

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
 David W. E. Hone. School of Biological and Chemical Sciences, Queen Mary University of London, Mile End Road, E1 4NS. d.hone@qmul.ac.uk
David W. E. Hone. School of Biological and Chemical Sciences, Queen Mary University of London, Mile End Road, E1 4NS. d.hone@qmul.ac.uk
Dave Hone received his B.SC. from the University of Bristol, an M.Sc. from Imperial College and his Ph.D. from the University of Bristol. He took postdoctoral positions in the Bavarian State Collection for Palaeontology, Munich and the Institute of Vertebrate Paleontology in Beijing. He is currently a Lecturer in Zoology at Queen Mary University of London and specialises in the behaviour and ecology of the non-avian dinosaurs and the palaeobiology of the pterosaurs.

 Dylan Wood. School of Biological and Chemical Sciences, Queen Mary University of London, Mile End Road, E1 4NS. d.wood@se12.qmul.ac.uk
Dylan Wood. School of Biological and Chemical Sciences, Queen Mary University of London, Mile End Road, E1 4NS. d.wood@se12.qmul.ac.uk
Dylan Wood is a masters student at Queen Mary, University of London, BSc (Hons) Zoology. Interested in evolutionary biology, sexual selection and sexual dimorphism, with current focus on Mesozoic archosaurs.
 Robert J. Knell. School of Biological and Chemical Sciences, Queen Mary University of London, Mile End Road, E1 4NS. r.knell@qmul.ac.uk
Robert J. Knell. School of Biological and Chemical Sciences, Queen Mary University of London, Mile End Road, E1 4NS. r.knell@qmul.ac.uk
Rob Knell is the Reader in Evolutionary Ecology at Queen Mary, University of London having completed his BSc at Imperial College and his PhD at the University of Liverpool. His research interests include mating system evolution and its consequences for animal evolution and ecology, in both extant and extinct species, and also population ecology and adaptation to changing environments.
FIGURE 1. Size categories of specimens of Protoceratops andrewsi used in this study. Right to left: young juvenile, juvenile, subadult, adult. Scale bar is 1 m. Image modified from Hone et al. (2014a), original illustration by David Maas.

FIGURE 2. Changes in skull shape in Protoceratops andrewsi. All skulls are drawn to the same total length and are seen in dorsal view (upper row) and right lateral view (lower row). Left to right (with sources in parentheses) small juveniles (Fastovsky et al., 2011), juveniles (MPC-D 100/526), subadults (MPC-D 100534), putative ‘female’ morph, putative ‘male’ morph (both Dodson, 1976). The large fenestrae seen in the smallest animals are supratemporal fenestra and are not homologous with the frills of the fenestra in the larger animals.
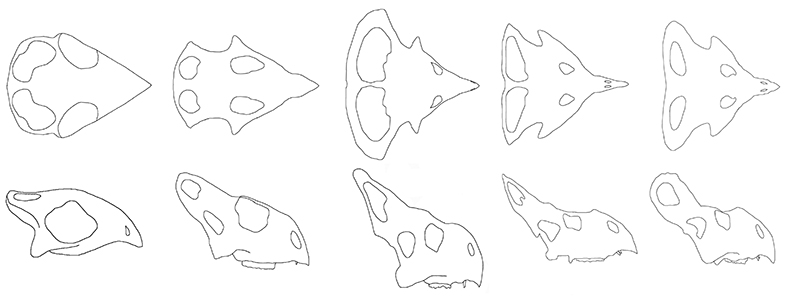
FIGURE 3. Measurements taken from skulls of Protoceratops based on an idealised adult in dorsal view (above) and lateral view (below). Black lines and numbers indicate the measurements taken according to the variable of Dodson (1976). These are: 1, basal skull length; 2, total length (frill length is variable 2 subtracted from variable 1); 8, jugal width; 9, frill width; 13, orbit length; 14, orbit height. The grey lines indicate the maximum and minimum lengths of the frill as measured in juvenile animals. See text for further details.
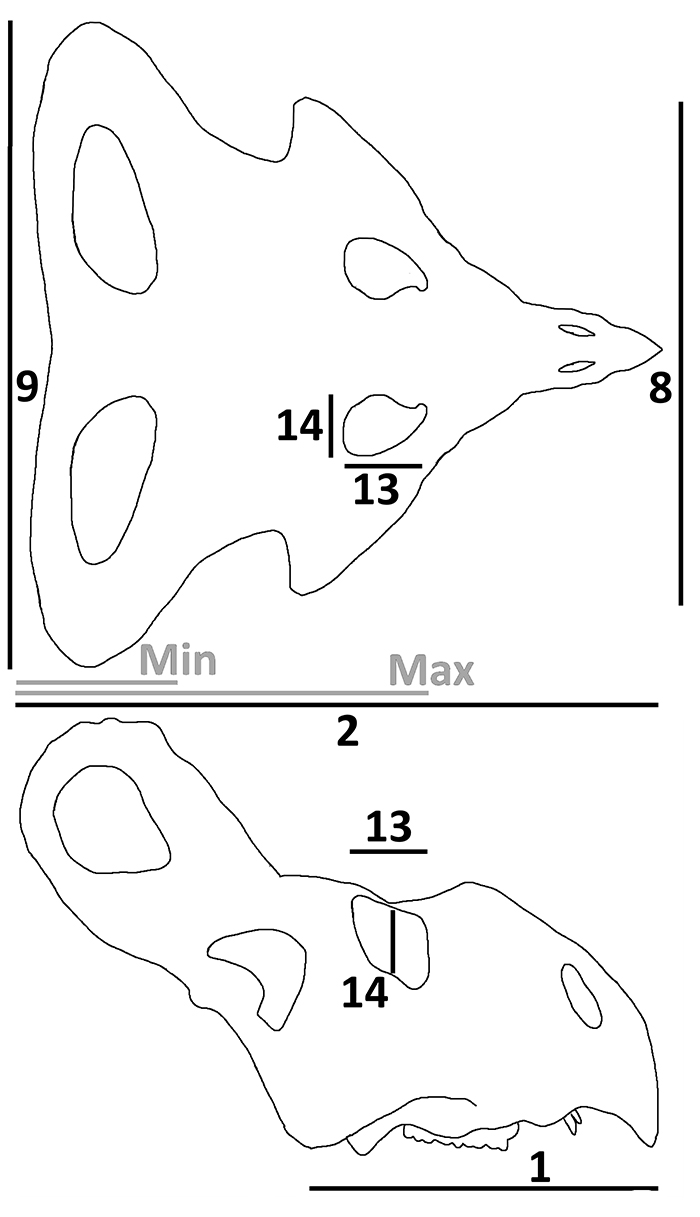
FIGURE 4. Allometric relationships for frill length ( 1), frill width ( 2) and the width across the jugal bosses ( 3). Solid lines show the fitted lines from SMA regression, dashed grey lines show the line of isometry (slope = 1 and intercept = 0). All measurements were originally in mm prior to log transformation.
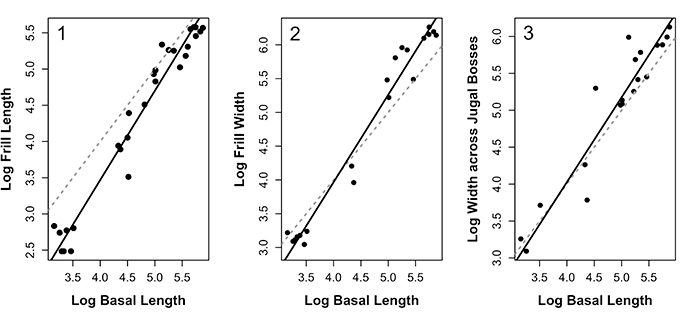
FIGURE 5. Life restoration of adult Protoceratops andrewsi (foreground) engaging in speculative display postures, an activity in which non-mature animals (background) do not take part. Artwork by Rebecca Gelernter, who retains the copyright on this image–used with permission.

TABLE 1. SMA slopes and associated statistics for frill and other skull traits from Protoceratops andrewsi.
| Measure | Intercept | Slope | Slope CIs | Test statistic | P slope ≠ 1 |
| Frill length | -1.46 | 1.23 | 1.14-1.34 | r = 0.719, 26df | <0.0001 |
| Frill width | -1.21 | 1.29 | 1.19-1.41 | r = 0.827, 18df | <0.0001 |
| Jugal Boss width | -0561 | 1.15 | 0.983-1.34 | r = 0.403, 18df | 0.078 |
| Orbit width | 0.272 | 0.699 | 0.650-0.751 | r = -0.877, 30df | <0.0001 |
| Orbit height | -0.583 | 0.843 | 0.760-0.935 | r = -0.526, 30df | 0.002 |
APPENDIX
Table of all data used in the analyses. All measurements are in mm. AMNH = American Museum of Natural History, New York; MPC/D = Mongolian Paleontological Centre, Ulan Baator.
Positive allometry for exaggerated structures in the ceratopsian dinosaur Protoceratops andrewsi supports socio-sexual signaling
Plain Language Abstract
Many dinosaurs had large and often elaborate bony crests on their heads. The function of these crests has been much discussed but little consensus has been reached. Here we analyse the changes in the shape of the large 'frill' on the head of specimens of a small horned dinosaur from Mongolia called Protoceratops. We show that the frill started at a small size in young animals and only grew large when the animal was close to maturity. Therefore, the frill must have a function that operates in adults, but not juveniles. Such growth patterns are typical for features under sexual selection where animals compete for the best mates. We conclude that the frill was likely a structure used by adults in dominance contests.
Resumen en Español
La alometría positiva para estructuras exageradas en los dinosaurios ceratópsidos Protoceratops andrewsi corrobora la señalización socio-sexual
Se evalúa la alometría en los salientes del collar óseo y del yugal de los pequeños dinosaurios ornitisquios Protoceratops andrewsi. El análisis de 37 ejemplares, que abarca cuatro clases de tamaños diferentes de animales, muestra que el collar (en longitud y anchura) y yugal de esta especie experimentan una alometría positiva durante la ontogenia. Esto apoya, junto con otros datos, que estas características estaban bajo selección como señales de dominación socio-sexuales.
Palabras clave: ceratópsidos; selección sexual; comportamiento; gregarismo
Traducción: Enrique Peñalver
Résumé en Français
L'allométrie positive des structures disproportionnées du dinosaure cératopsien Protoceratops andrewsi supporte un signal socio-sexuel
L'allométrie de la collerette et des protubérances jugales du petit dinosaure ornitischien Protoceratops andrewsi est examinée. Une analyse de 37 spécimens, englobant quatre classes de taille distinctes, montre que la collerette (à la fois en termes de longueur et de largeur) et les jugaux de cette espèce suivent une allométrie positive pendant l'ontogénie. En accord avec d'autres données, cela supporte l'hypothèse selon laquelle ces caractéristiques sont soumises à la sélection en tant que signaux de dominance socio-sexuelle.
Mots-clés : Ceratopsia ; sélection sexuelle ; comportement ; grégarisme
Translator: Kenny J. Travouillon or Antoine Souron
Deutsche Zusammenfassung
Positive Allometrie von verlängerten Strukturen bei dem ceratopsiden Dinosaurier Protoceratops andrewsi unterstützt die sozio-sexuelle Signalgebung
Die Allometrie des Nackenschildes und der Wangenhörner des kleinen Vogelbeckensauriers Protoceratops andrewsi wird bewertet. Eine Analyse von 37 Stücken, die vier klar getrennte Größengruppen unterscheidet, zeigt dass das Nackenschild (sowohl in der Länge als auch in der Breite) und die Wangen dieser Art während der Ontogenie eine positive Allometrie erfahren. Zusammen mit anderen Daten unterstützt dies die Annahme, dass diese Merkmale als sozio-sexuelle Dominanzsignale unter Selektion waren.
Schlüsselwörter: Ceratopsier; sexuelle Selektion; Verhalten; Herdentier
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024