von Okinawajima beschränkt. Darüber hinaus wurde keine dieser Arten in den mittelholozänen Ablagerungen in einer dieser Fundstellen, der Sashiki-Spalte gefunden. Unsere Funde zeigen, dass das südliche Kalkgebiet von Okinawajima im späten Pleistozän einen dichten und feuchten Wald mit diversen lotischen Habitaten beherbergte, ähnlich dem nördlichen bewaldeten Gebiet der heutigen Insel und dass diese Umweltbedingungen in der Mitte des Holozäns verloren gingen.
Schlüsselwörter: Biogeographie; Frösche; Kalkstein; Molche; Paläoumwelt
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024