

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
FIGURE 1. Measurements used for comparisons.

FIGURE 2. Scelidotherium leptocephalum. Skulls in dorsal view. 1, MMP 1155-M; 2, MACN 9625. Scale bar equals 10 mm.
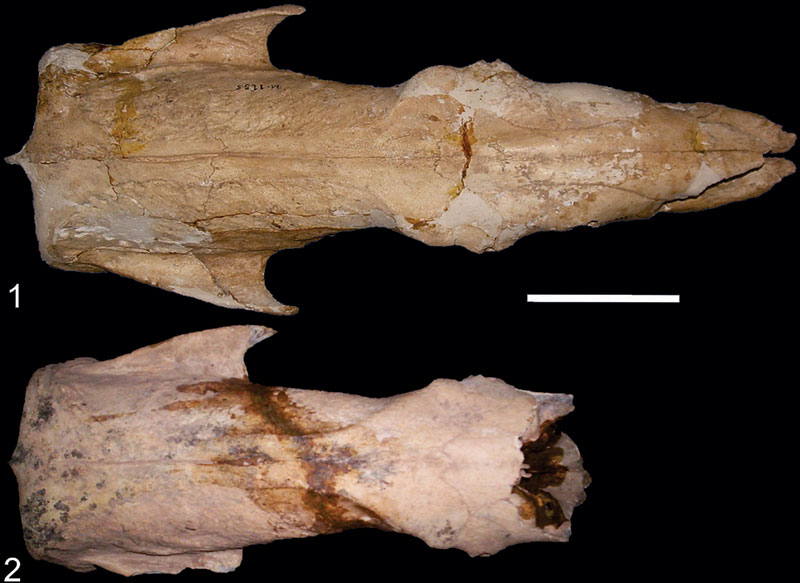
FIGURE 3. Scelidotherium bravardi. Skulls in dorsal view. 1, MMP 157-S; 2, holotype BM(NH)M 37626. Scale bar equals 10 mm.

FIGURE 4. Valgipes bucklandi. Skulls in dorsal view. 1, MCL 4294; 2, MCL 4293. Scale bar equals 10 mm.

FIGURE 5. Catonyx tarijensis. Skull and mandible in dorsal view. 1 and 2 FMNH P 14243. Scale bar equals 10 mm.

FIGURE 6. Catonyx tarijensis. Skull and mandible in dorsal view. 1 and 2 FMNH P 14238. Scale bar equals 10 mm.

FIGURE 7. Catonyx chiliensis. Skulls in dorsal view. 1, BM(NH)M 2819; 2, MHIN-UNSL-GEO V 199. Scale bar equals 10 mm.

FIGURE 8. Catonyx cuvieri. Skulls in dorsal view. 1, MCL 4265; 2, MCL 22683. Scale bar equals 10 mm.

FIGURE 9. Skull outlines in dorsal view. 1 and 2, Scelidotherium leptocephalum (MMP 1155-M, MACN 9625); 3 and 4, Scelidotherium bravardi (MMP 157-S, BM(NH)M 37626); 5 and 6, Valgipes bucklandi (MCL 4294, 4393).
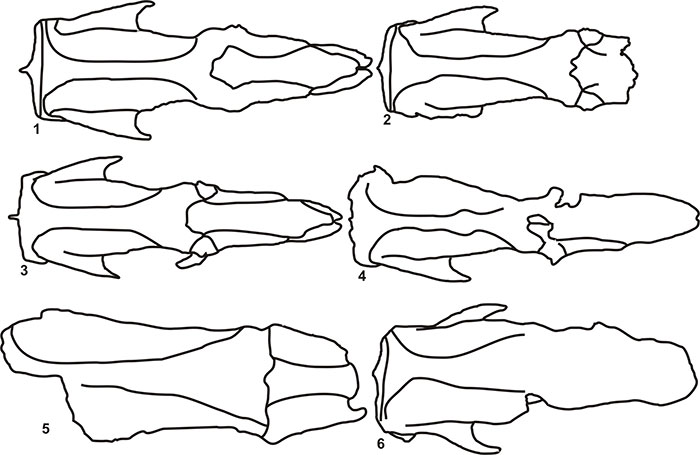
FIGURE 10. Skull and mandible outlines in dorsal view. 1 and 2, Catonyx tarijensis (FMNH P 14243); 3 and 4, Catonyx tarijensis (FMNH P 14238); 5 and 6, Catonyx chiliensis (BM(NH)M 2819 Type, MHIN-UNSL-GEO V 199); 7 and 8, Catonyx cuvieri (MCL 4265, MCL 22683).
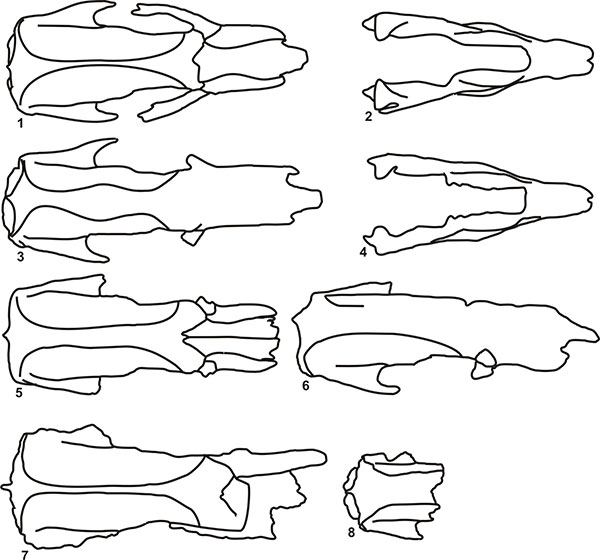
TABLE 1. Comparative measurements (in mm) of skulls. *Approximate.
TABLE 2. Acronyms and list of specimens of Scelidotherium, Valgipes, and Catonyx mentioned in the text.
TABLE 3. Mean of the measurements of the skulls from the two morph types of dorsal crests of Scelidotherium leptocephalum and Catonyx tarijensis. (±) is standard deviation. (*) indicate significant differences in the medians with a Kruscal-Wallis Test (p<0.05).
 Ángel R. Miño-Boilini
Ángel R. Miño-Boilini
Centro de Ecología Aplicada del Litoral (CECOAL-CONICET) y Facultad de Ciencias Exactas y Naturales y Agrimensura
Universidad Nacional del Nordeste
Ruta 5, km 2,5 (CP 3400, CC 128)
Corrientes, Argentin
angelmioboilini@yahoo.com.ar
Ángel R. Miño Boilini was born in Corrientes city, Corrientes, Argentina in 1979. He degree in Biology in 2005 and he gas got the PhD diploma in 2012 at the Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. Currently, he is Professor of Paleobiogeography at the Universidad Nacional del Nordeste and Researcher at the National Research Council of Argentina (CONICET). His main field of research involves the anatomical and systematic study of the Xenarthra Mylodontidae Scelidotheriinae, actively collaborated with numerous paleontologists of Argentina and Bolivia. He has been involved in numerous research projects and published papers, especially in those concerning Xenarthra Pilosa Scelidotheriinae.

 Alfredo E. Zurita
Alfredo E. Zurita
Centro de Ecología Aplicada del Litoral (CECOAL-CONICET) y Facultad de Ciencias Exactas y Naturales y Agrimensura
Universidad Nacional del Nordeste
Ruta 5, km 2,5 (CP 3400, CC 128)
Corrientes, Argentina
azurita@cecoal.com.ar
Alfredo Zurita was born in Presidencia Roque Saénz Peña, Chaco, Argentina in 1974. He degree in Biology in 2000 and he gas got the PhD diploma in 2007 at the Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. Currently, he is Professor of Vertebrate Paleontology at the Universidad Nacional del Nordeste and Researcher at the National Research Council of Argentina (CONICET). His main field of research involves the anatomical and systematic study of the Xenarthra Glyptodontidae, actively collaborated with numerous paleontologists of Argentina, Uruguay, Brazil, Bolivia, Colombia, Venezuela and United States. He has been involved in numerous research projects and published papers, especially in those concerning Xenarthra Glyptodontidae.
Dimorphism in Quaternary Scelidotheriinae (Mammalia, Xenarthra, Phyllophaga)
Plain Language Abstract
Previous contributions have postulated hypotheses about cases of sexual dimorphism (differences in form between males and females of the same species) in fossil ground sloths, more precisely in megatherines and mylodontines from the Pliocene (5-2.6 Ma) and Pleistocene (2.6-0.011 Ma) of South and North America. In this contribution, we present evidence of sexual dimorphism in Quaternary scelidotheres, a particular fossil group of endemic ground sloths from South America that includes three genera and six species. The results show that it is possible to recognize two marked morphological variations within each species at the level of the dorsal crests of the skull and of the anterior part of the mandible. We postulate the hypothesis that specimens with sagittal crest are males whereas specimens lacking sagittal crest are females.
Resumen en Español
Dimorfismo en Scelidotheriinae (Mammalia, Xenarthra, Phyllophaga) del Cuaternario
Son escasas las contribuciones relacionadas a posibles casos de dimorfismo sexual en perezosos actuales y fósiles. Hasta el momento, los estudios sobre dimorfismo sexual en perezosos terrestres se limitan a las subfamilias Megatheriinae (Eremotherium) y Mylodontinae (Paramylodon) del Plioceno y Pleistoceno de América del Sur y América del Norte. Los Scelidotheriinae constituyen un linaje de perezoso terrestre endémico de América del Sur, con un biocrón que abarca el lapso "Friasense"-Lujanense (Mioceno medio-Holoceno temprano). Una revisión filogenética y taxonómica integral de los Scelidotheriinae del Cuaternario sugiere que es posible reconocer tres géneros y seis especies: Scelidotherium Owen (Scelidotherium leptocephalum y S. bravardi), Valgipes Gervais (Valgipes bucklandi), y Catonyx Ameghino (Catonyx cuvieri, C. tarijensis y C. chiliensis). Uno de los aspectos más notables es que en algunos de los materiales estudiados (n = 47) fue la presencia de dos morfologías en cada especie a nivel de las crestas dorsales del cráneo (crestas parasagitales y la cresta sagital) y a nivel de la región más distal de la mandíbula (sólo en C. tarijensis). En todos los taxones estudiados (excepto en S. leptocephalum y S. bravardi), los dos tipos implican la ausencia y presencia de una cresta sagital. Sugerimos que los especímenes con la cresta sagital son machos y los especímenes que carecen de cresta sagital son hembras. Este representa el tercer clado de perezoso terrestre que presenta pruebas de dimorfismo sexual a nivel del cráneo y de la mandíbula.
Palabras clave: perezosos terrestres; Mylodontidae; América del Sur; variabilidad; cráneo; mandíbula
Traducción: Authors and Enrique Peñalver
Résumé en Français
Dimorphisme chez Scelidotheriinae (Mammalia, Xenarthra, Phyllophaga) du Quaternaire
Les contributions concernant des cas possibles de dimorphisme sexuel chez les paresseux fossiles et vivants sont rares. Jusqu'à présent, les études de dimorphisme sexuel chez les paresseux terrestre fossiles ont été limitées aux sous-familles Megatheriinae (Eremotherium) et Mylodontinae (Paramylodon) du Pliocène et du Pléistocène d'Amérique du Sud et du Nord. Scelidotheriinae constitue une lignée endémique de paresseux terrestres d'Amérique du Sud, avec un âge biochrone allant du « Frasien » au Lujanien de SALMAs (Miocène Moyen-Holocène Inferieur). Une révision phylogénétique et taxonomique intégrante de Scelidotheriinae du Quaternaire montre qu'il est possible de reconnaître trois genres et six espèces: Scelidotherium Owen (Scelidotherium leptocephalum et S. bravardi), Valgipes Gervais (Valgipes bucklandi), et Catonyx Ameghino (Catonyx cuvieri, C. tarijensis, et C. chiliensis). L'un des aspects notables dans tous les spécimens analysés (n = 47) était la présence de deux types morphologiques dans chaque espèce au niveau des crêtes dorsales du crâne (crêtes parasagittales et crête sagittale) et au niveau de l'extrémité distale de la mandibule (uniquement chez C. tarijensis). Dans chaque espèce sauf deux (S. leptocephalum et S. bravardi), les deux types impliquent l'absence et la présence d'une crête sagittale. Nous suggérons que les spécimens avec des crêtes sagittales sont des mâles, et les spécimens sans crête sagittale sont des femelles. Ceci représente le troisième clade de paresseux terrestre rapporté avec preuve de dimorphisme sexuel du crâne et de la mandibule.
Mots-clés: paresseux terrestre; Mylodontidae; Amérique Du Sud; variabilité; crâne; mandibule
Translator: Kenny J. Travouillon
Deutsche Zusammenfassung
Dimorphismus bei quartären Scelidotheriinae (Mammalia, Xenarthra, Phyllophaga)
Die Beiträge zu möglichen Fällen von Sexualdimorphismus bei fossilen und lebenden Faultieren sind rar. Bis jetzt waren Studien über den Sexualdimorphismus fossiler Riesenfaultiere auf die Unterfamilien Megatheriinae (Eremotherium) und Mylodontinae (Paramylodon) aus dem Pliozän und Pleistozän von Südamerika und Nordamerika beschränkt. Die Scelidotheriinae beinhalten eine endemische Linie von Riesenfaultieren mit einem biochronen Alter was den Lapsus "Friasian"-Lujanian SALMAs (mittleres Miozän bis frühes Holozän) überspannt. Eine ganzheitliche phylogenetische und taxonomische Revision der quartären Scelidotheriinae zeigt, dass es möglich ist drei Gattungen und sechs Arten zu unterscheiden: Scelidotherium Owen (Scelidotherium leptocephalum und S. bravardi), Valgipes Gervais (Valgipes bucklandi) und Catonyx Ameghino (Catonyx cuvieri, C. tarijensis und C. chiliensis). Einer der bemerkenswertesten Aspekte bei allen analysierten Stücken (n=47) war das Vorhandensein von jeweils zwei Morphotypen bei jeder Art und zwar im Bereich der dorsalen Schädelkämme (Parasagittalkämme und Sagittalkämme) und im Bereich der distalsten Mandibelregion (nur bei C. tarijensis). Bei allen außer zwei Arten (S. leptocephalum und S. bravardi) beinhalten die zwei Typen das Fehlen und das Vorhandensein eines Sagittalkamms. Wir schlagen vor, dass die Stücke mit den Sagittalkämmen Männchen sind und die ohne Sagittalkamm Weibchen. Hiermit wurde über die dritte Klade von Riesenfaultieren mit Nachweis über Sexualdimorphismus in Schädel und Mandibel berichtet.
Schlüsselwörter: Riesenfaultiere; Mylodontidae; Südamerika; Variabilität; Schädel; Mandibel
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024