

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
TABLE 1. Fitting surfaces of fossil eggshell fragments to intact extant eggshells using root mean square values.
Within cells, top lines refer to rigid fits, where only rotations and translations were used, while bottom lines correspond to rigid and scaled fits, where both the size of the fossil fragment and rotations/translations were used to arrive at the best ‘fit’. Rigid and scaled fits were performed after arriving at the best ‘fit’ using rigid fits (i.e., rotations and translations only). (a)Values are best-fitting black eagle examples from Table 2: TM 33879, TM42505, TM43471, and TM43505 for fragment T92-88; TM 088 for fragment T93-17. (b)Values are best-fitting guinea fowl examples from Table 3: eggs 2, 4, and 5 for fragment T92-88; egg 5 for fragment T93-17. (c)Fragment includes either the apical or blunt pole so both were assessed. *best match in surface curvature fitting. Number of iterations to achieve an RMS change less than 0.001 are reported in parentheses for each line.
TABLE 2 . Fitting surfaces of fossil eggshell fragments to intact extant Black eagle eggshells using root mean square values.
Within cells, top lines refers to rigid fits, where only rotations and translations were used, while bottom lines correspond to rigid and scaled fits, where both the size of the fossil fragment and rotations/translations were used to arrive at the best ‘fit’. Rigid and scaled fits were performed after arriving at the best ‘fit’ using rigid fits (i.e., rotations and translations only). (a)Egg from an infertile zoo individual. (b)Fragment includes either the apical or blunt pole so both were assessed. *best match in surface curvature fitting amongst non-zoo specimens. Number of iterations to achieve an RMS change less than 0.001 are reported in parentheses.
TABLE 3 . Fitting surfaces of fossil eggshell fragments to intact extant Guinea fowl eggshells using root mean square values.
Within cells, top lines refers to rigid fits, where only rotations and translations were used, while bottom lines correspond to rigid and scaled fits, where both the size of the fossil fragment and rotations/translations were used to arrive at the best ‘fit’. Rigid and scaled fits were performed after arriving at the best ‘fit’ using rigid fits (i.e., rotations and translations only). (a)Fragment includes either the apical or blunt pole so both were assessed. *best match in surface curvature fitting. Number of iterations to achieve an RMS change less than 0.001 are reported in parentheses.
TABLE 4. Carbon and oxygen isotope data for fossil eggshells, eggshell fragments and sediment all collected from the Plio-Pleistocene PCS calcrete exposed at the base of the Dart Pinnacle, Taung.
FIGURE 1. Fossil eggs from Taung, T92-88 on the right and T93-17 on the left.
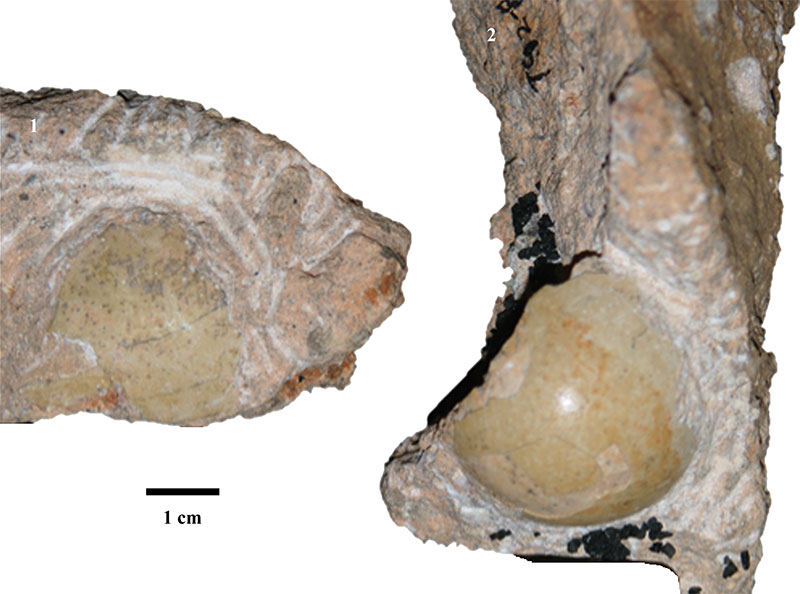
FIGURE 2. All specimens, both extant eggs and fossil eggshells, showing surface morphology at 6.3X magnification.

FIGURE 3. All specimens, both extant eggs and fossil eggshells, showing surface morphology at 12.5X magnification.

FIGURE 4. All specimens, both extant eggs and fossil eggshells, showing surface morphology at 57X magnification.

FIGURE 5. Renderings of fossil eggshell fragments embedded in matrix ( 1 - 2) and indication of the internal boundary between eggshell and matrix ( 3 - 4). Images 1 and 3 are fossil eggshell fragment T92-88, and images 2 and 4 are fossil eggshell fragment T93-17. Note the relatively consistent thickness at the edges and how this maintained surface curvatures indicated by the external surfaces.
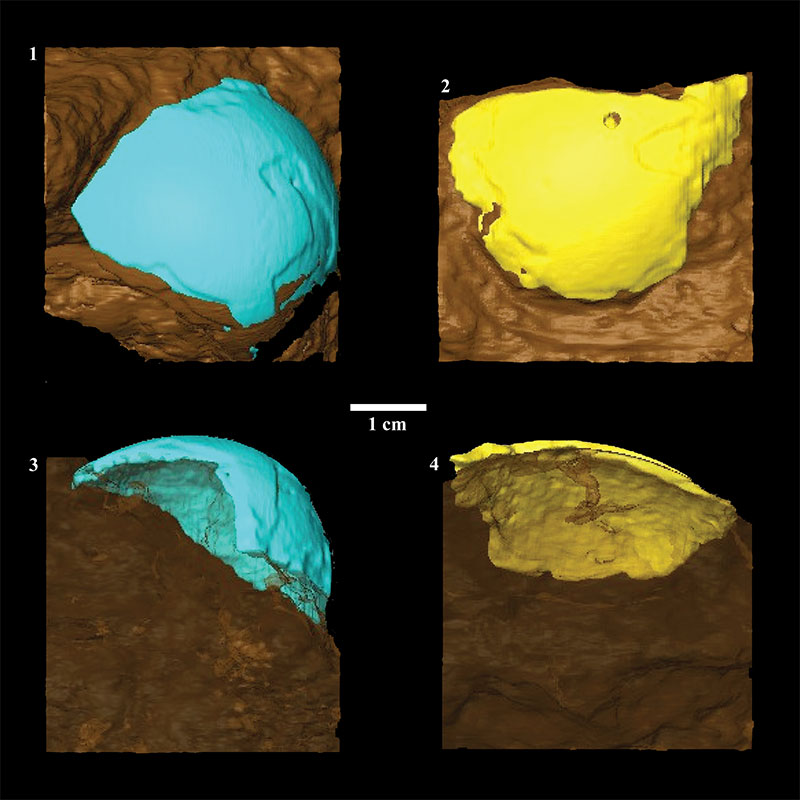
FIGURE 6. Fossil eggshell fragments T92-88 (blue) and T93-17 (yellow) fit to eggs from five extant birds. Fits depicted visually in the figure (rigidly constrained size) correspond to root mean square (RMS) values reported in Table 2. Note that visual correspondence supports quantification of RMS values in that fossil eggshell fragment T92-88 fits best with the blunt pole of the extant guinea fowl egg. Extant black eagle (blunt pole) and giant eagle owl (apical pole) eggs provide the next closest, but still worse fits. Fossil eggshell fragment T93-17 fits best with the equatorial region of the black eagle egg. The extant guinea fowl egg provides the next closest fit for fossil eggshell fragment T93-17.
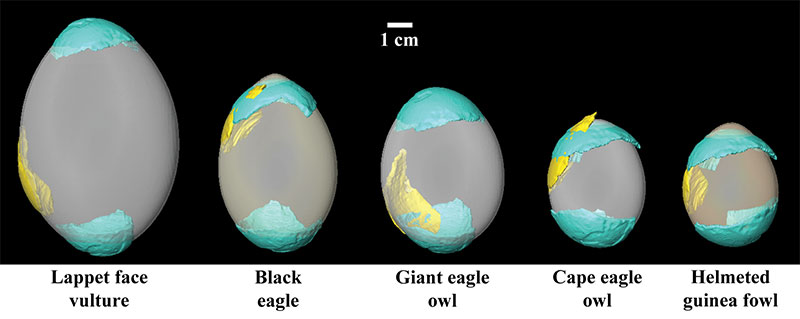
FIGURE 7. Representative SEM results from one (TDES 3) of the fossil eggshells.

FIGURE 8. (1) Fossilized eggshell fragments T92-88 (blue) and T93-17 (yellow) indicating best fit to the same black eagle egg as illustrated in Figure 6, plus six additional black eagle eggs. Note despite intraspecific variability in egg shape, the fits are quite similar (also see Table 3). This suggests even accounting for intraspecific variability in egg shape Results in Table 2 should be robust. (2) Fossil eggshell fragments T92-88 (blue) and T93-17 (yellow) indicating best fit to the same guinea fowl egg as illustrated in Figure 6 (far left), plus four additional guinea fowl eggs. Note that despite intraspecific variability in egg shape, the fits are quite consistent (also see Table 3). This suggests that even accounting for intraspecific variability in egg shape results of the surface curvature analyses are robust.
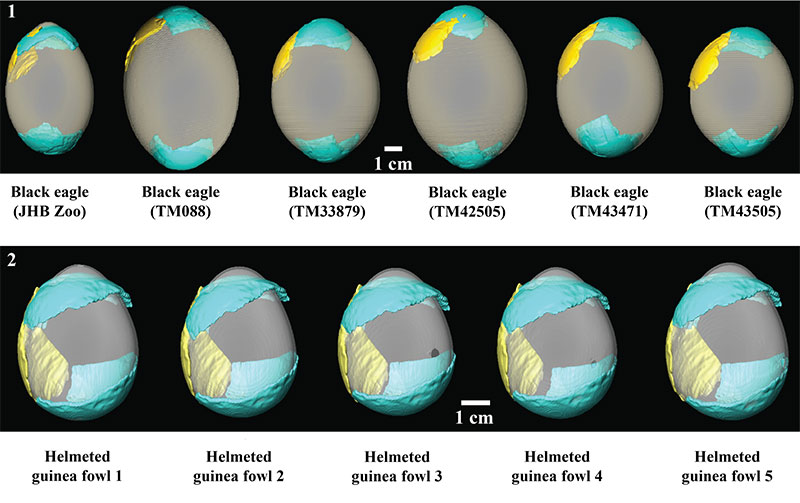
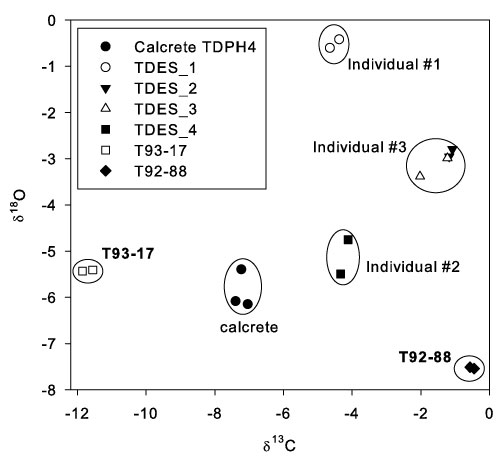
FIGURE 9. Cross-plot of oxygen and carbon isotope values from fossil eggshells and sediment, base of the Dart Pinnacle, Taung. There is a high degree of consistency between replicate samples, and each eggshell fragment shows a distinct isotopic signature, indicating that the eggshells are derived from different individuals (with the possible exception of TDES 2 & TDES 3). The δ13C values indicate that all eggshells have a predominantly C4 (savannah grass) dietary signal, with the exception of T93-17 which has a pure (or almost pure) C3 diet.
 Brian F. Kuhn
Brian F. Kuhn
Evolutionary Studies Institute
University of the Witwatersrand
Johannesburg South Africa
brian.kuhn@wits.ac.za
Dr. Brian Kuhn is a Senior Researcher with the Evolutionary Studies Institute, University of the Witwatersrand, South Africa. His specialties are comparative anatomy, carnivore ecology and fossil carnivores. He is the current permit holder and principle investigator for the Taung World Heritage site.

 Kristian J. Carlson
Kristian J. Carlson
Evolutionary Studies Institute
University of the Witwatersrand
Johannesburg South Africa
and Department of Anthropology
Indiana University
Bloomington, Indiana, USA
kristian.carlson@wits.ac.za
Dr. Kristian Carlson is presently a Senior Researcher with the Evolutionary Studies Institute, University of the Witwatersrand, South Africa. He is a functional morphologist specializing in the use of computer-assisted methods to better understand both extinct and extant species.
 Philip J. Hopley
Philip J. Hopley
Department of Earth and Planetary Sciences
Birkbeck
University of London
London, England
and Department of Earth Sciences
University College London
London, England
p.hopley@ucl.ac.uk
Dr. Phil Hopley is a Lecturer in Palaeoclimatology at Birkbeck, University of London. He is interested in the environmental and climatic context of hominin evolution in Africa. He uses the stable isotope geochemistry of carbonate rocks and fossil tooth enamel to reconstruct Plio-Pleistocene climate change and to investigate the habitat preferences of early hominins.
 Bernhard Zipfel
Bernhard Zipfel
Evolutionary Studies Institute
University of the Witwatersrand
Johannesburg South Africa
bernhard.zipfel@wits.ac.za
Dr. Bernhard Zipfel is the University Curator of Fossils and Rock Collections at the University of the Witwatersrand. He has a special interest in the biomechanics and evolution of the human foot, the origins of hominin bipedalism, palaeopathology and the preservation of natural history collections. He holds a Ph.D. from the University of the Witwatersrand.
 Lee R. Berger
Lee R. Berger
Evolutionary Studies Institute
University of the Witwatersrand
Johannesburg South Africa
lee.berger@wits.ac.za
Professor Lee R. Berger is the Research Professor in Human Evolution and the Public Understanding of Science at the University of the Witwatersrand's Evolutionary Studies Institute. He is also an Explorer in Residence for the National Geographic Society.
Identification of fossilized eggshells from the Taung hominin locality, Taung, Northwest Province, South Africa
Brian F Kuhn, Kristian J. Carlson, Philip J. Hopley, Bernhard Zipfel, and Lee R. Berger
Plain Language Abstract
Fossil eggshells have been associated with the Taung Child since Raymond Dart first saw the accumulation of fossil material brought to him in Johannesburg. For nearly 90 years these eggshells have been mentioned anecdotally, used to both support and refute a theory that the Taung Child was preyed upon by a large eagle, or simply ignored. One thing is for certain, until now no systematic attempt to identify the species responsible for laying the eggs has been completed. Here we use modern technology to identify the species of bird that laid the eggs, as well as discuss the implications of the identifications for the broader picture of Taung as a site.
Resumen en Español
Identificación de cáscaras de huevo fosilizadas de la localidad de homínidos Taung, Taung, Northwest Province, Sudáfrica
Si bien el componente en aves presente en los restos de fauna de Dart Deposits, Taung, Sudáfrica, ha sido discutido durante casi un siglo, los taxones presentes no han sido identificados a nivel de especie. Aquí se lleva a cabo un análisis sistemático de los fragmentos de cáscara de huevo fosilizados con el fin de documentar la presencia de las especies de ave en Taung durante el Plio-Pleistoceno. Un análisis comparativo de la morfología de la superficie y de la curvatura de la superficie de las cáscaras de huevo fragmentarias descartó todos menos tres taxones de aves existentes como fuentes potenciales de los fragmentos fosilizados: un águila de gran tamaño, un búho real (Bubo sp.) o una gallina de Guinea (subfamilia Numidinae). La probabilidad, para cada uno de estos tres taxones, de que sean la fuente de las cáscaras es analizada mediante la evaluación de hasta qué punto la curvatura de la superficie es coincidente entre los fragmentos fosilizados y las cáscaras de huevo actuales. Las dos cáscaras de huevo fósiles más completas recuperadas de Taung tienen distintas firmas de isótopos de carbono lo que indica que pertenecen a dos grupos de ave diferentes, granívoras y carnívoras. Si bien, mediante el establecimiento de una evidencia directa de un componente del grupo de las rapaces en la asociación faunística de Taung, estas identificaciones contribuyen al debate sobre si hubo o no un agente del grupo de las aves en la selección-colección para los fósiles de Taung, incluyendo quizás el Niño de Taung, no permiten establecer un comportamiento depredador-presa específico.
Palabras clave: huevos fósiles; acumuladores de fauna; del grupo de las aves; Taung; Plio-Pleistoceno
Traducción: Enrique Peñalver
Résumé en Français
Identification des coquilles d'œufs fossilisés de la localité d'hominidé Taung, Taung, Province du Nord-Ouest, Afrique du Sud
Alors qu'une composante aviaire au sein des restes fauniques des Dépôts de Dart, à Taung, en Afrique du Sud, a été discuté pendant près d'un siècle, les taxons présents n'ont pas été identifiés au niveau de l'espèce. Ici, nous procédons à une analyse systématique des fragments de coquilles fossilisées afin de documenter la présence de taxons aviaire spécifique à Taung au cours du Plio-Pléistocène. Une analyse comparative de la morphologie de la surface et de la courbure de la surface des coquilles d'œufs fragmentaires a éliminé tous sauf trois taxons aviaires existants comme sources potentielles des fragments fossilisés: un grand aigle, un hibou grand-duc (Bubo sp.) ou une pintade (sous-famille Numidinae). La probabilité pour chacun de ces trois taxons comme source est examinée en évaluant les assortissements de courbure de la surface entre les fragments et les coquilles d'œufs fossilisés et existants. Les deux coquilles fossiles les plus complètes récupérées à Taung ont des signatures isotopiques de carbone distinctes indiquant qu'elles appartiennent à deux guildes différentes, granivores et carnivores. Bien que ces identifications contribuent au débat sur l'opportunité ou non d'avoir un agent de collection aviaire pour les fossiles de Taung, y compris peut-être l'enfant de Taung, en établissant des preuves directes d'un composant rapace dans l'assemblage faunique de Taung, elles ne peuvent pas aborder le comportement de prédateur-proie spécifiquement.
Mots-clés: oeufs fossiles; accumulateurs fauniques; aviaire; Taung; Plio-Pléistocène
Translator: Kenny J. Travouillon
Deutsche Zusammenfassung
Identifizierung fossiler Eischalen aus der Taung Homininen Fundstelle, Taung, Nordost-Provinz, Südafrika
Während beinahe ein Jahrhundert lang über eine Vogelkomponente innerhalb der Faunenüberreste der Dart-Ablagerungen von Taung, Südafrika, diskutiert wurde, wurden die vorliegenden Taxa nicht auf die Art herunter bestimmt. Für eine Dokumentation der Vorkommen spezifischer Vogeltaxa in Taung während des Plio-Pleistozäns führen wir hier eine systematische Analyse der fossilisierten Eierschalenfragmente durch. Eine Vergleichsanalyse der Oberflächenmorphologie und Oberflächenkrümmung der fragmentierten Eierschalen schloss alle außer drei heutige Vogeltaxa als potentielle Quellen für die fossilisierten Fragmente aus: ein großer Adler, eine Adlereule (Bubo sp.) oder ein Perlhuhn (Unterfamilie Numidinae). Die Wahrscheinlichkeit jeder dieser drei Taxa als Quelle wird diskutiert, indem Treffer von Oberflächenkrümmung zwischen den fossilisierten Fragmenten und heutigen Eierschalen evaluiert werden. Die zwei am meisten kompletten Eierschalen aus Taung haben deutliche Kohlenstoffisotopen-Signaturen, was darauf hinweist, dass sie zu zwei unterschiedlichen – granivoren und karnivoren – Guilden gehörten. Auch wenn diese Identifizierungen zur Debatte beitragen, ob (zumindest ein Teil) der Ansammlung von Fossilien in Taung (u.U. inclusive des Taung Kindes) durch Greifvögel erfolgt ist, in dem sie klar anzeigen, dass in der Taung Fauna Greifvögel vorhanden waren, kann trotzdem keine eindeutige Aussage über Räuber-Beute-Verhalten getroffen werden.
Schlüsselwörter: fossile Eier; Pflanzenakkumulatoren; Vogel; Taung; Plio-Pleistozän
Translator: Eva Gebauer
Arabic
in progress
Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024