

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
APPENDIX 1. Codings for Kuroyuriella and Asagaolacerta in Gauthier et al. (2012) and Longrich et al. (2012).
Codings in Gauthier et al. (2012) and Longrich et al. (2012) - A=0&1, B=1&2; C=2&3; D=3&4:
Kuruyuriella (70.4% missing data)
0????????? ??????0?0? ?????????? ?????0?1?0 -00-000420
??0000100? ?20?0?C00? ????01???1 ??????01?2 -?00-?A??0
0111-00??? 00?A00?0?0 0?0000001? 0000?????? ??????????
????????0? ?????????? ????????00 1200?0???? ??????????
?????????? ?????????? ?????????? ?????????? 2?????????
??????1?1- 00?0?0???0 0????????? ??????0?00 ????1?????
?????????? ?????????? ?????????? ?????????? ??????????
????100001 4001002020 00000??0?0 ?0B?101000 0??0-??0?0
0??A0????? ?00-0000D3 3000000000 00?0000??? ??????????
?????????? ?????????? ?????????? ?????????? ??????????
?????????? ?????????- -1??0-0000 0?0??????? ??????????
?????????? 00?0000100 00
Asagaolacerta (85% missing data)
?????????? ?????????? ?????????? ?????0???? ??????????
?????????? ?????????? ?????????? ?B10?????? ??????????
?????????? 00?????1?? ?011?????? ?????????? ?00??10011
000020?00? ?????????? ??????1?00 1B00?02??? ??????????
?????????? ?????????? ?????????? ??0?????0? ???0?1- -??
?????????? ?????????? ?????????? ?????????? ??????????
?????????? ?????????? ?????????? ?????????? ??????????
????1000?1 ????0????? 000000?-?? ?????????? ??????????
?????????? ????0??1?C 2000000000 00?20?0??? ??????????
?????????? ??????11?? ?????????? ?????????? ??????????
?????????? ?????????0 000??????? ?????????? ???????001
?????????? ?????????? ?0???????? ?????????? ??????????
?????????? ?01?0??110 00
APPENDIX 2. Full nexus file of data matrix see zipped file.
FIGURE 1. Map showing the location of the Kaseki-Kabe fossil site, Ishikawa Prefecture.
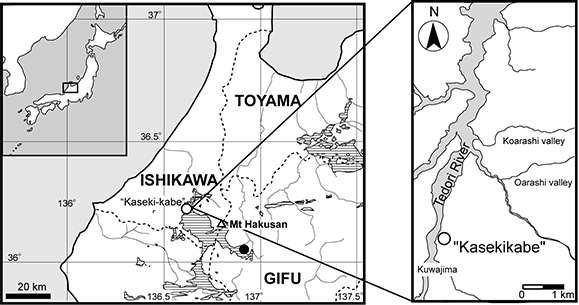
FIGURE 2. Kuroyuriella mikikoi gen. et sp. nov., holotype, SBEI 1510. 1, the four blocks making up the specimen; 2-3, the main association; 4-5, circumorbital bones, pterygoid and premaxilla; 6-7, frontal region in ventral view.
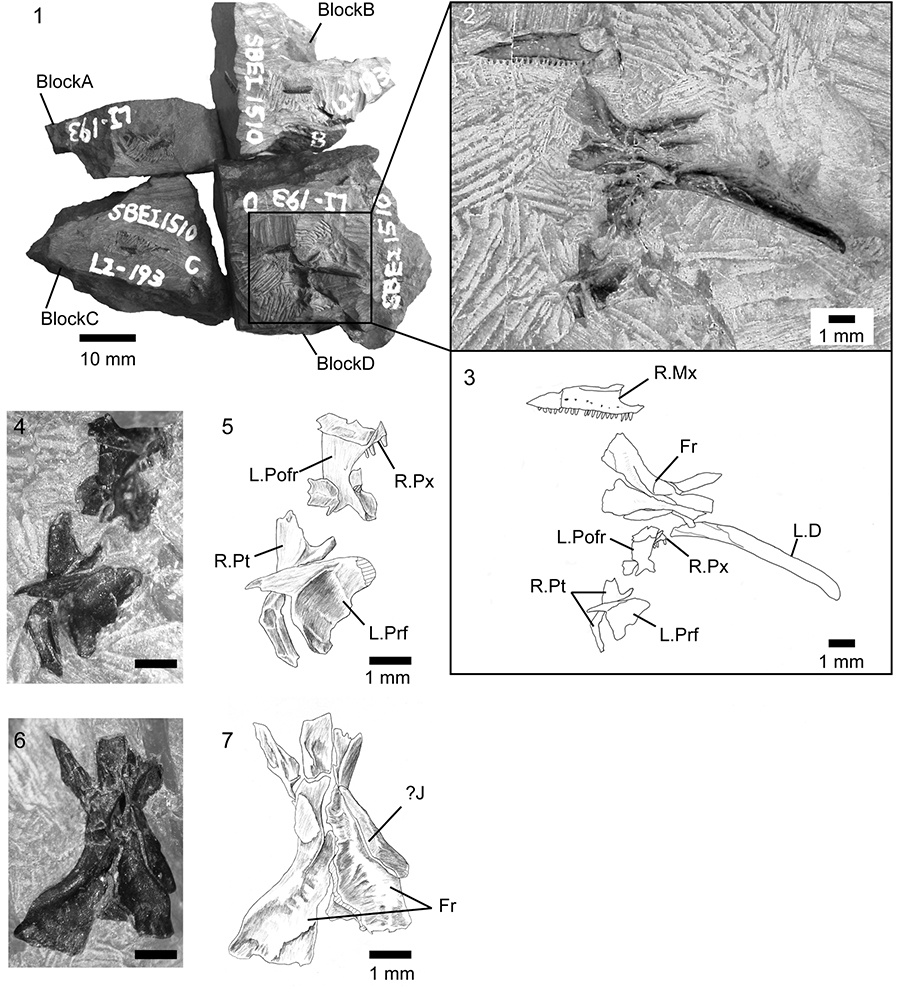
FIGURE 3. Kuroyuriella mikikoi gen. et sp. nov., referred specimen, SBEI 1608. 1-2, main association; 3-4, offset association; 5-6, detail of parietal and frontal; 7-8, detail of quadrate; 9, detail of postdentary bones. For abbreviations, see Material and Methods.
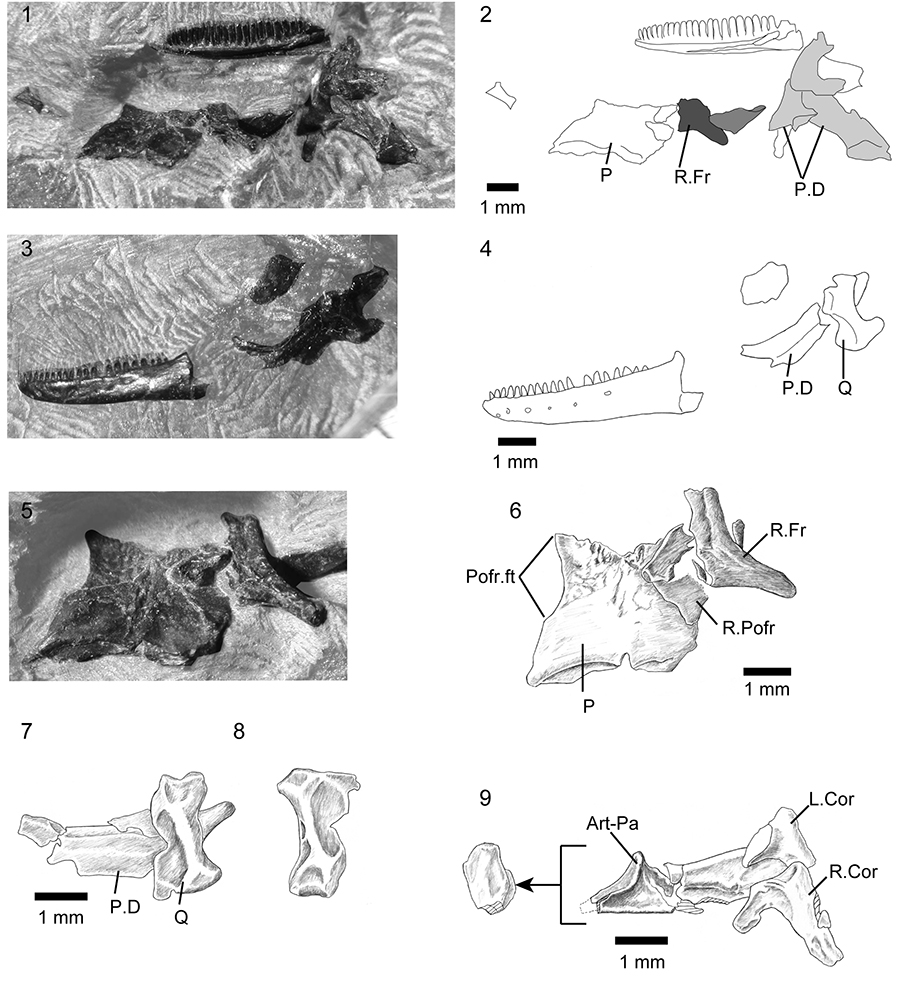
FIGURE 4. Kuroyuriella mikikoi gen. et sp. nov., jaw elements. 1-2, SBEI 1510, right maxilla in labial view; 3-4, SBEI 1608, left dentary in labial view; 5-6, SBEI 1608, right dentary in lingual view; 7-10, SBEI 1510, right mandible in 7-8, labial view, and 9-10, ventrolateral view. For abbreviations, see Material and Methods.
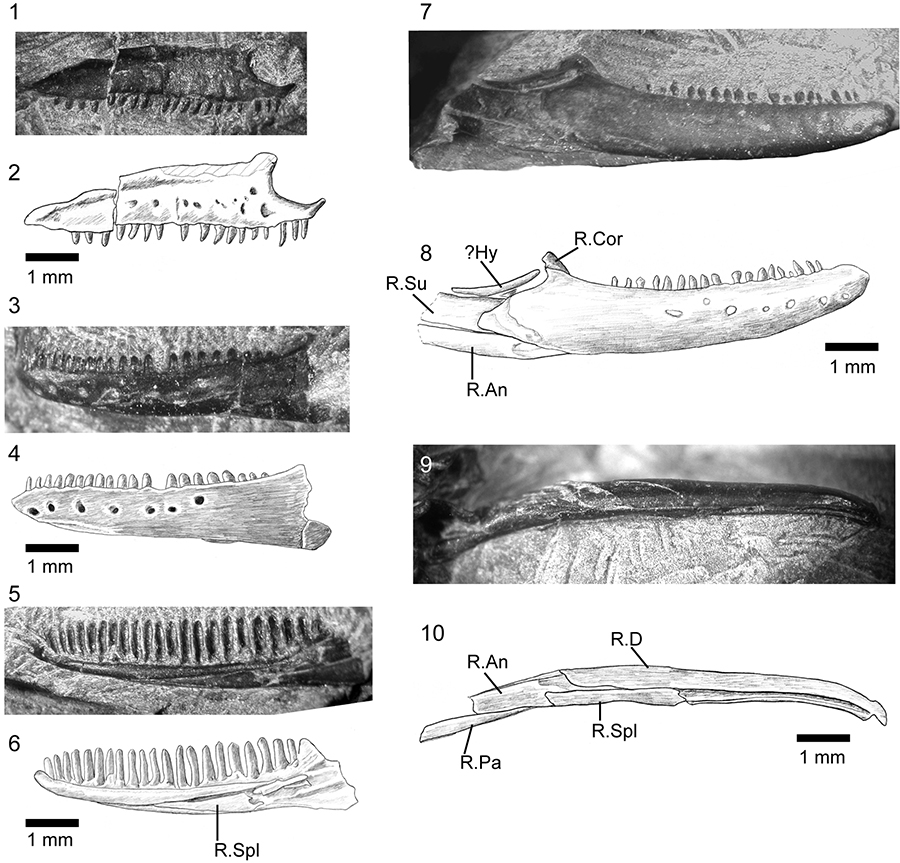
FIGURE 5. Phylogenetic position of Kuroyuriella mikikoi gen. et sp. nov. and Asagaolacerta tricuspidens gen. et sp. nov. in squamate trees, using the morphological data matrix of Gauthier et al. (2012), extended by Longrich et al. (2012). 1, Strict consensus of 1000 trees using unordered equally weighted characters, and run using TNT with 'minisearch.run'. Major clades are condensed. 1Note that Iguania was not monophyletic and comprised four smaller clades whose position was unresolved in relation to Borioteiioidea. Support values at nodes are Bremer/Jacknife/Symmetric resampling; 2, one of three trees (identical at this level) with clades condensed, run using the same matrix as in Figure 5.1 and analysed with TNT (with sectorial search, ratchet [20 iterations], and tree fusion all activated), but with the molecular tree of Wiens et al. (2010) and Pyron et al. (2013) providing the backbone constraint.
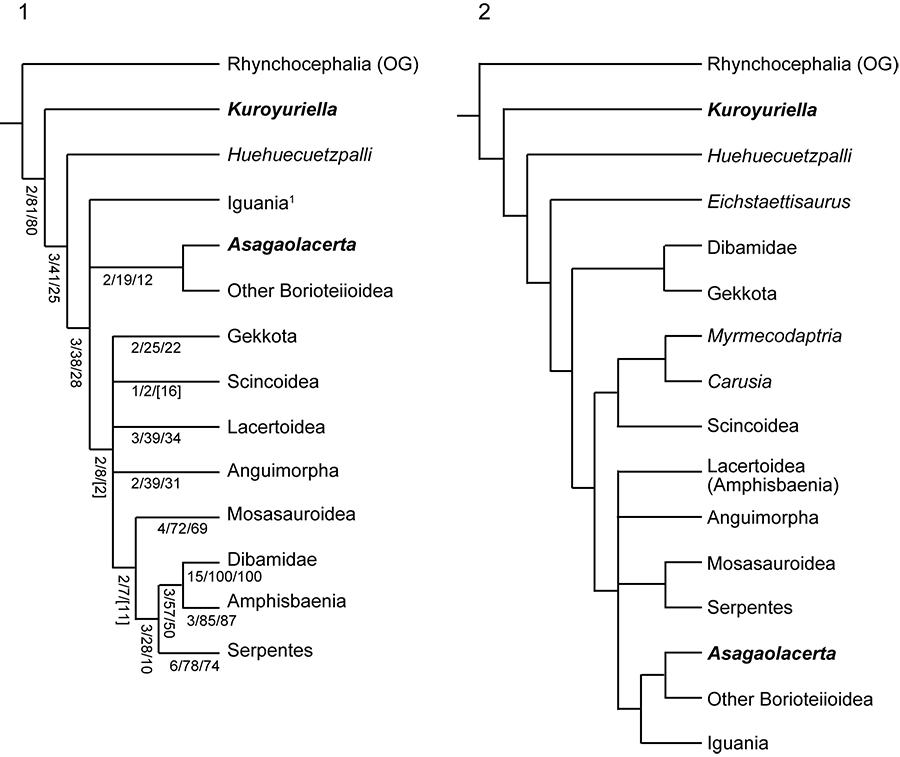
FIGURE 6. The phylogenetic position of Kuroyuriella mikikoi gen. et sp. nov. as recovered by analyses run with Implied Weighting (k=7), and character ordering. 1, no backbone constraint; 2, backbone constraint tree based on the molecular trees of Wiens et al. (2010) and Pyron et al. (2013). Only the scincoid section of each tree is shown as in all analyses with equally weighted taxa, Kuroyuriella lies in the stem-squamate position shown in Figure 5.
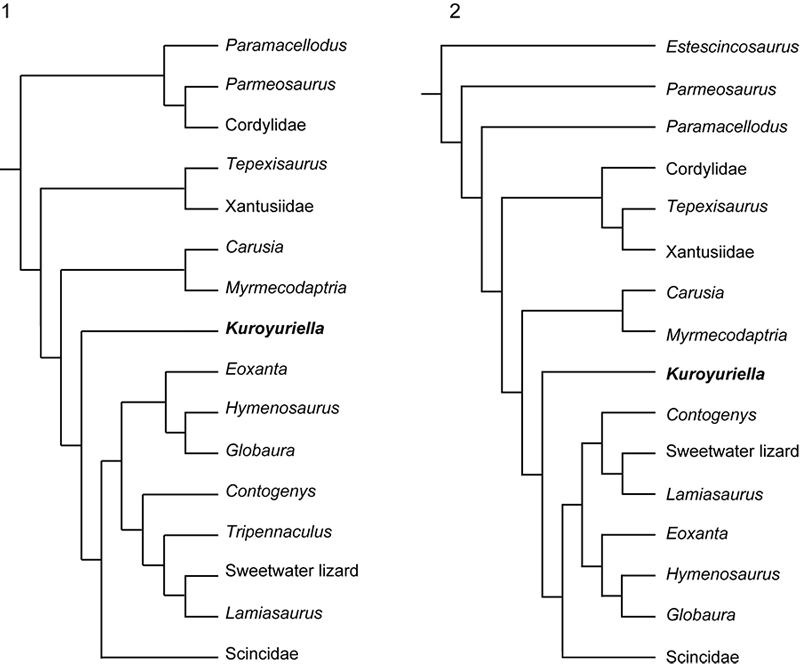
FIGURE 7. Asagaolacerta tricuspidens gen. et sp. nov., holotype, SBEI 1566. 1, the main association with the offset second bone group in the smaller image; 2, explanatory drawing of the same. For abbreviations, see Material and Methods.
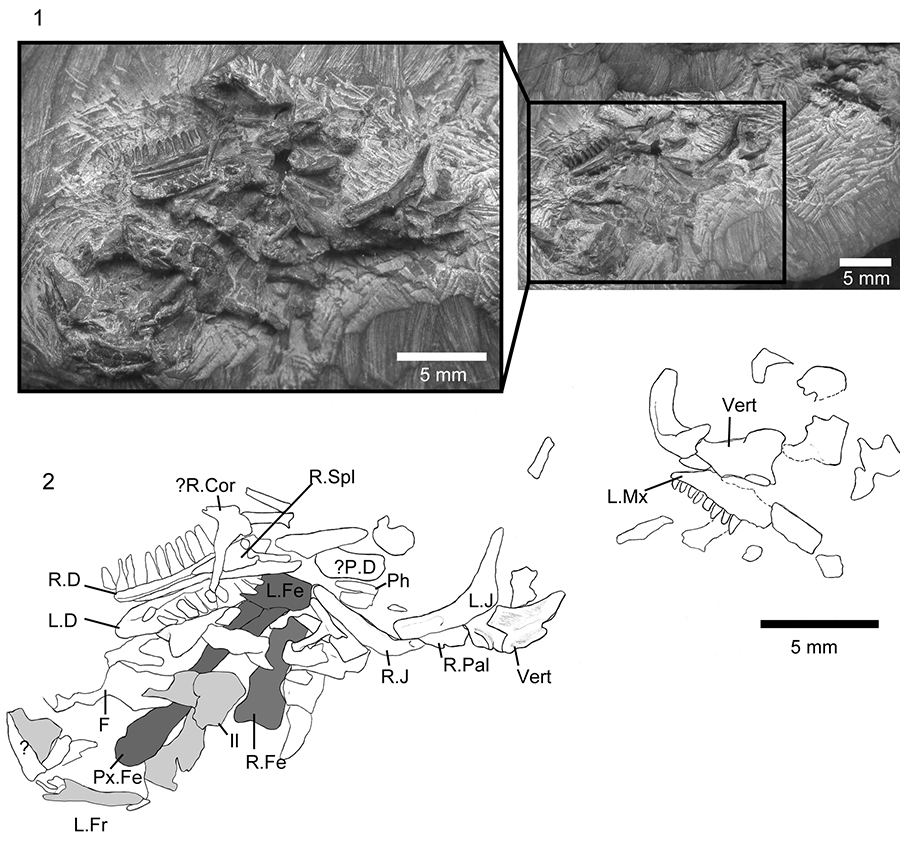
FIGURE 8. Asagaolacerta tricuspidens gen. et sp. nov., holotype, SBEI 1566. 1, Left partial maxilla in labial view; 2, left dentary in lingual view; 3, right dentary in lingual view; 4, details of teeth on the right dentary (enlarged from 3). For abbreviations, see Material and Methods.
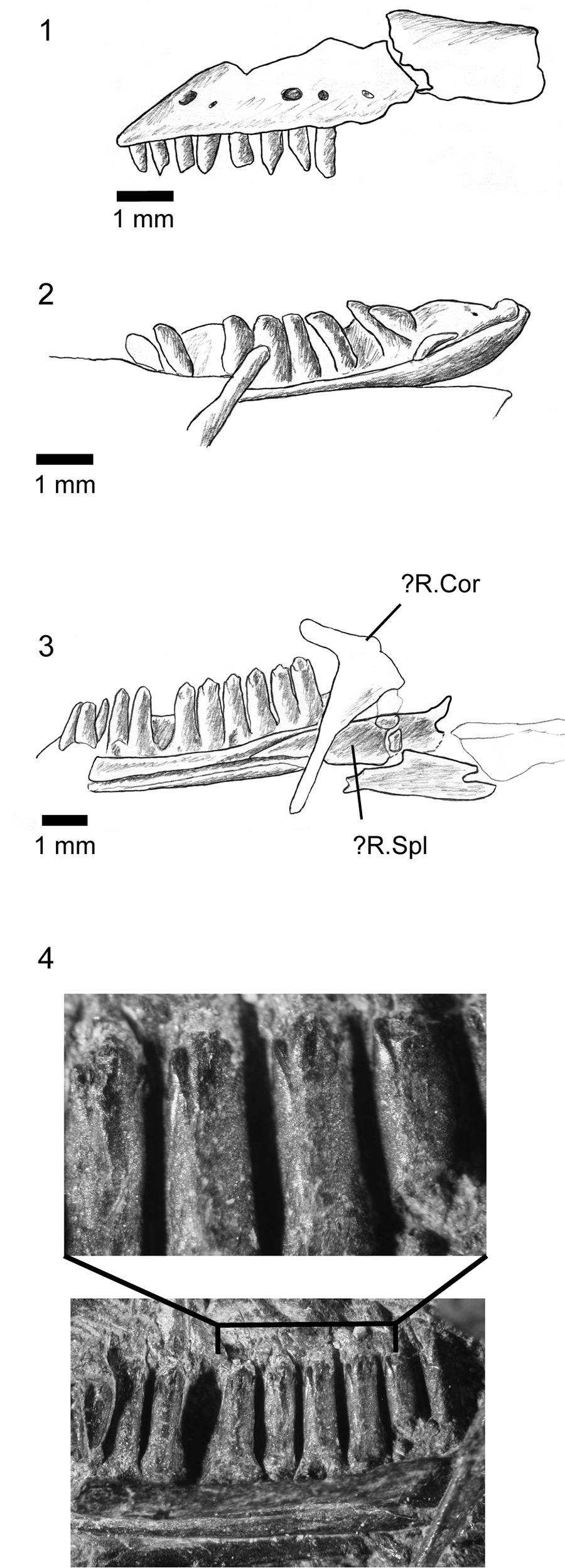
FIGURE 9. Asagaolacerta tricuspidens gen. et sp. nov., holotype, SBEI 1566, details of cranial bones. 1, left and right jugals; 2-3, explanatory drawings of right and left jugal respectively; 4-5, right quadrate in 4, lateral, and 5, anterior views; 6, biradiate bone, possibly coronoid. For abbreviations, see Material and Methods.

FIGURE 10. Asagaolacerta tricuspidens gen. et sp. nov., holotype, SBEI 1566, postcranial elements. 1, presacral vertebrae; 2, left ilium and distal head of right femur. For abbreviations, see Material and Methods.
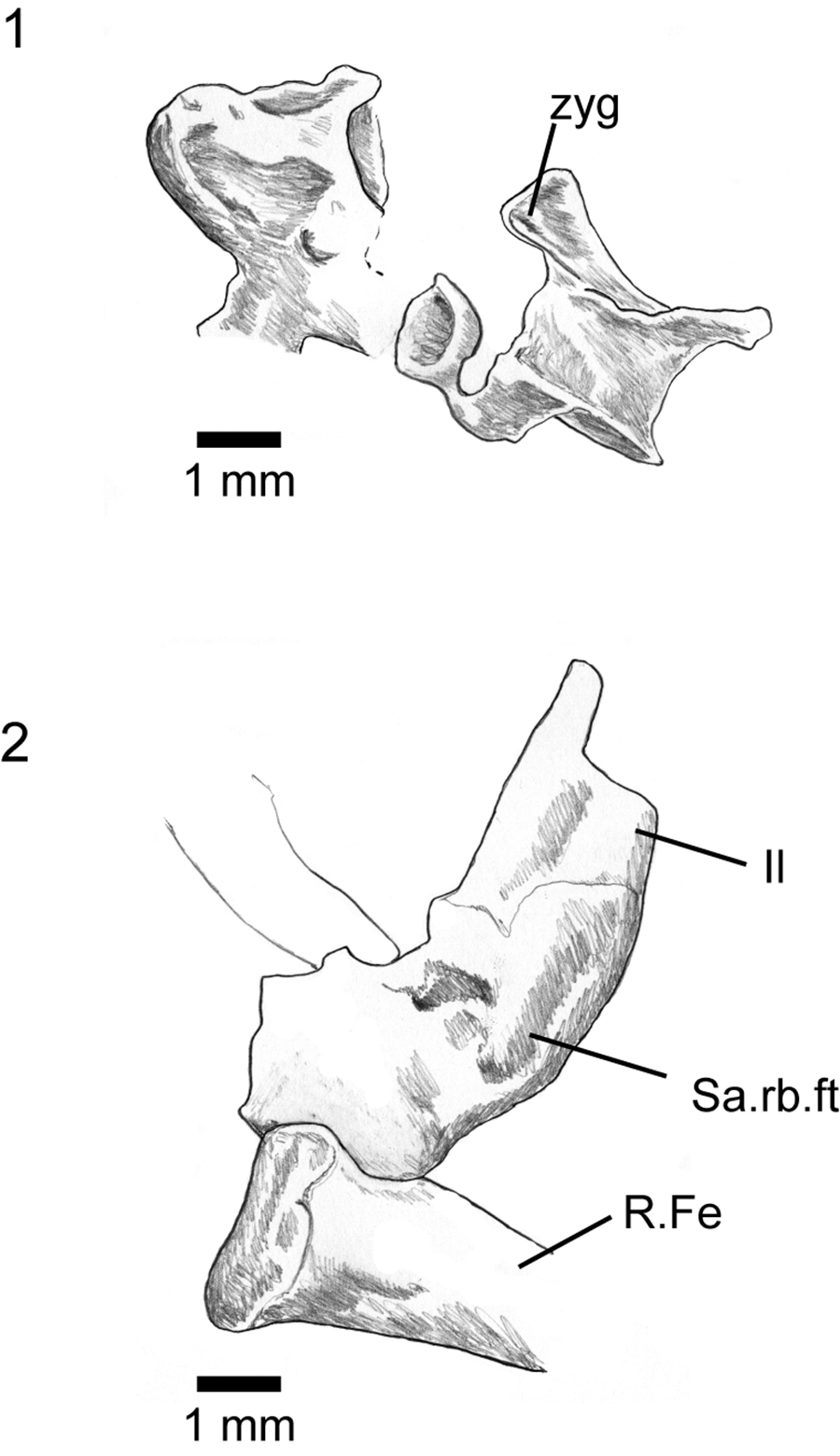
FIGURE 11. Associated postcranial specimens that may be referable to Asagaolacerta gen. nov. 1-2, SBEI 190; 3-4, SBEI 193. For abbreviations, see Material and Methods.
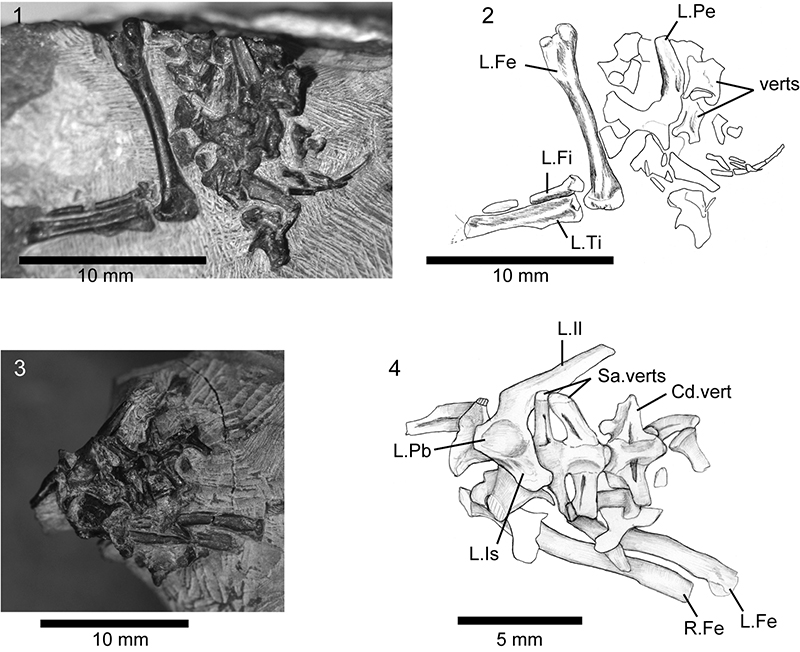
FIGURE 12. The phylogenetic position of Asagaolacerta tricuspidens gen. et sp. nov. tested using different analytical protocols within TNT. 1, detail of Strict Consensus of 1000 trees using the protocol that yielded the tree in Figure 5.1, node support values Bremer/Jacknife/Symmetric sampling; 2, one of three trees resulting from an analysis using with the molecular backbone constraint tree, but no character ordering or weighting; 3, one of 58 trees from an analysis run as in (2), but with character ordering as per Gauthier et al. (2012) and Longrich et al. (2012), and Implied Weighting (k=7); 4, one of 34 trees from an analysis run as in (3), but without the molecular backbone constraint; 5, 70% MRT of 19 trees resulting from an analysis (characters ordered but equally weighted, no constraints) run using only the boreoteiioid taxa, with Gekko gecko as the outgroup taxon. The 70%MRT is presented rather than the unresolved Strict Consensus to show that Asagaolacerta tricuspidens is usually (88% of trees) placed in the basal position (see text for further discussion). Abbreviation: poly, polyglyphanodont (as used in Longrich et al., 2012).
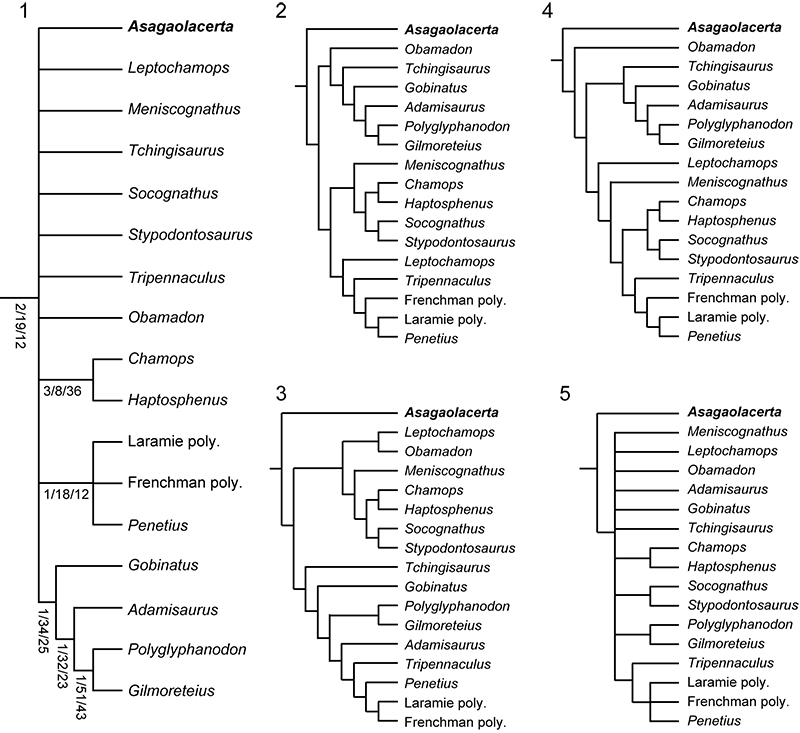
FIGURE 13. Hakuseps imberis gen. et sp. nov., holotype left dentary, SBEI 2086. 1-2, labial view; 3-4, lingual view; 5-6, occlusal view; and 7, ventral view. For abbreviations, see Material and Methods.
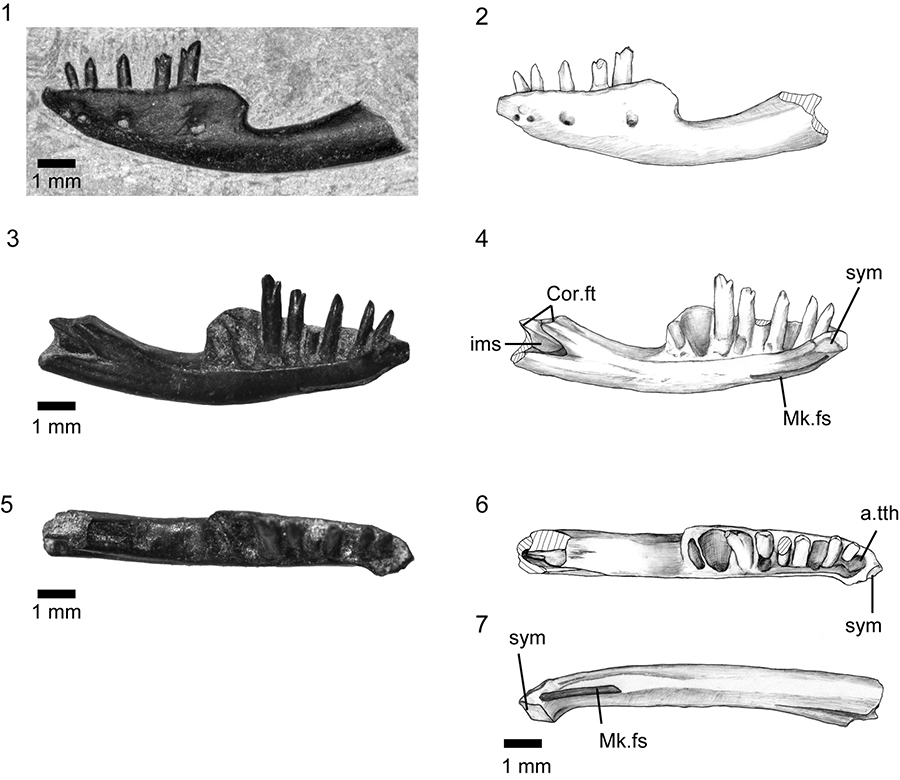
FIGURE 14. Comparison of Pachygenys thlastesa (Gao and Cheng, 1999) and Hakuseps imberis gen. et sp. nov. Labial and lingual views respectively of 1-2, Pachygenys thlastesa (redrawn from Gao and Cheng, 1999), 3-4, Hakuseps imberis gen. et sp. nov.
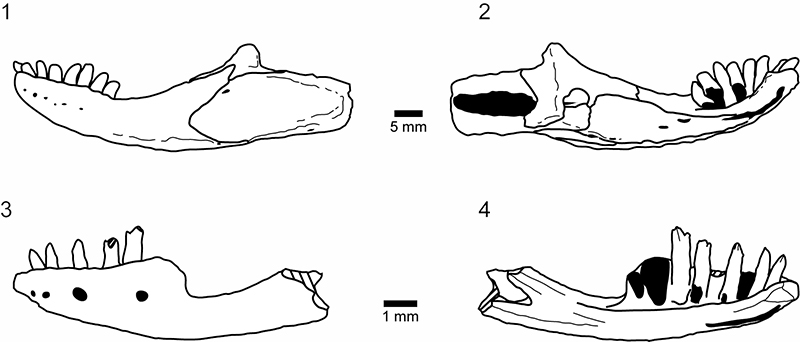
FIGURE 15. Shiramine Morphotype A, bicuspid dentition. 1-2, left maxilla, SBEI 1525 in labial view; 3-4, right maxilla, SBEI 1501, in labial view; 5-10, left dentary, SBEI 808, in two parts, in labial view, with 5-6, symphysial region, 7-8, posterior dentary, and 9-10, detail of bicuspid teeth.
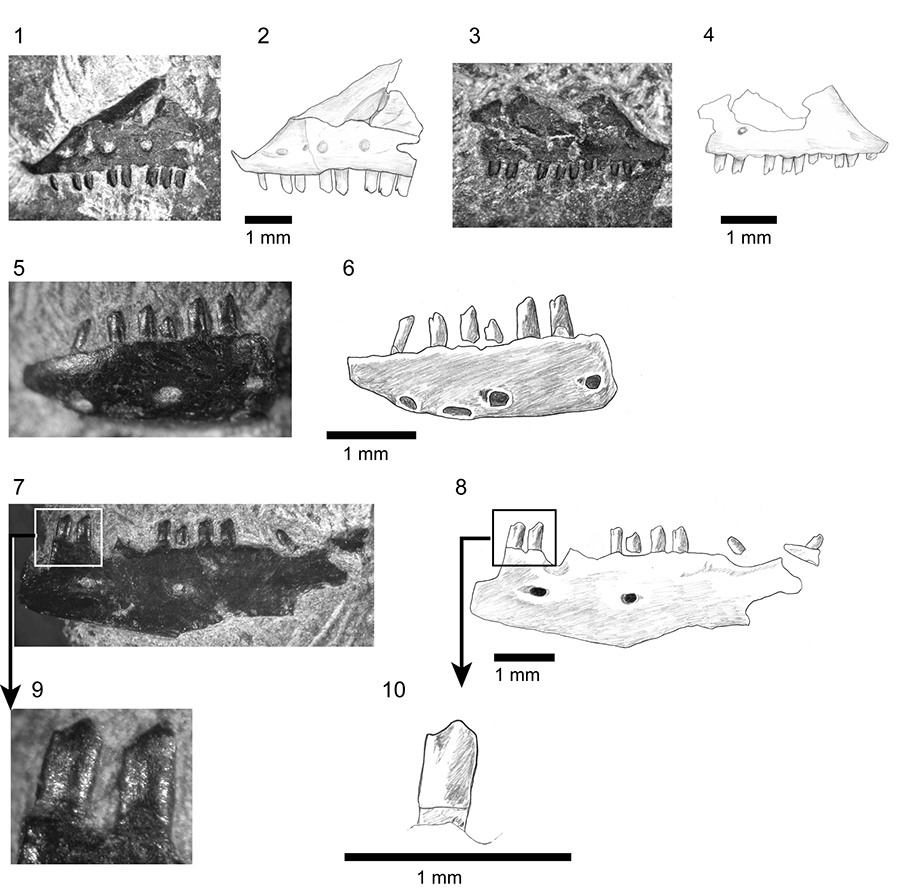
FIGURE 16. Shiramine Morphotype B, right mandible SBEI 827. 1-2, dentary in labial view; 3-4, dentary and associated splenial in lingual view; 5-11, postdentary bones in 5-6 dorsomedial view; 7, ventral view; 8, dorsal view; 9, medial view; and 10, posterior view of articular surface. For abbreviations, see Material and Methods.
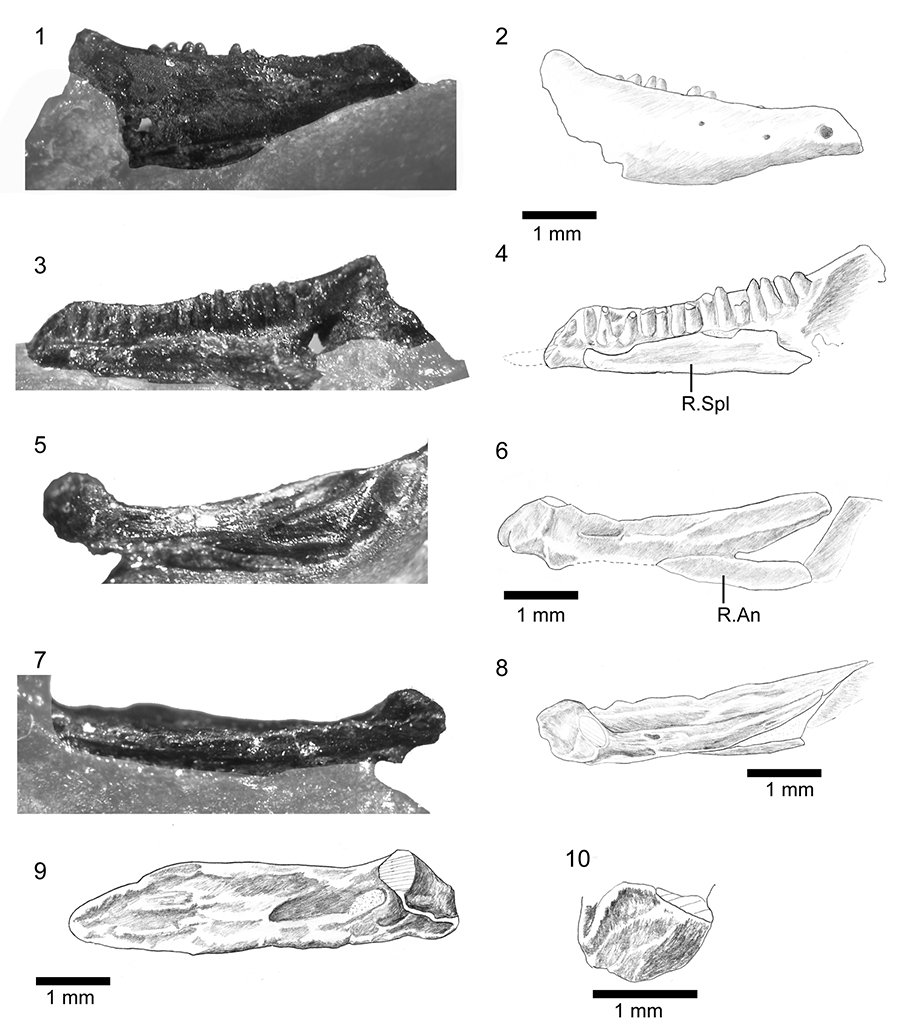
FIGURE 17. Comparison of Shiramine Morphotype B and Myrmecodaptria microphagosa (Gao and Norell, 2000) from the Late Cretaceous of Mongolia. 1-2, left lateral and dorsal views respectively of the skull of Myrmecodaptria microphagosa, redrawn from Gao and Norell (2000). Scale bar equals 5mm; 3, dentary of morphotype B, SBEI 827, labial view; 4, isolated left maxilla, SBEI 2407, in labial view; 5, isolated median frontal, SBEI 1803, in dorsal view.
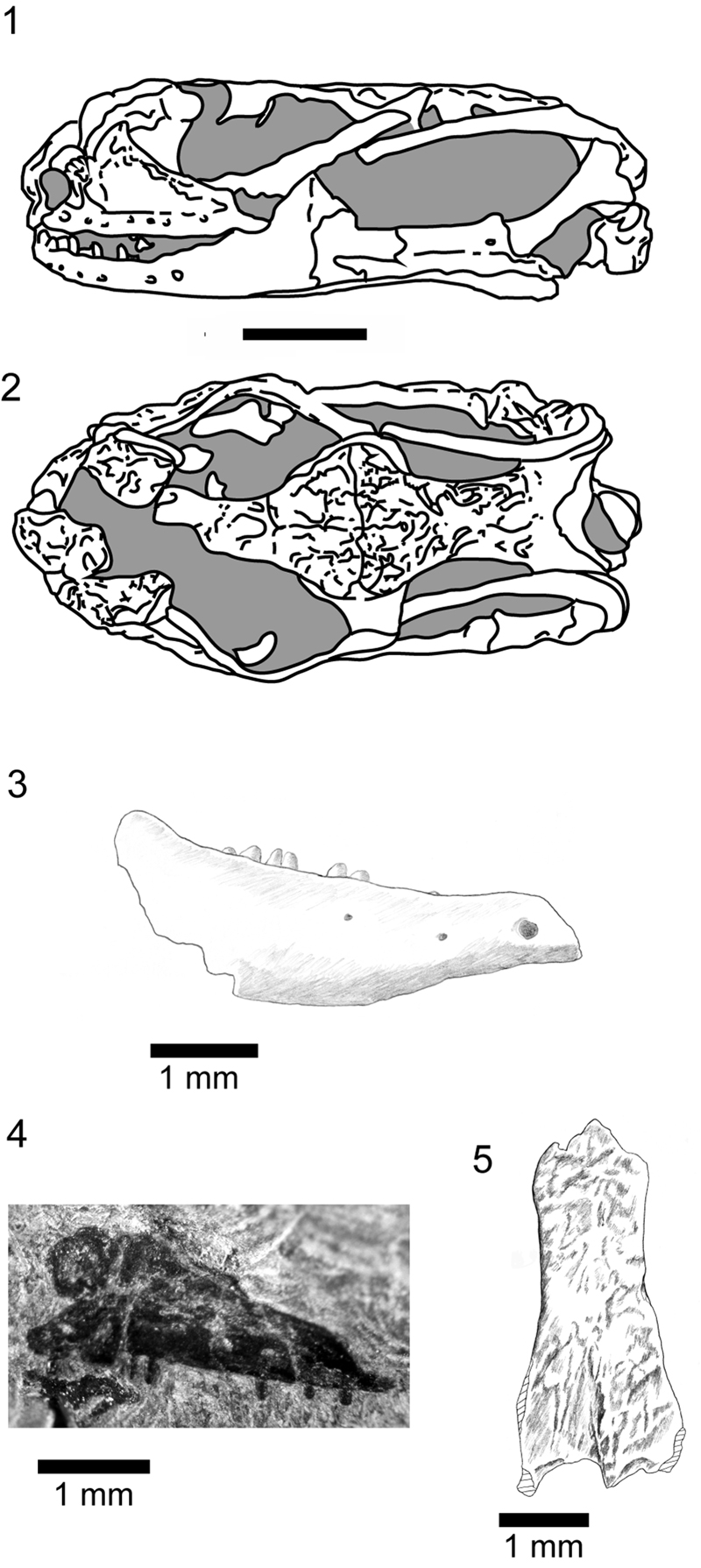
FIGURE 18. Shiramine Morphotype C, SBEI 1277, posterior region of a left dentary in 1-2, labial view.

 Susan E. Evans
Susan E. Evans
Department of Cell and Developmental Biology
University College London
Gower Street
London WC1E 6BT
UK
ucgasue@ucl.ac.uk (corresponding author)
Susan E. Evans completed a Ph.D. in Vertebrate Palaeontology in 1977 at University College London. After a period in the Arabian Gulf, she joined the Middlesex Hospital Medical School in 1984, and rejoined UCL in 1986 when the medical schools merged. She is currently Professor of Vertebrate Morphology and Palaeontology, in the Department of Cell and Developmental Biology at UCL. Her primary research interests are in the evolution, systematics, and functional morphology of reptiles and amphibians.

 Ryoko Matsumoto
Ryoko Matsumoto
Kanagawa Prefecture Museum of Natural History
499 Iryuuda
Odawara
Kanagawa Prefecture, 250-0035
Japan
ryokosaur@gmail.com
Ryoko Matsumoto obtained her Ph.D. at University College London in 2011. After completion, she spent one year at the National Museum of Nature and Science, Tokyo, on a JSPS (Japan Society for the Promotion of Science) Postdoctoral Fellowship. She is currently Curator of Herpetology at the Kanagawa Prefectural Museum of Natural History, Japan. Her main field of study is the functional anatomy and phylogenetic relationships of Mesozoic and Cenozoic amphibians and reptiles (especially choristoderes).
An assemblage of lizards from the Early Cretaceous of Japan
The Early Cretaceous deposits of western Japan are roughly the same age as the more famous feathered dinosaur deposits of China. The most productive of the Japanese localities is the "Fossil Cliff" in the village of Shiramine. To date, this locality has yielded a diverse wetland vertebrate assemblage including fish, amphibians, dinosaurs, birds, turtles, choristoderes, mammals, and lizards. Three of the lizards, Kaganaias hakusanensis, Kuwajimalla kagaensis, and Sakurasaurus shokawensis, have been described elsewhere. Here we describe further lizard material from the locality, three of which are given new scientific names. Together with the species already described, the new material demonstrates the presence of nine or ten different lizards in the Early Cretaceous deposits of Japan, a diversity rivalling the assemblages of contemporaneous or near-contemporaneous deposits in North America, Europe, and other parts of Asia.
Resumen en Español
Una asociación de lagartos del Cretácico Inferior de Japón
Los depósitos del Cretácico Inferior del Grupo Tetori del oeste de Japón han proporcionado una fauna diversa de vertebrados de humedales, tanto componentes acuáticos como terrestres. Estos últimos incluyen varios lagartos, tres de los cuales han sido nominados y descritos en detalle: Kuwajimalla kagaensis, un herbívoro del grupo Borioteiioidea, y Sakurasaurus shokawensis, un pariente del género Yabeinosaurus de la paleobiota china de Jehol. Aquí se describen ejemplares de lagarto de la localidad de Shiramine que representan cinco o seis taxones adicionales, tres de los cuales se nombran en este documento: un pequeño lagarto representado por dos asociaciones, pero con una posición filogenética no resuelta; un lagarto ligeramente más grande con dientes de tres cúspides que está relacionado con Borioteiioidea; y un lagarto extraño con dientes de dos cúspides representado por una sola mandíbula, aunque esta es única en su morfología. Los tres morfotipos de lagarto adicionales quedan sin nombrar. Uno tiene dientes con dos cúspides en maxilares no especializados. El segundo tiene dientes pequeños monocúspide en un dentario con un proceso coronoides profundo y que es parecido al dentario de la enigmática especie del Cretácico Superior de Mongolia denominada Myrmecodaptria microphagosa. El tercer morfotipo está representado por un único ejemplar fragmentario que tiene pequeños dientes en una mandíbula profunda. Juntos, los lagartos de Kuwajima forman una asociación filogenética y morfológicamente diversa.
Palabras clave: Squamata; nuevo género; nuevas especies; Japón; Asia; Cretácico
Traducción: Enrique Peñalver
Résumé en Français
Un assemblage de lézards du Crétacé ancien du Japon
Les dépôts du Crétacé ancien du groupe de la Tetori, dans l'ouest du Japon, ont livré une faune diverse de vertébrés de zones humides, incluant à la fois des composantes aquatique et terrestre. Cette dernière inclut plusieurs lézards, dont trois ont été nommés et décrits en détail : Kaganaias hakusanensis, un lézard aquatique à corps allongé ; Kuwajimalla kagaensis, un Borioteiioidea herbivore ; et Sakurasaurus shokawensis, apparenté à Yabeinosaurus, un genre provenant du Jehol de Chine. Dans cet article, nous décrivons du matériel de lézards provenant de la localité de Shiramine et représentant cinq ou six taxons supplémentaires, dont trois sont nommés dans cet article : un lézard de petite taille représenté par deux ensembles de spécimens associés, mais dont la position phylogénétique n'est pas résolue ; un lézard de taille légèrement plus grande, avec des dents tricuspides, apparenté aux Borioteiioidea ; et un lézard bizarre à dents bicuspides, représenté par une seule mâchoire dont la morphologie est cependant unique. Les trois morphotypes supplémentaires de lézards ne sont pas nommés. Le premier d'entre eux présente des dents bicuspides mais des mâchoires non spécialisées. Le second a de petites dents unicuspides implantées dans un os dentaire à processus coronoïde profond et ressemblant à l'os dentaire de Myrmecodaptria microphagosa, une espèce énigmatique du Crétacé récent de Mongolie. Le troisième morphotype est représenté par un seul spécimen fragmentaire qui présente de petites dents implantées dans une mâchoire profonde. Dans l'ensemble, les lézards de Kuwajima forment un assemblage à morphologies et positions phylogénétiques diverses.
Mots-clés : Squamata ; nouveau genre ; nouvelle espèce ; Japon ; Asie ; Crétacé
Translator: Antoine Souron
Deutsche Zusammenfassung
Eine Assemblage von Eidechsen aus der frühen Kreide von Japan
Die frühkretazischen Ablagerungen der Tetori-Gruppe von Westjapan haben eine diverse Feuchtgebiet-Wirbeltierfauna hervorgebracht, die sowohl aquatische als auch terrestrische Komponenten enthält. Letztere beinhaltet einige Eidechsen von denen drei benannt und im Detail beschrieben wurden: Kaganaias hakusanensis, eine aquatische Eidechse mit länglichem Körper; Kuwajimalla kagaensis, ein herbivorer Borioteiioide und Sakurasaurus shokawensis, ein Verwandter der chinesischen Jehol-Gattung Yabeinosaurus. Hier beschreiben wir das Eidechsenmaterial aus der Shiramine-Lokalität, vertreten mit fünf oder sechs zusätzlichen Taxa, von denen drei hier benannt sind: eine kleine Eidechse die mit zwei kleinen Gesellschaften vertreten ist, deren phylogenetische Position aber ungelöst ist; eine etwas größere Eidechse mit tricuspiden Zähnen, die mit den Borioteiioiden verwandt ist; und eine bizarre Eidechse mit bicuspiden Zähnen, die mit einem einzigen aber morphologisch einzigartigem Kiefer vertreten ist. Die drei zusätzlichen Eidechsen-Morphotypen sind unbenannt. Eine hat bicuspide Zähne aber unspezialisierte Kiefer. Die zweite hat kleine unicuspide Zähne in einem Dentale mit tiefem Coronoidfortsatz, das dem Dentale der enigmatischen spätkretazischen Myrmecodaptria microphagosa aus der Mongolei ähnelt. Der dritte Morphotyp ist durch ein einziges fragmentarisches Stück repräsentiert und hat kleine Zähne in einem tiefen Kiefer. Zusammen bilden die Kuwajima-Eidechsen eine phylogenetisch und morphologisch diverse Assemblage.
Schlüsselwörter: Squamata; neue Gattung; neue Art; Japan; Asien; Kreide
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024