

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
 Linda Frey. Palaeontological Institute and Museum, University of Zurich, Karl-Schmid-Str. 4, CH-8006, Switzerland. linda.frey@pim.uzh.ch
Linda Frey. Palaeontological Institute and Museum, University of Zurich, Karl-Schmid-Str. 4, CH-8006, Switzerland. linda.frey@pim.uzh.ch
Linda Frey is a doctoral student at the Paläontologisches Institut und Museum in Zürich, Switzerland. She is interested in diversity changes and palaeoecology and she mainly works on Devonian invertebrates and fishes, currently focusing on chondrichthyans.
She received a MSc of Science in Biology with a specialisation in Palaeontology from the University of Zürich in 2013.

 Erin E. Maxwell. Staatliches Museum für Naturkunde, Rosenstein 1, DE-70191 Stuttgart, Germany. erin.maxwell@smns-bw.de
Erin E. Maxwell. Staatliches Museum für Naturkunde, Rosenstein 1, DE-70191 Stuttgart, Germany. erin.maxwell@smns-bw.de
Erin Maxwell is a vertebrate paleontologist at the Staatliches Museum für Naturkunde in Stuttgart, Germany. Her research focuses on understanding body plan evolution and ecomorphology in Mesozoic fishes and marine reptiles.
She received a BSc in Evolutionary Biology from the University of Alberta, and a PhD from McGill University in 2008.
 Marcelo R. Sánchez-Villagra. Palaeontological Institute and Museum, University of Zurich, Karl-Schmid-Str. 4, CH-8006, Switzerland. m.sanchez@pim.uzh.ch (corresponding author)
Marcelo R. Sánchez-Villagra. Palaeontological Institute and Museum, University of Zurich, Karl-Schmid-Str. 4, CH-8006, Switzerland. m.sanchez@pim.uzh.ch (corresponding author)
Marcelo R. Sánchez-Villagra is Associate Professor of Palaeobiology at the University of Zürich. A native from Buenos Aires, he studied Biology in Caracas and obtained a PhD at Duke University and the Habilitation in Zoology from the University of Tübingen. After being Researcher at the Natural History Museum in London, he moved to Zürich and started the 'Evolutionary Morphology and Palaeobiology of Vertebrates' research group. He published Embryos in Deep Time (2012), which deals with much of his past research interest, the development and palaeobiology of vertebrate animals. He regularly leads collaborative field palaeontological work in Venezuela and he is currently working on the evo-devo of mammalian domestication.
APPENDIX
Meristic and morphometric data collected for this study.
TABLE 1. Examples of previous studies on phenotypic variation in populations of fossil fishes.
TABLE 2. Mean ± standard deviation and range (in parentheses) of morphometric and meristic characters for specimens of Prolebias rhenanus, Pr. stenoura and Paralebias cephalotes.
TABLE 3. Coefficient of variation (COV) for morphometric and meristic characters.
TABLE 4. Correlation matrix (correlation coefficient below the diagonal; p-values above the diagonal) for fin vs. vertebral and meristic vs. morphometric characters of Prolebias rhenanus. Significant correlations (α = 0.05) are highlighted in bold face fonts. Abbreviations for morphometric characters: SL, standard length; TVL, total length of vertebral column; PCL and CL, length of precaudal and caudal region of vertebral column; DPL and APL, length of dorsal and anal fin bases. Abbreviations for meristic characters: TV, total number of vertebrae; PV and CV, number of precaudal and caudal vertebrae; DP and AP, number of dorsal and anal pterygiophores.
TABLE 5. Correlation matrix (correlation coefficient below the diagonal; p-values above the diagonal) for fin vs. vertebral and meristic vs. morphometric characters of Prolebias stenoura. Significant correlations (α = 0.05) are highlighted in bold face fonts. Abbreviations are the same as in Table 4.
TABLE 6. Correlation matrix (correlation coefficient below the diagonal; p-values above the diagonal) for fin vs. vertebral and meristic vs. morphometric characters of Paralebias cephalotes. Significant correlations (α = 0.05) are highlighted in bold face fonts. Abbreviations are the same as in Table 4.
FIGURE 1. Photograph of a plate with Paralebias cephalotes (P.36131, NHM, collected from Aix-en-Provence, France). Scale bar equals 6 cm.

FIGURE 2. Photographs of fossil killifish species examined in this study. 1, Prolebias rhenanus (Ru 99, Naturhistorisches Museum Basel); 2, Pr. stenoura (28491 n, NHM); 3, Paralebias cephalotes (P.1831a, NHM). Scale bars equal 1 cm.

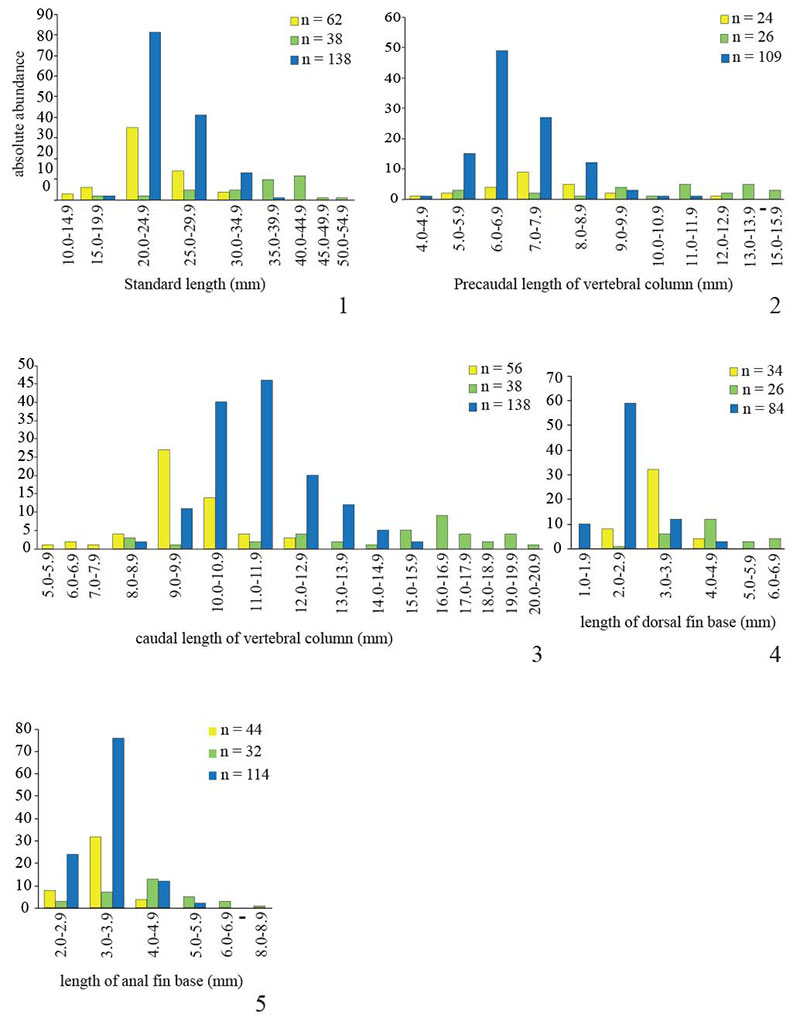
FIGURE 3. Bar charts showing frequency distribution of meristic characters for the three fossil Killifish species Prolebias rhenanus (yellow), Pr. stenoura (green) and Paralebias cephalotes (blue). 1, number of vertebrae; 2, number of precaudal vertebrae; 3, number of caudal vertebrae; 4, number of dorsal pterygiophores; 5, number of anal pterygiophores.
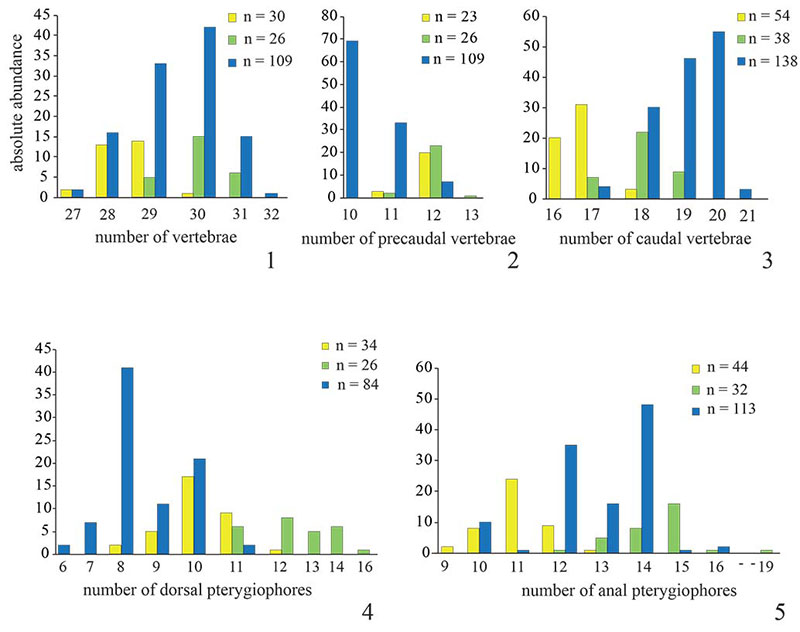
FIGURE 4. Bar charts showing frequency distribution of morphometric data (in mm) for the three fossil Killifish species Prolebias rhenanus (yellow), Pr. stenoura (green) and Paralebias cephalotes (blue). 1, standard length; 2, precaudal length of vertebral column; 3, caudal length of vertebral column; 4, length of dorsal fin base; 5, length of anal fin base.
Intraspecific variation in fossil vertebrate populations: Fossil killifishes (Actinopterygii: Cyprinodontiformes) from the Oligocene of Central Europe
Plain Language Abstract
The study of variation of fossil populations is very important to establish the boundaries among species, and once this has been done, to understand the evolution of variability and the relations among parts of the organisms and their integration. Killifishes from Central Europe are a good subject of study given their samples from different localities and habitats. We quantified variation in discrete characters of the vertebral column and the fins, as well as in several body measurements, in three species. We found similar degrees of variation among them, and vertebral counts were unexpectedly rather invariant. Some patterns do exist: e.g., in Prolebias spp. The caudal vertebral counts are significantly more variable than precaudal counts. Traits pertaining to the dorsal and anal fins are extremely variable, more so than vertebral ones in general, possibly because fins are more peripherally positioned and form later during development. We found short-term conservation in patterns of variation within closely related species and longer-term divergence of trait variability between 'families' Poecillidae versus Valenciidae.
Resumen en Español
Variaciones intraespecíficas en poblaciones de vertebrados fósiles: peces killi fósiles (Actinopterygii: Cyprinodontiformes) del Oligoceno de Europa Central
Una cuantificación de la variación intraespecífica de características morfológicas es necesaria no solamente como base de un trabajo taxonómico pero también para entender un aspecto básico de la evolución. Las comparaciones entre especies podrían revelar diferencias en la plasticidad del desarrollo entre ellas e incluso brindar claves sobre evolucionabilidad. Estudios de este tipo en especies extintas son escasos debido a la falta de muestras adecuadas, pero existen algunas excepciones notables. Los datos morfométricos y merísticos fueron recolectados para tres especies de peces killi fósiles con el objetivo de cuantificar la variación intraespecífica: 67 especímenes de Prolebias rhenanus del Oligoceno Inferior de Baden-Württemberg, 40 especímenes de Pr. stenoura del Oligoceno Inferior de Puy-de-Dôme, y 141 especímenes de Paralebias cephalotes del Oligoceno Superior de Aix-en-Provence. Patrones casi idénticos de variación intraespecífica existen entre las dos especies de Prolebias. Las longitudes de las bases de las aletas y las longitudes precaudales de la columna vertebral son las características más variables en ambas especies, seguidas por el conteo de pterigióforos, luego por otras medidas de longitud axiales y, menos variable, el conteo vertebral. En ninguna especie se encontró una diferencia significativa en la variación observada entre las características de aletas y vértebras o características morfométricas y merísticas. La única correlación significativa compartida por las dos especies es la relación positiva entre la longitud caudal de la columna vertebral y la longitud de la base de la aleta anal. La longitud precaudal de la columna vertebral es menos variable en Paralebias que en Prolebias, mientras que el conteo vertebral precaudal es más variable, en valores absolutos del coeficiente de variación. La variabilidad en la elongación de las vértebras precaudales puede ser la causa subyacente de la ausencia de correlación entre la reducción de variación en la longitud precaudal de la columna vertebral y el aumento de variación en el conteo precaudal en Paralebias cephalotes. Registramos patrones más similares de variación entre congéneres de Prolebias que entre Prolebias y Paralebias, sugiriendo que la variabilidad de características cambia a lo largo del tiempo evolutivo en este linaje de actinopterigios.
Palabras clave: Atherinomorpha; morfología; variabilidad; taxonomía; esqueleto; modularidad
Traducción: Diana Elizabeth Fernández
Résumé en Français
Variation intraspécifique dans des populations de vertébrés fossiles : killis (Actinopterygii : Cyprinodontiformes) fossiles de l'Oligocène d'Europe centrale
Une quantification de la variation intraspécifique des traits morphologiques est nécessaire non seulement comme base du travail de taxinomie, mais aussi pour comprendre un aspect basique de l'évolution. Des comparaisons entre espèces pourraient révéler des différences de plasticité de leur développement, et même donner des indices sur leur évoluabilité. Les études de ce genre sont rares pour les espèces de vertébrés éteintes en raison d'un manque d'échantillons adéquats, mais il existe des exceptions notables. Des données morphométriques et méristiques ont été collectées pour trois espèces de killis fossiles afin de quantifier la variation intraspécifique : 67 spécimens de Prolebias rhenanus de l'Oligocène inférieur de Baden-Württemberg, 40 spécimens de Pr. stenoura de l'Oligocène inférieur du Puy-de-Dôme, et 141 spécimens de Paralebias cephalotes de l'Oligocène supérieur d'Aix-en-Provence. Les deux espèces de Prolebias présentent des schémas de variation intraspécifique presque identiques. Les longueurs de la base des nageoires et les longueurs de la partie précaudale de la colonne vertébrale sont les traits les plus variables chez les deux espèces, suivis du nombre de ptérygiophores, puis des autres mesures de longueur axiale. Le nombre de vertèbres est le trait le moins variable. Aucune des espèces ne montre de différence significative dans la variation observée entre les traits des nageoires et des vertèbres d'une part, et entre les traits méristiques et morphométriques d'autre part. La seule corrélation significative partagée entre les deux espèces est la relation positive entre la longueur de la partie caudale de la colonne vertébrale et la longueur de la base de la nageoire anale. La longueur de la partie précaudale de la colonne vertébrale est moins variable chez Paralebias que chez Prolebias, alors que le nombre de vertèbres précaudales est plus variable en termes de valeurs absolues des coefficients de variation. La variabilité de l'élongation des vertèbres précaudales pourrait être la cause sous-jacente de l'absence de corrélation entre la variation plus faible de la longueur de la partie précaudale de la colonne vertébrale et la variation plus forte du nombre de vertèbres précaudales chez Paralebias cephalotes. Nous signalons des schémas de variation plus similaires entre les espèces du genre Prolebias qu'entre les genres Prolebias et Paralebias, ce qui suggère que la variabilité des traits change au cours de l'évolution dans cette lignée d'actinoptérygien.
Mots-clés : Atherinomorpha ; morphologie ; variabilité ; taxinomie ; squelette ; modularité
Translator: Antoine Souron
Deutsche Zusammenfassung
Intraspezifische Variation in fossilen Wirbeltier-Populationen: Fossile Zahnkärpflinge (Actinopterygii: Cyprinodontiformes) aus dem Oligozän von Zentraleuropa.
Die Quantifizierung intraspezifischer Variation von morphologischen Merkmalen ist nicht nur für die taxonomische Arbeit notwendig sondern auch um die grundlegenden Aspekte der Evolution zu verstehen. Innerartliche Vergleiche können Unterschiede in der Plastizität der Entwicklung enthüllen und sogar Hinweise auf deren Evolvierbarkeit geben. Untersuchungen dieser Art sind aufgrund der ungenügenden Stichprobengrössen bei ausgestorbenen Wirbeltierarten selten, allerdings gibt es beachtenswerte Ausnahmen. Morphometrische und meristische Daten von drei fossilen Zahnkärpflings-Arten wurden gesammelt um die intraspezifische Variation zu quantifizieren: 67 Exemplare von Prolebias rhenanus aus dem Unteren Oligozän von Baden-Württemberg, 40 Exemplare von Pr. stenoura aus dem Unteren Oligozän von Puy-de-Dôme, und 141 Exemplare von Paralebias cephalotes aus dem Oberen Oligozän von Aix-en-Provence. Die beiden Prolebias-Arten weisen fast identische Muster in intraspezifischer Variation auf. In den zwei Arten sind die Flossenbasis-Längen und die präcaudalen Längen der Wirbelsäule die variabelsten Merkmale, gefolgt von der Anzahl Pterygiophoren, anderen axialen Längenmesswerten und von dem am wenigsten variablen Merkmal der Anzahl Wirbel. In keiner der beiden Arten gab es einen signifikanten Unterschied in der Variation zwischen Flossen- und Wirbelmerkmalen beziehungsweise zwischen meristischen und morphologischen Merkmalen. Die einzige vorhandene signifikante Korrelation ist die positive Beziehung zwischen der caudalen Länge der Wirbelsäule und der Länge der analen Flossenbasis in den zwei Prolebias-Arten. Die präcaudale Länge der Wirbelsäule ist in Paralebias weniger variable als in Prolebias, während die absoluten Zahlen des Variationskoeffizienten von der Anzahl präcaudaler Wirbel in Paralebias variabler sind. Der Abwesenheit der Korrelation zwischen reduzierter Variation in der precaudalen Länge der Wirbelsäule und erhöhter Variation in Anzahl präcaudaler Wirbel in Paralebias cephalotes mag die Variabilität der Verlängerung der präcaudalen Wirbel zu Grunde liegen. Wir berichten hier über ähnlichere Variationsmuster innerhalb von Prolebias-Artverwandten als zwischen Prolebias und Paralebias und leiten daraus ab, dass sich die Merkmalsvariabilität in dieser Actinopterygier-Linie über evolutionäre Zeit hinweg verändert hat.
Schlüsselwörter: Atherinomorpha; Morphologie; Variabilität; Taxonomie; Skelett; Modularität
Translator: Author
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024