

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
 Barbara Rzebik-Kowalska. Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Sławkowska 17, 31-016 Kraków, Poland rzebik@isez.pan.krakow.pl
Barbara Rzebik-Kowalska. Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Sławkowska 17, 31-016 Kraków, Poland rzebik@isez.pan.krakow.pl
Barbara Rzebik-Kowalska is a professor in the Institute of Systematics and Evolution of Animals at the Polish Academy of Sciences in Kraków (Poland). She graduated from the Jagiellonian University (Kraków) and received her PhD on the ground of the thesis "Studies on the diet of the carnivores in Poland" in 1973. She has published numerous papers and books concerning recent and fossil (Miocene – Quaternary) mammals (especially insectivore mammals) of Europe, as well as of the Middle and Far East, the Greater Antilles, and of North Africa. For five years (1979-1983) she worked with her husband, Kazimierz Kowalski, in Algeria where they lectured and collected extensive materials of mammals publishing the book "Mammals of Algeria" in 1991.

 Leonid I. Rekovets. University of Environmental and Life Sciences of Wrocław, Chełmoński 38c, 51-630 Wrocław, Poland
Leonid I. Rekovets. University of Environmental and Life Sciences of Wrocław, Chełmoński 38c, 51-630 Wrocław, Poland
and
National Museum of Natural History, B.Khmelnicki 15, 01030 Kiev, Ukraine leonid.rekovets@up.wroc.pl
Leonid I. Rekovets is a professor in the University of Environmental and Life Sciences of Wroclaw (Poland) and in the Institute of Zoology and Museum of History and Nature in Kiev (Ukraine). He graduated from the Gogol University in Nizhyn (Ukraine), faculty Biology. He specializes in paleotheriology (faunistics and systematics of small mammals of the Neogene and the Pleistocene of Europe) as well as in biostratigraphy and paleogeography. He is the author of five monographs and 200 scientific articles. He has found and studied a lot of localities of theriofauna in Ukrainie.
FIGURE 1. The Ukrainian localities in which insectivore mammals were studied. 1, Regional map of Europe indicating location of Ukraine (Inset, 2). 2-3, Ukraine Miocene to Pleistocene localities: Inset (3), Miocene localities (A-Popovo 3; B-Verkhnya Krynitsa 2; C-Lobkove), Pliocene localities (A-Popovo 2; B- Verkhnya Krynitsa 1), Plio/Pleistocene locality (A-Popovo 1); Pleistocene locality (A-Popovo 0; D- Medzhybozh).
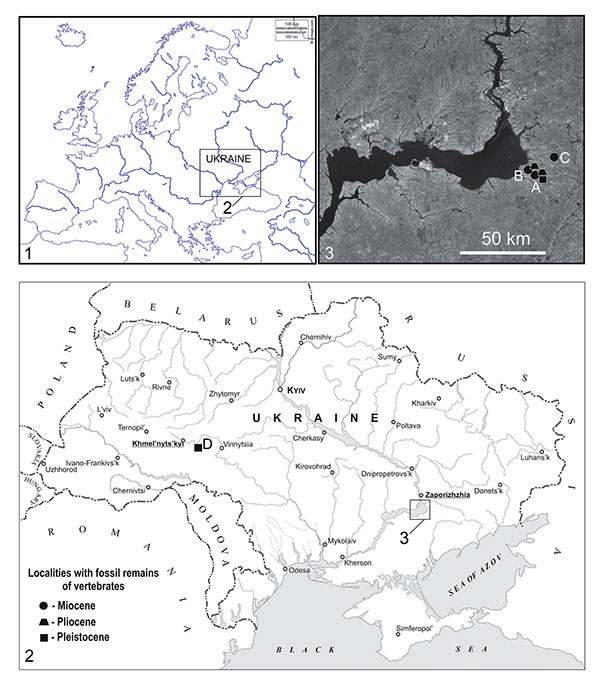
FIGURE 2. Schizogalerix sp. from Popovo 3 (1) and Verkhnya Krynitsa 2 (2-5). 1, right P4, occlusal view (catalogue number 29/III/1/1). (2-4) left m1; 2, occlusal; 3, lingual; 4, posterior view (c.n. 29/2/1/1); 5, right m2; occlusal view (c.n. 29/2/1/2).
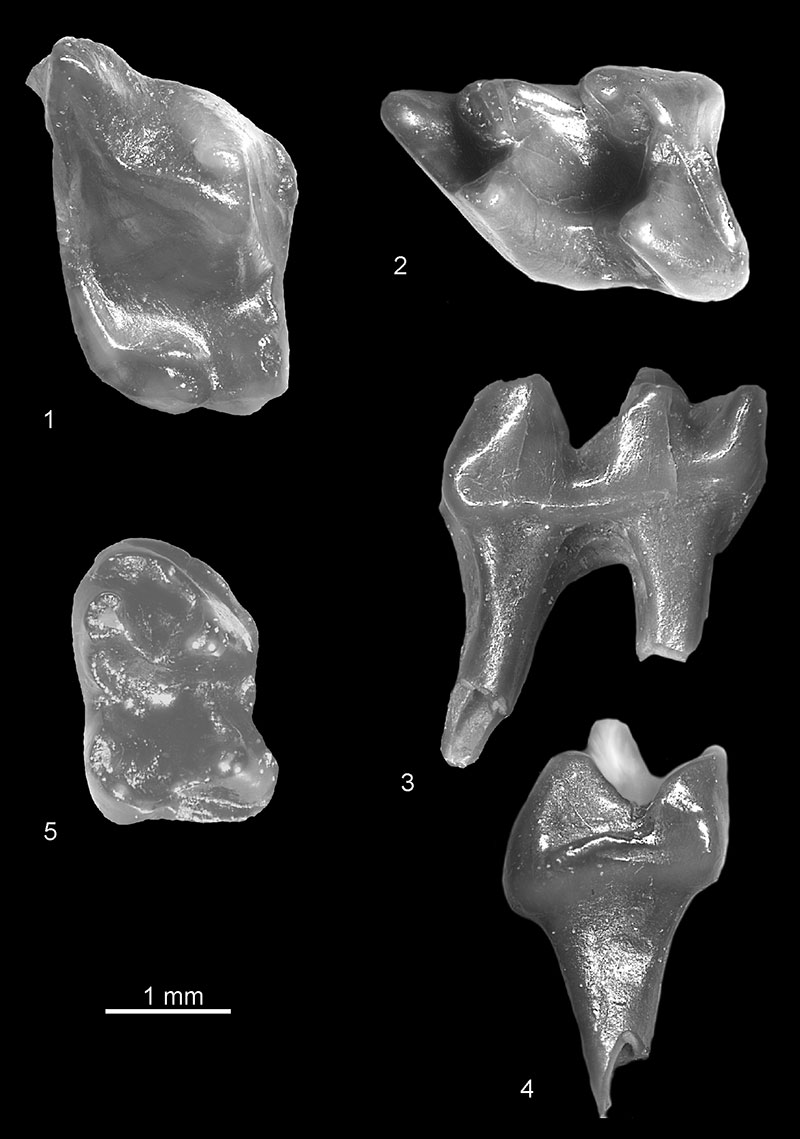
FIGURE 3. 1, cf. Mygalinia hungarica from Verkhnya Krynitsa 2, left m2, occlusal view (c.n. 29/2/2/1); 2, Ruemkelia sp. from Verkhnya Krynitsa 2, fragment of left m2, occlusal view (c.n. 29/2/3/1).
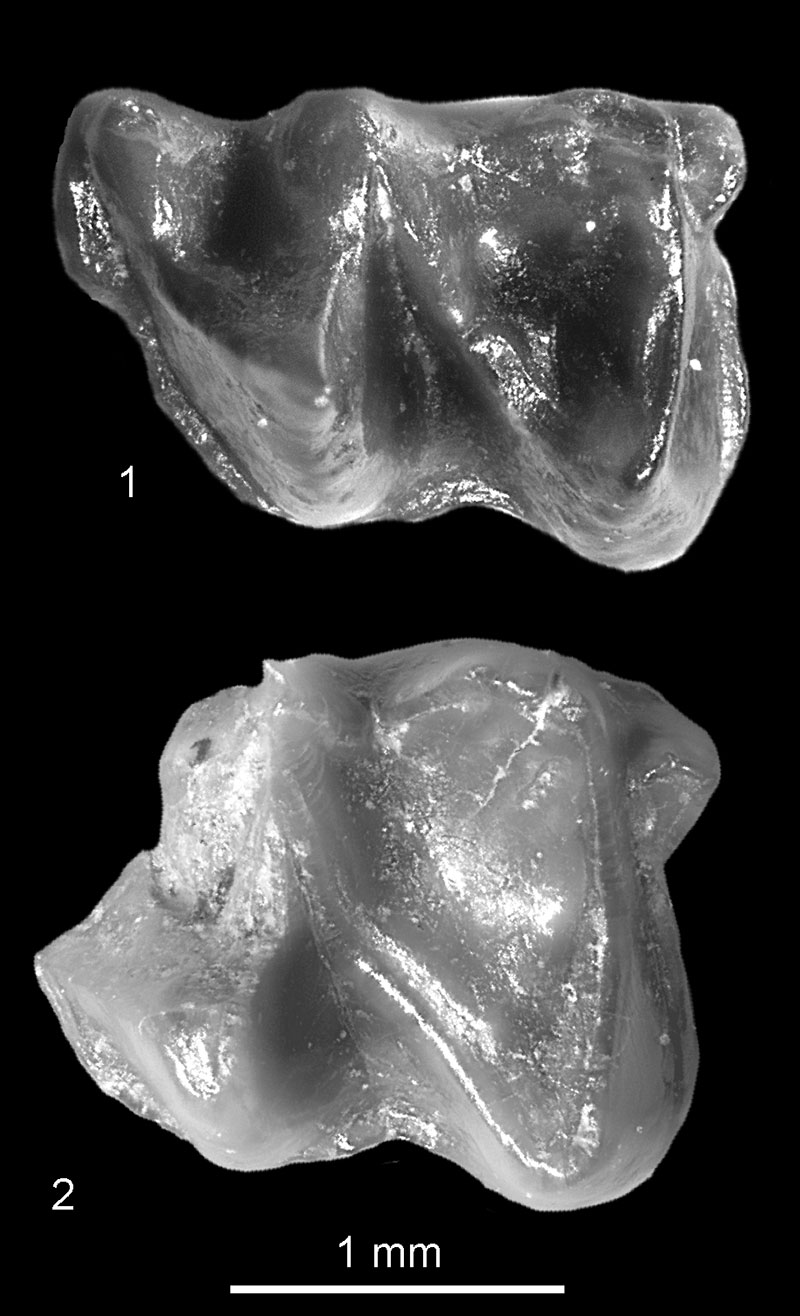
FIGURE 4. Desmana sp. from Verkhnya Krynitsa 1 (1-2) and from Popovo 2 (3-7). Left ?I2, 1, buccal; 2, occlusal view (c.n. 29/1/4/1). Left C: 3, buccal; 4, occlusal view (c.n. 29/II/4/1). 5, Left P2, occlusal view (c.n. 29/II/4/2). 6, Left P4, occlusal view (c.n. 29/II/4/3). 7, Right M1, occlusal view (c.n. 29/II/4/4).
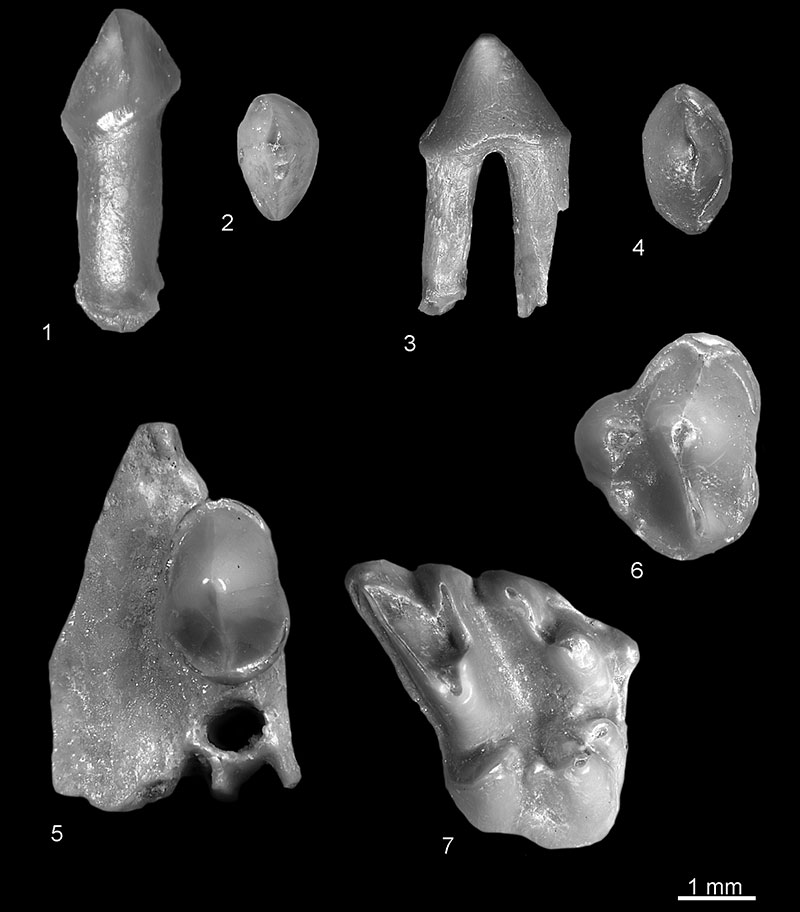
FIGURE 5. Desmana sp. from Popovo 2 (1-4 and 6) and Popovo 1 (5). Left ?i3: 1, buccal, 2, occlusal view (c.n. 29/II/4/5). Left p2: 3, buccal; 4, occlusal view (c.n. 29/II/4/6). 5, Left m1, occlusal view (c.n. 29/I/4/1). 6, Left m3, occlusal view (c.n. 29/II/4/7).

FIGURE 6. 1, Desmana sp. from Popovo 1 (1). fragment of right mandible with m1-m3 in buccal view (c.n. 29/I/4/2); (2-3) cf. Desmana sp. from Verkhnya Krynitsa 2, left humerus, 2, dorsal side, 3, ventral side (c.n. 29/2/5/1).
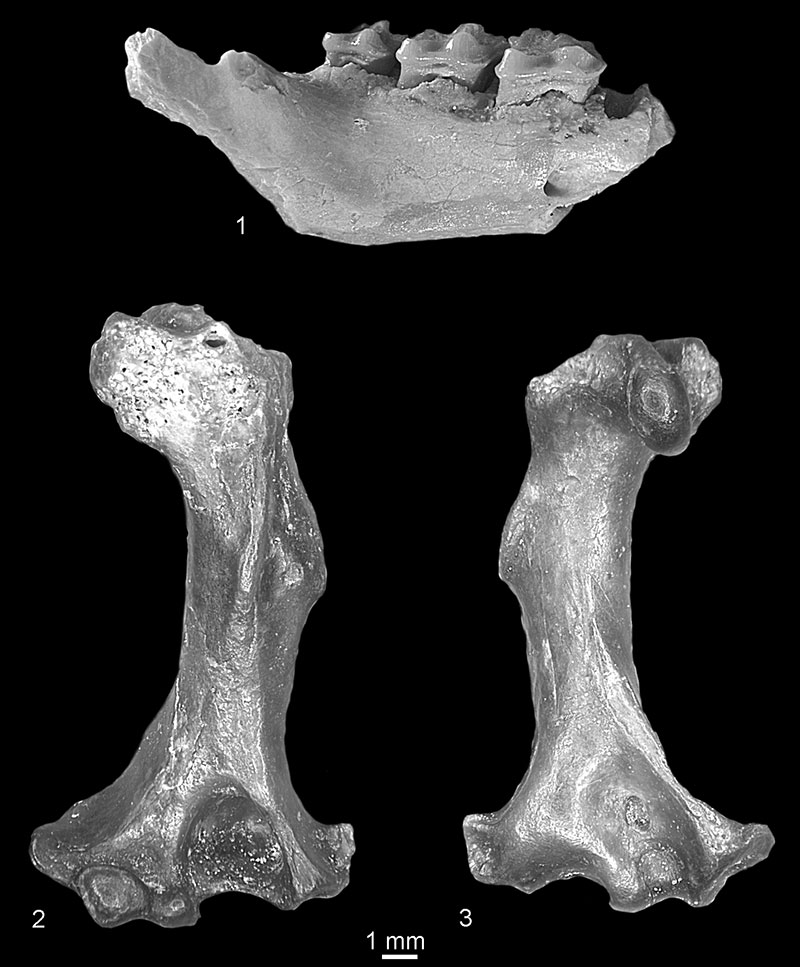
FIGURE 7. 1, cf. Miosorex sp. from Verkhnya Krynitsa 2, right I1, buccal view (c.n. 29/2/6/1). 2, Miosorex grivensis from Popovo 3, fragment of right mandible with m2 and trigonid of m3, buccal view (c.n. 29/III/7/1).
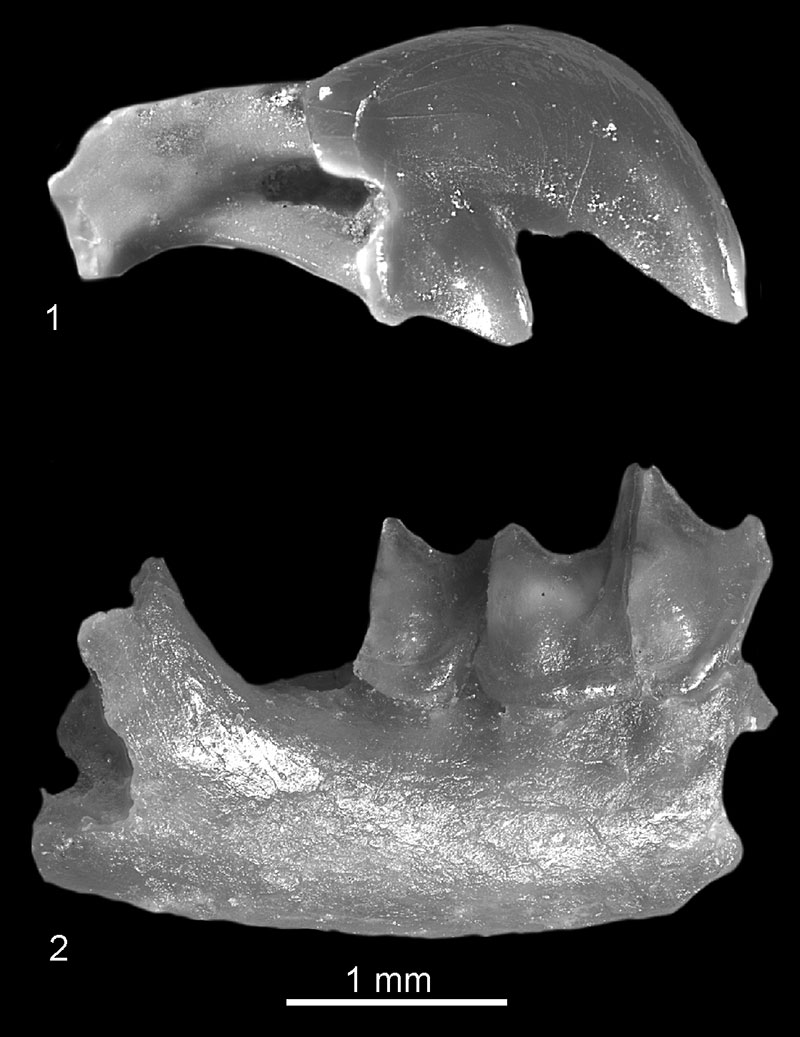
FIGURE 8. Crusafontina cf. kormosi from Verkhnya Krynitsa 2. 1, fragment of left mandible with i1-p4 in buccal view (c.n. 29/2/8/1); 2, fragment of left mandible with m1-m2 in occlusal view (c.n. 29/2/8/2); 3, fragment of left mandible with m2-m3 and coronoid and condyloid proceses in lingual view (c.n. 29/2/8/3). (4-5), Fragment (talonid) of right m2 in: 4, occlusal; and 5, lingual view (c.n. 29/2/8/5).
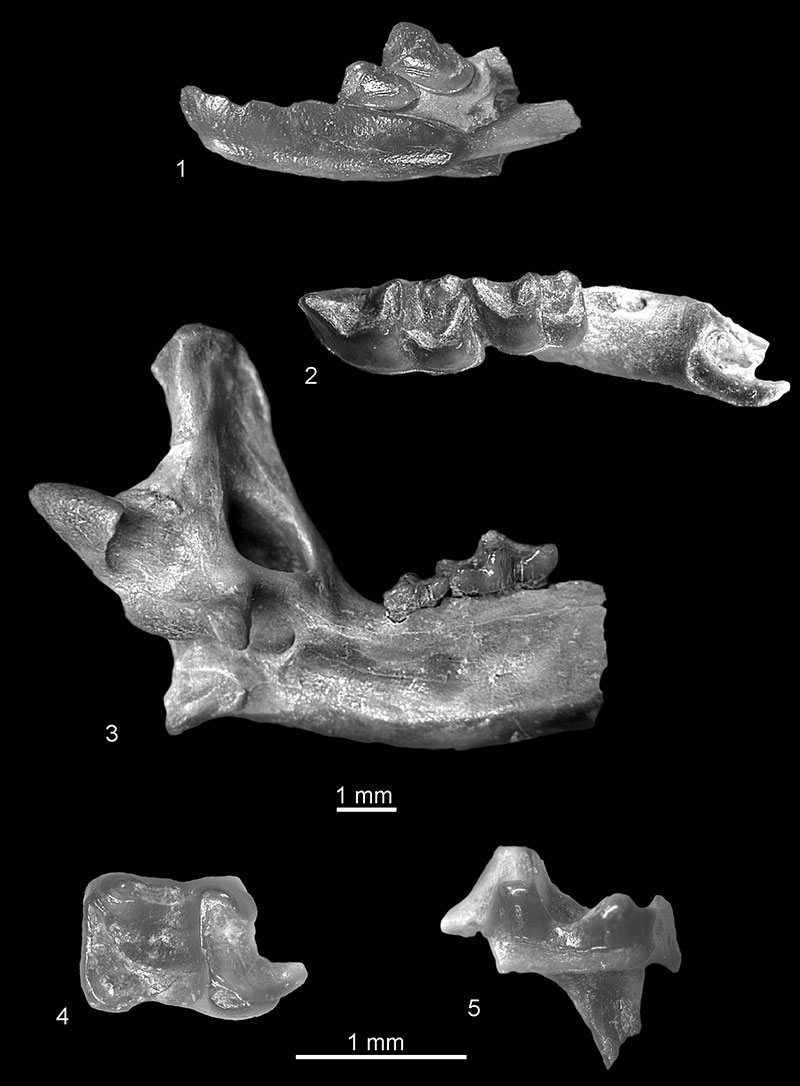
FIGURE 9. (1-2) cf. Asoriculus sp. from Popovo 3: 1, fragment of right mandible with coronoid process, buccal view (c.n. 29/III/9/1). 2, fragment of right mandible with m2-m3, lingual view (c.n. 29/III/9/2). (3) Neomysorex alpinoides from Verkhnya Krynitsa 2, fragment of right mandible with p4-m3, lingual view (c.n. 29/2/10/1), (4-6) Neomys newtoni from Medzhybozh: 4, left condyloid process, posterior view (c.n. 29/4/11/2); fragment of left mandible with m1-m2, in 5, lingual, and 6, occlusal view (c.n. 29/4/11/1).
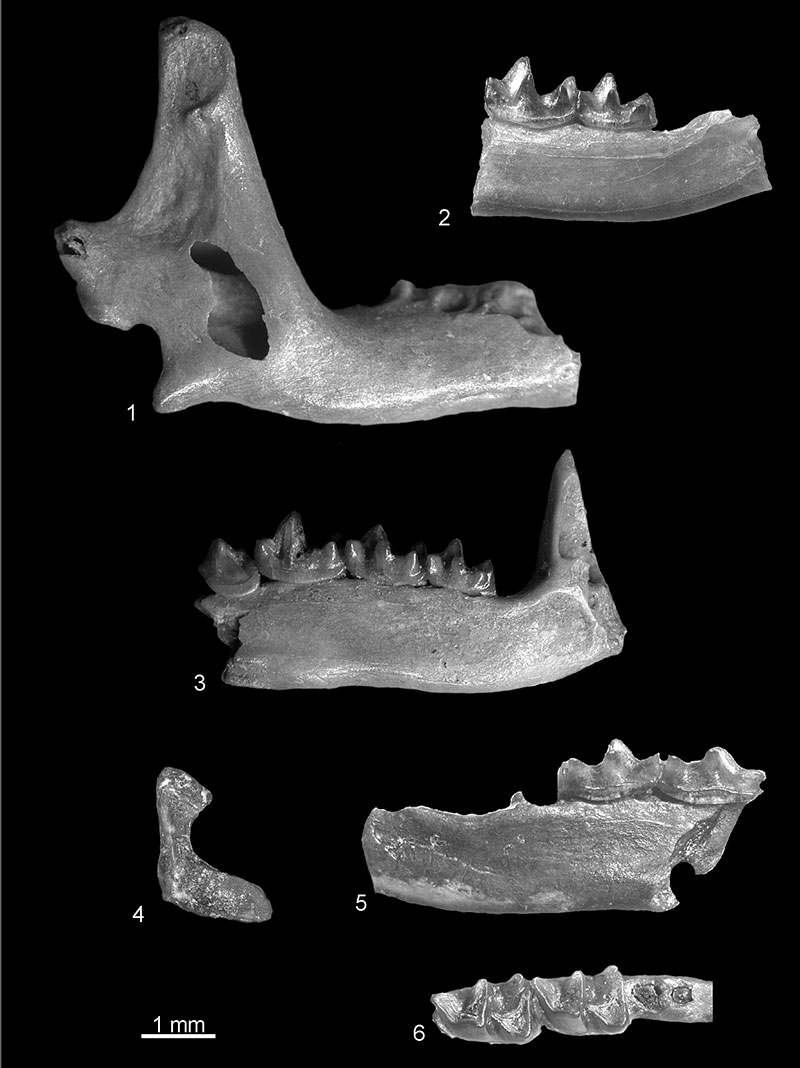
FIGURE 10. 1, Beremendia fissidens from Popovo 1, right I1, in anterior view (c.n. 29/I/12/1); 2, cf. Beremendia minor from Verkhnya Krynitsa 1, fragment of left mandible with proximal part of i1 in buccal view (c.n. 29/1/13/1); Petenyia dubia from Verkhnya Krynitsa 2 (3) and Lobkove (4): 3, left condyloid process, posterior view (c.n. 29/2/14/1); 4, left M1, occlusal view (c.n. 29/3/14/1). 5, Zelceina sp. from Verkhnya Krynitsa 2, fragment of left maxillae with M1-M2, occlusal view (c.n. 29/2/16/1).
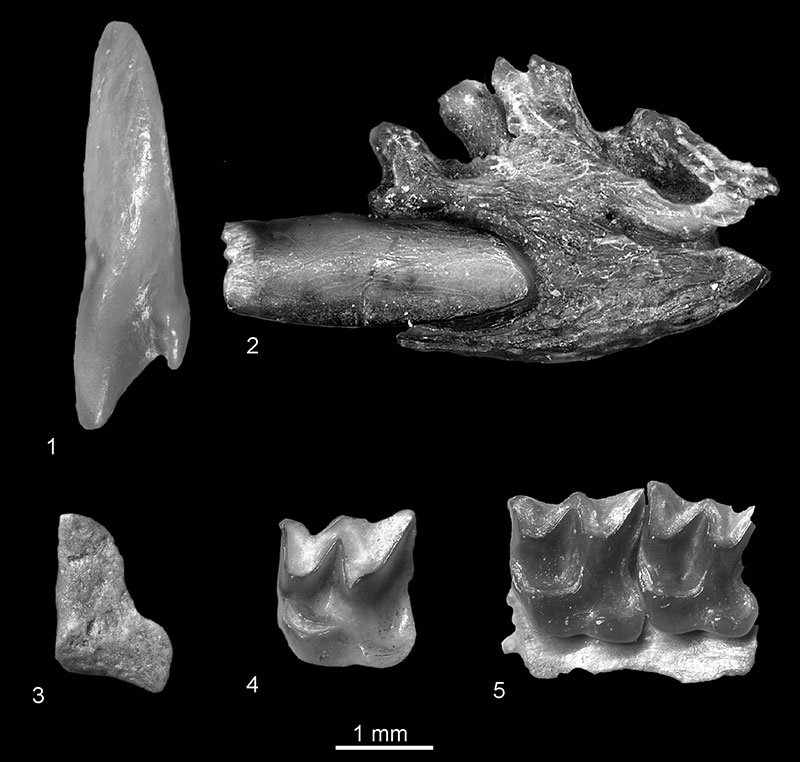
TABLE 1. Dimensions (in mm) of Schizogalerix species.
TABLE 2. Dimensions (in mm) of lower teeth of Mygalinia, Storchia and Ruemkelia (except of R. vinea and R. dekkersi ).
TABLE 3. Dimensions (in mm) of upper teeth of Desmana species (*specimen a little damaged; **calculated by author from minimum - maximum measurements; ***dimension made by author from picture, plate 1, 1, p. 109, Rümke, 1985).
TABLE 4. Dimensions (in mm) of lower teeth and humerus of Desmana species.
TABLE 5. Dimensions (in mm) of mandible and upper and lower teeth of Miosorex species.
TABLE 6. Dimensions (in mm) of mandible and lower teeth of Crusafontina.
TABLE 7. Dimensions (in mm) of mandible and lower teeth of Asoriculus species (*specimen a little damaged).
TABLE 8. Dimensions (in mm) of mandible and lower teeth of Neomysorex species.
TABLE 9. Dimensions (in mm) of mandible and lower teeth of Neomys newtoni.
TABLE 10. Dimensions (in mm) of mandible and upper and lower teeth of Beremendia species.
TABLE 11. Dimensions (in mm) of mandible and upper and lower teeth of Petenyiadubia.
TABLE 12. Dimensions (in mm) of upper teeth of Zelceina species.
TABLE 13. Number of insectivore species in the studied localities.
New data on Eulipotyphla (Insectivora, Mammalia)
from the Late Miocene to the Middle Pleistocene of Ukraine
Plain Language Abstract
This paper describes the remains of Erinaceidae, Talpidae and Soricidae (Eulipotyphla, Mammalia) from eight Ukrainian localities dated to the Late Miocene (Popovo 3, MN11, Verkhnya Krynitsa 2, MN11/MN12 and Lobkove MN12), the Late Pliocene (Verkhnya Krynitsa 1 and Popovo 2, MN16), the Pliocene/Pleistocene boundary (Popovo 1, MN16/MN17), the Early Pleistocene (Popovo 0, MN17) and the early Middle Pleistocene (Medzhybozh, Q3). Sixteen taxa found in the material include: one hedgehog (Schizogalerix sp.); four moles (cf. Mygalinia hungarica, Ruemkelia sp., Desmana sp., cf. Desmana sp.); and 11 shrews (Miosorex grivensis, cf. Miosorex sp., Crusafontina cf. kormosi, cf. Asoriculus sp., Neomysorex alpinoides, Neomys newtoni, Petenyia dubia, Petenyia hungarica, cf. Beremendia minor, Beremendia fissidens and Zelceina sp.). Their remains are fragmentary and the number of individuals and species in each locality is low with the exception of the Late Miocene Verkhnya Krynitsa 2 where 10 taxa of three families were found.
In the studied material the hedgehog is represented only by an extinct spineless Galericinae, the moles only by semiaquatic Desmaninae and shrews by old and primitive Crocidosoricinae and by red-toothed Soricinae. Among them the most interesting are the Pleistocene water shrew, Neomys newtoni, and Pliocene ?Beremendia minor, both species recorded for the first time in Ukrainian territory.
The paper gives measurements, illustrations, systematic positions and distributions of all studied taxa as well as the number of species found in each studied locality.
The authors emphasize that in comparison to other parts of Europe Ukrainian fossil insectivore mammals are still few in number and insufficiently studied.
Polish Abstrakt
Praca zawiera opis szczątków Erinaceidae, Talpidae i Soricidae (Eulipotyphla, Mammalia) z ośmiu kopalnych stanowisk Ukrainy. Podano dla nich krótką charakterystykę geologiczną oraz ich położenie i pozycję stratygraficzną. Trzy z nich (Popovo 3, MN11, Verkhnya Krynitsa 2, MN11/MN12 i Lobkove, MN12) datowane są na późny miocen, dwa (Verkhnya Krynitsa 1, początek MN16 i Popovo 2, koniec MN16) na późny pliocen, jedno (Popovo 1, MN16/MN17) na granicę pliocenu i plejstocenu, jedno (Popovo 0, MN17) na wczesny plejstocen i jedno (Medzhybozh, Q3) na wczesny środkowy plejstocen. Lista znalezionych ssaków owadożernych zawiera: Schizogalerix sp., cf. Mygalinia hungarica, Ruemkelia sp., Desmana sp., cf. Desmana sp., Miosorex grivensis, cf. Miosorex sp., Crusafontina cf. kormosi, cf. Asoriculus sp., Neomysorex alpinoides, Neomys newtoni, Petenyia dubia, Petenyia hungarica, cf. Beremendia minor, Beremendia fissidens i Zelceina sp. Poszczególne stanowiska różnią się zarówno liczbą szczątków jak i liczbą taksonów. Najbardziej obfita w szczątki i różnorodna w gatunki (10 taksonów z trzech rodzin) jest późnomioceńska fauna z Verkhnya Krynitsa 2. W pozostałych 7 stanowiskach znaleziono jedynie od jednego do trzech taksonów. W pracy zamieszczono pomiary i ilustracje wyżej wspomnianych taksonów, podano ich pozycję systematyczną, rozmieszczenie i liczbę taksonów w poszczególnych stanowiskach.
Ukrainian АБСТРАКТ
В статті описані залишки Erinaceidae, Talpidae і Soricidae (Eulipotyphla, Mammalia) відомі з восьми місцезнаходжень України. Три з них - з пізнього міоцену: Попово 3 (MN11), Верхня Криниця 2 (границя MN11/MN12) та Лобкове (MN12), два – з пліоцену: Верхня Криниця 1 (початок зони MN16) і Попово 2 (кінець зони MN16)), одне – з границі пліоцен-плейстоцен: Попово 1 (зони MN16/MN17), одне – з раннього плейстоцену: Попово 0 (MN17) і одне – з середнього плейстоцену: Меджибож (Q3). Представлена коротка характеристика геологічних умов (алювіальний тафономічний тип) місцезнаходжень, а також регіональне і стратиграфічне їх розміщення. Подається список видів комахоїдних: Schizogalerix sp., cf. Mygalinia hungarica, Ruemkelia sp., Desmana sp., cf. Desmana sp., Miosorex grivensis, cf. Miosorex sp., Crusafontina cf. kormosi, cf. Asoriculus sp., Neomysorex alpinoides, Neomys newtoni, Petenyia dubia, Petenyia hungarica, cf. Beremendia minor, Beremendia fissidens, Zelceina sp. Найбільше різноманіття (десять таксонів трьох родин) відмічено для Верхньої Криниці 2. В інших семи місцезнаходженнях представлено по одному, два, або три види. Представлені таблиці промірів та ілюстрації остеологічних решток, обговорені також систематичні позиції і поширення описаних видів.
Resumen en Español
Nuevos datos sobre Eulipotyphla (Insectivora, Mammalia) del Mioceno tardío al
Pleistoceno Medio de Ucrania
Se describen restos fósiles de Erinaceidae, Talpidae y Soricidae (Eulipotyphla, Mammalia) de ocho localidades: tres (Popovo 3, MN11, Verkhnya Krynitsa 2, MN11/MN12 y Lobkove, MN12) del Mioceno tardío, dos (Verkhnya Krynitsa 1, MN16 temprano y Popovo 2, MN16 tardío) del Plioceno, una (Popovo 1, MN16/MN17) del límite Plioceno/Pleistoceno, una (Popovo 0, MN17) del Pleistoceno temprano y, finalmente, otra (Medzhibozh, Q3) del Pleistoceno Medio temprano. Se describen brevemente las características geológicas, así como las posiciones locales y estratigráficas. La lista de especies insectívoras incluye: Schizogalerix sp., cf. Mygalinia hungarica, Ruemkelia sp., Desmana sp., cf. Desmana sp., Miosorex grivensis, cf. Miosorex sp., Crusafontina cf. kormosi, cf. Asoriculus sp., Neomysorex alpinoides, Neomys newtoni, Petenyia dubia, Petenyia hungarica, cf. Beremendia minor, Beremendia fissidens y Zelceina sp. Tanto la abundancia de restos de insectívoros como su diversidad taxonómica varían entre las localidades. La asociación fósil más numerosa y diversa (10 taxones en tres familias) es la del Mioceno tardío de Verkhnya Krynitsa 2. Las siete localidades restantes produjeron entre una y tres especies. Los restos fósiles se describen, se miden y se ilustran, y también se discute sus posiciones y distribuciones sistemáticas.
Palabras clave: mamíferos insectívoros; Neógeno; Cuaternario; Europa Oriental
Traducción: Enrique Peñalver (Sociedad Española de Paleontología)
Résumé en Français
Nouvelles données sur les Eulipotyphla (Insectivora, Mammalia) d'Ukraine du Miocène récent au Pléistocène moyen
Les restes d'Erinaceidae, Talpidae et Soricidae (Eulipotyphla, Mammalia) de huit localités sont décrits : trois du Miocène récent (Popovo 3, MN11, Verkhnya Krynitsa 2, MN11/MN12, et Lobkove, MN12), deux du Pliocène (Verkhnya Krynitsa 1, début de MN16 et Popovo 2, fin de MN16), une de la limite Pliocène-Pléistocène (Popovo 1, MN16/MN17), une du Pléistocène ancien (Popovo 0, MN17) et une du Pléistocène moyen ancien (Medzhybozh, Q3). Les caractéristiques géologiques ainsi que les positions locales et stratigraphiques sont brièvement décrites. La liste des espèces d'insectivores inclut : Schizogalerix sp., cf. Mygalinia hungarica, Ruemkelia sp., Desmana sp., cf. Desmana sp., Miosorex grivensis, cf. Miosorex sp., Crusafontina cf. kormosi, cf. Asoriculus sp., Neomysorex alpinoides, Neomys newtoni, Petenyia dubia, Petenyia hungarica, cf. Beremendia minor, Beremendia fissidens et Zelceina sp. L'abondance des restes d'insectivores et leur diversité taxinomique varient selon les localités. L'assemblage présentant le plus de restes et le plus de diversité taxinomique (10 taxons de trois familles) est celui de la localité Verkhnya Krynitsa 2 du Miocène récent. Les sept autres localités ont livré de une à trois espèces. Les restes sont décrits, mesurés et illustrés, et leurs positions systématiques et leurs distributions sont discutées.
Mots-clés : mammifères insectivores ; Néogène ; Quaternaire ; Europe orientale
Translator: Antoine Souron
Deutsche Zusammenfassung
Neue Informationen zu Eulipotyphla (Insectivora, Mammalia) aus dem späten Miozän bis mittleren Pleistozän der Ukraine
Es werden Überreste von Erinaceidae, Talpidae und Soricidae (Eulipotyphla, Mammalia) aus acht Fundstellen beschrieben: drei (Popovo 3, MN11, Verkhnya Krynitsa 2, MN11/MN12 und Lobkove, MN12) aus dem späten Miozän, zwei (Verkhnya Krynitsa 1, frühes MN16 und Popovo 2, spätesMN16) aus dem Pliozän, eine (Popovo 1, MN16/MN17) von der Pliozän/Pleistözän Grenze, eine (Popovo 0, MN17) aus dem frühen Pleistozän und eine (Medzhybozh, Q3) aus dem frühen Mittleren Pleistozän. Kurze geologische Kenndaten sowie lokale und stratigraphische Positionen werden beschrieben. Die Liste der insectivoren Arten beinhaltet: Schizogalerix sp., cf. Mygalinia hungarica, Ruemkelia sp., Desmana sp., cf. Desmana sp., Miosorex grivensis, cf. Miosorex sp., Crusafontina cf. kormosi, cf. Asoriculus sp., Neomysorex alpinoides, Neomys newtoni, Petenyia dubia, Petenyia hungarica, cf. Beremendia minor, Beremendia fissidens, Zelceina sp. Sowohl die Häufigkeit der Überreste als auch deren taxonomische Diversität ist zwischen den Fundstellen unterschiedlich. Die spätmiozäne Verkhnya Krynitsa 2 weist die zahlreichste und diverseste Assemblage auf (10 Taxa von drei Familien). Die übrigen sieben Fundstellen lieferten eine bis drei Arten. Die Überreste werden beschrieben, vermessen und illustriert und ihre systematische Positionen und Verbreitungen werden diskutiert.
Schlüsselwörter: Insektivore Säugetiere; Neogen; Quartär; Osteuropa
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024