

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
 Jorge D. Carrillo-Briceño. Paleontological Institute and Museum, University of Zurich, Karl-Schmid-Strasse 4, Zürich, 8006, Switzerland. jorge.carrillo@pim.uzh.ch
Jorge D. Carrillo-Briceño. Paleontological Institute and Museum, University of Zurich, Karl-Schmid-Strasse 4, Zürich, 8006, Switzerland. jorge.carrillo@pim.uzh.ch
Jorge is Geographer and Magister in Oceanography with ample field and lab experience in diverse South American countries. Currently he is a doctoral student in the Palaeontological Institute and Museum of the University of Zurich. His research interest is the diversity and paleobiology of Mesozoic and Cenozoic Chondrichthyans from Tropical America.

 Orangel A. Aguilera. Universidade Federal Fluminense, Instituto de Biologia, Departamento de Biologia Marinha, Programa de Pós-graduação em Biología Marinha. Campus do Valonguinho, Outeiro São João Batista, s/nº. 24020-141, Niterói, RJ, Brasil. orangel.aguilera@gmail.com
Orangel A. Aguilera. Universidade Federal Fluminense, Instituto de Biologia, Departamento de Biologia Marinha, Programa de Pós-graduação em Biología Marinha. Campus do Valonguinho, Outeiro São João Batista, s/nº. 24020-141, Niterói, RJ, Brasil. orangel.aguilera@gmail.com
Orangel Aguilera is Professor of the postgraduate program in Marine Biology of the Universidade Federal Fluminense, Brazil. He received his first degree in Oceanography at the Fundação Universidade de Rio Grande in Brasil, his Masters in Marine Biology at the Venezuelan Oceanographic Institute of the Universidad de Oriente, and his PhD from the Central University of Venezuela. He was a post-doctoral fellow at the Smithsonian Tropical Research Institute. His research spans alpha taxonomy - describing new living and fossil species, paleoecology, and macroecology, being interested in large scale trends in species diversity and community evolution. Many of his investigations focus on a group of freshwater and marine fishes (recent and fossil), reptiles and mammals (Neogene species) in Tropical America.
 Carlos De Gracia. Institute of Geology and Paleontology, Faculty of Science, Charles University in Prague, Albertov 6, 128 43 - Prague 2, Czech Republic. degraciac@gmail.com
Carlos De Gracia. Institute of Geology and Paleontology, Faculty of Science, Charles University in Prague, Albertov 6, 128 43 - Prague 2, Czech Republic. degraciac@gmail.com
Carlos De Gracia is biologist with expertise in marine paleontology and has experience working on field and laboratory in Panama, Costa Rica, Colombia and Venezuela. Currently is a Master student in Geobiology in the Institute of Geology and Paleontology of Charles University in Prague. His research interest is the paleoecology and paleobiology of Cenozoic bony fishes in tropical America.
 Gabriel Aguirre-Fernández. Paleontological Institute and Museum, University of Zurich, Karl-Schmid-Strasse 4, Zürich, 8006, Switzerland. gabriel.aguirre@pim.uzh.ch
Gabriel Aguirre-Fernández. Paleontological Institute and Museum, University of Zurich, Karl-Schmid-Strasse 4, Zürich, 8006, Switzerland. gabriel.aguirre@pim.uzh.ch
Gabriel is a marine biologist with a PhD in Geology. He currently works as a postdoctoral researcher at the University of Zurich. His research focuses on the systematics of fossil whales and dolphins.
 René Kindlimann. Paleontological Institute and Museum, University of Zurich, Karl-Schmid-Strasse 4, Zürich, 8006, Switzerland. mad_design@gmx.ch
René Kindlimann. Paleontological Institute and Museum, University of Zurich, Karl-Schmid-Strasse 4, Zürich, 8006, Switzerland. mad_design@gmx.ch
René Kindlimann is a private paleontologist researching and collecting chondrichthyan remains for more than 45 Years. He's focusing on everything that matters with extinct and extant chondrichthyans (sharks, rays and chimaeras). He published several papers and he is also co-authoring in other "shark" papers.
 Marcelo R. Sánchez-Villagra. Paleontological Institute and Museum, University of Zurich, Karl-Schmid-Strasse 4, Zürich, 8006, Switzerland. m.sanchez@pim.uzh.ch
Marcelo R. Sánchez-Villagra. Paleontological Institute and Museum, University of Zurich, Karl-Schmid-Strasse 4, Zürich, 8006, Switzerland. m.sanchez@pim.uzh.ch
Marcelo Sánchez is Associate Professor of Palaeontology at the University of Zürich and head of the lab of evolutionary morphology and palaeobiology of vertebrates at that institution.
FIGURE 1. Location and stratigraphy of the Cantaure Formation. 1. Fossiliferous localities. 2. Stratigraphic section of locality 1 (artesian well) and locality 2 (outcrop). Stratigraphic column modified after Aguilera (2010) and Aguilera et al. (2013).
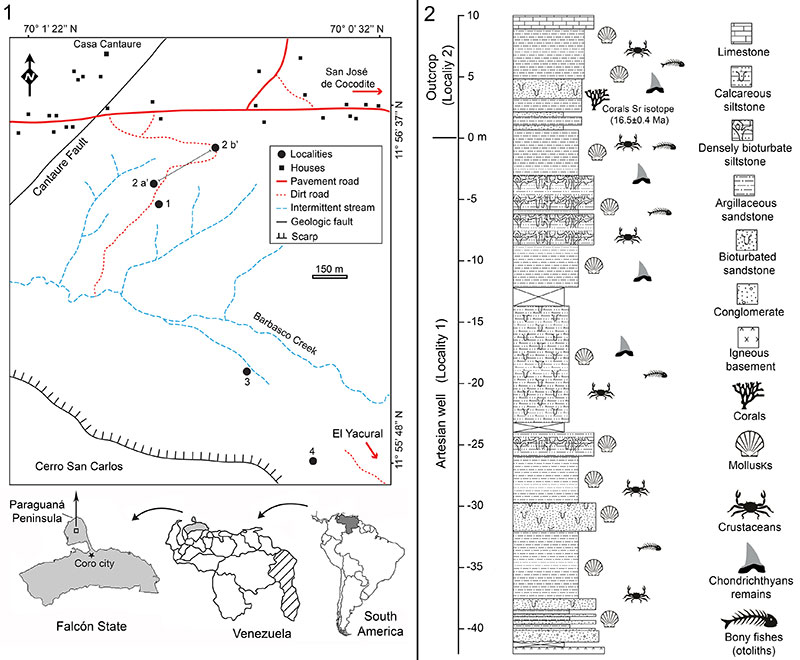
FIGURE 2. Elasmobranch paleodiversity of the Cantaure Formation. 1. Orders and 2. Families and genera.
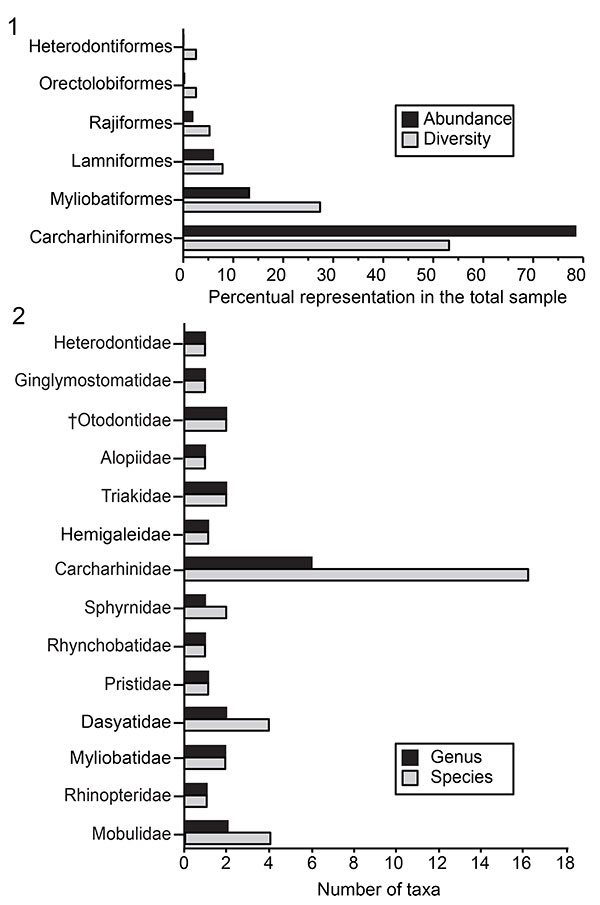
FIGURE 3. Orectolobiformes and Lamniformes of the Cantaure Formation. 1-9. Nebrius sp. (AMU-CURS-986); 10-11. † Paratodus benedenii (AMU-CURS-645). 12-21. † Carcharocles megalodon (12: AMU-CURS-997; 13: AMU-CURS-512; 14: UNEFM-PF-349; 15: AMU-CURS-996; 16: AMU-CURS-518; 17-18: UNEFM-PF-s/n; 19: AMU-CURS-512 and 20-21: AMU-CURS-515). 22-26. Alopias cf. A. vulpinus (22-23: AMU-CURS-985; 24-25: AMU-CURS-983 and 26: AMU-CURS-984). Jaw position: upper (10-11? 12, 14? 17, 22-25), lower (13, 15, 18) and indet. (1-9, 16, 19-21, 26). View: labial (1, 6, 10, 13, 20-21, 23, 24), lingual (2, 7, 11-12, 14-19, 22, 25-26), profile (3, 8), occlusal (4) and basal (5, 9).
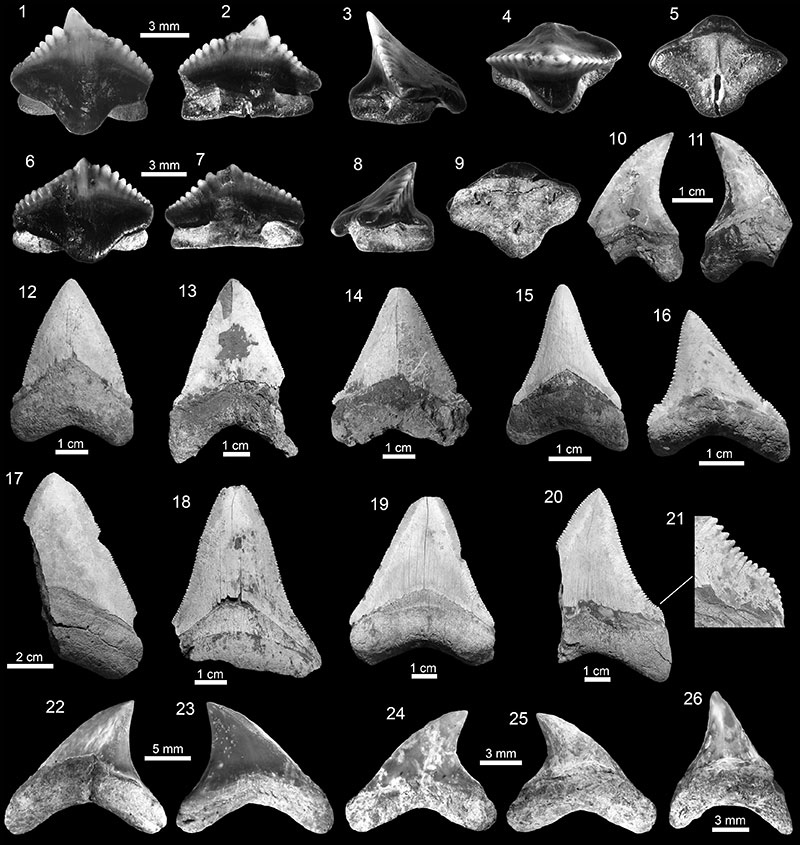
FIGURE 4. Carcharhiniformes of the Cantaure Formation. 1-2. Galeorhinus cf. G. galeus (AMU-CURS-974). 3-7. Mustelus sp. (AMU-CURS-975). 8-13. † Hemipristis serra (8: AMU-CURS-731 and 9-13: AMU-CURS-644). 14-18. † Galeocerdo aduncus (14-15: AMU-CURS-730 and 16-18: AMU-CURS-647). 19-23. † Galeocerdo mayumbensis (19-20: AMU-CURS-995 and 21-23: AMU-CURS-646). Jaw position: upper (9-12), lower (8? 13) and indet. (1-7, 14-23). View: labial (1, 6, 7, 10, 15, 19, 21-22), lingual (2, 4, 8-9, 11-14, 16-18, 20, 23) and occlusal-lingual (3, 5).
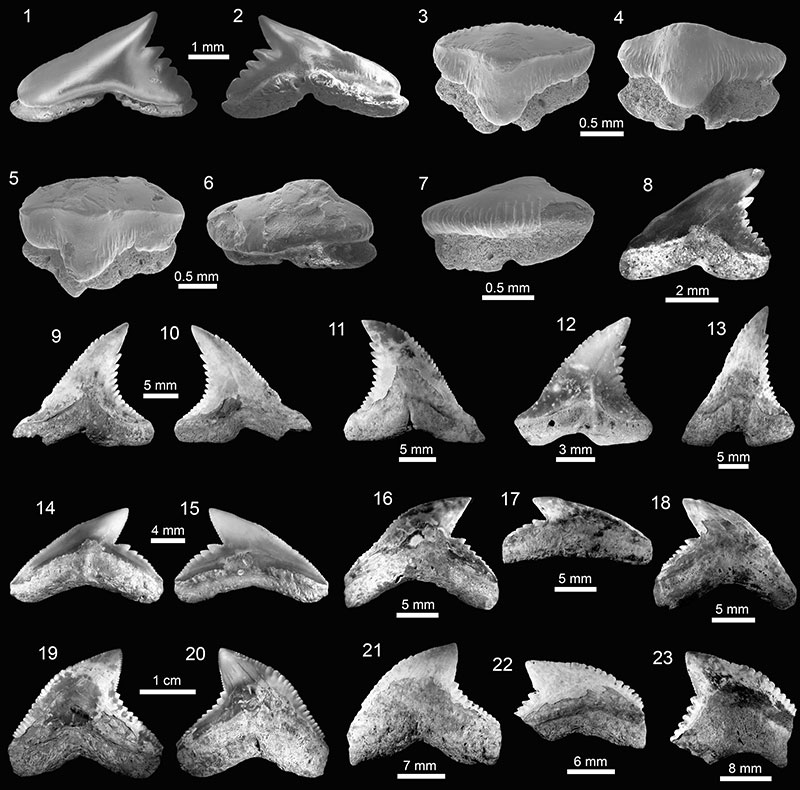
FIGURE 5. Carcharhiniformes of the Cantaure Formation. 1-5. Rhizoprionodon sp. (1-3, 5: AMU-CURS-975 and 4: AMU-CURS-960). 6-10. † Carcharhinus ackermannii (6-7: AMU-CURS-713 and 8-10: AMU-CURS-714). 11-15. Carcharhinus brachyurus (AMU-CURS-990). 16-20. Carcharhinus falciformis (AMU-CURS-991). 21-25. Carcharhinus cf. C. galapagensis (AMU-CURS-973). Jaw position: upper (2, 4-14, 16-25) and lower (1, 3. 15). View: labial (6, 9, 12-13, 17-18, 20-21, 24) and lingual (1-5, 7-8, 10-11, 14-16, 19, 22-23, 25).

FIGURE 6. Carcharhiniformes of the Cantaure Formation. 1-4. †Carcharhinus gibbesii (AMU-CURS-958). 5-8. Carcharhinus cf. C. limbatus (AMU-CURS-944). 9-12. Carcharhinus cf. C. macloti (AMU-CURS-957). 13-17. Carcharhinus perezi (AMU-CURS-956). 18-22. Carcharhinus sp.1 (AMU-CURS-943). 23-24. Carcharhinus spp. (23: AMU-CURS-941 and 24: AMU-CURS-940). Jaw position: upper (1-24). View: labial (1, 4, 6-7, 10, 12, 14, 17-18, 20, 22-24) and lingual (2-3, 5, 8-9, 11, 13, 15-16, 19, 21).
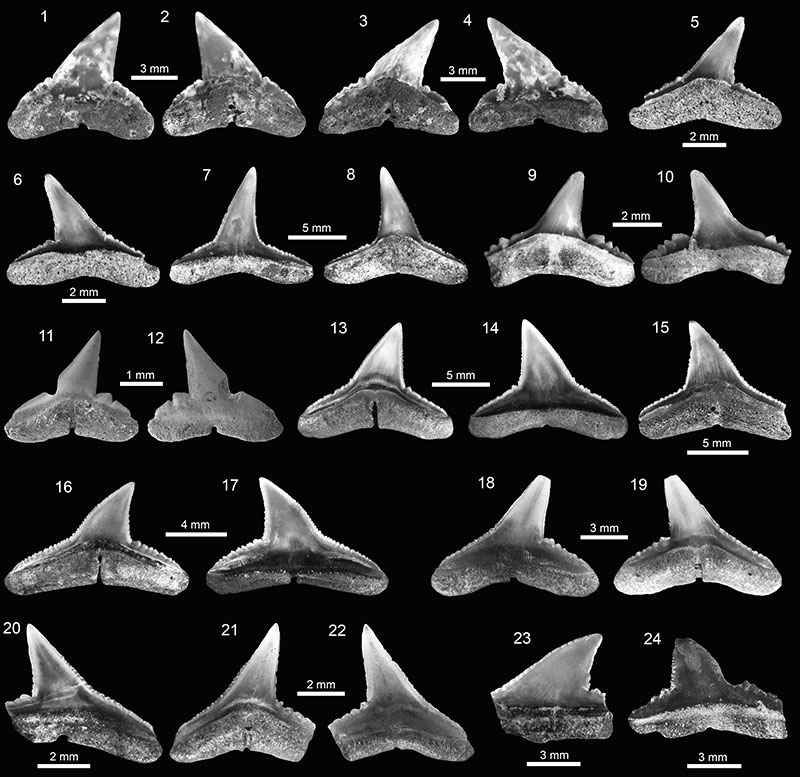
FIGURE 7. Carcharhiniformes of the Cantaure Formation. 1-5. † Isogomphodon acuarius (1-2: AMU-CURS-950 and 3-5: AMU-CURS-951). 6-10. † Negaprion eurybathrodon (6-9: AMU-CURS-989 and 10: AMU-CURS-987). 11-15. † Physogaleus contortus (11-12: AMU-CURS-719 and 13-15: AMU-CURS-648). 16-20. Carcharhinidae indet. (AMU-CURS-939). 21-25. † Sphyrna arambourgi (21-24: AMU-CURS-961 and 25: AMU-CURS-962). Jaw position: upper (1-3, 6-9, 16-19? 21-22, 25), lower (4-5, 10, 20? 23-24) and indet. (11-15). View: labial (2, 6, 9, 11, 13, 17-18, 22, 24-25) and lingual (1, 3-5, 7-8, 10, 12, 14-16, 19-21, 23).
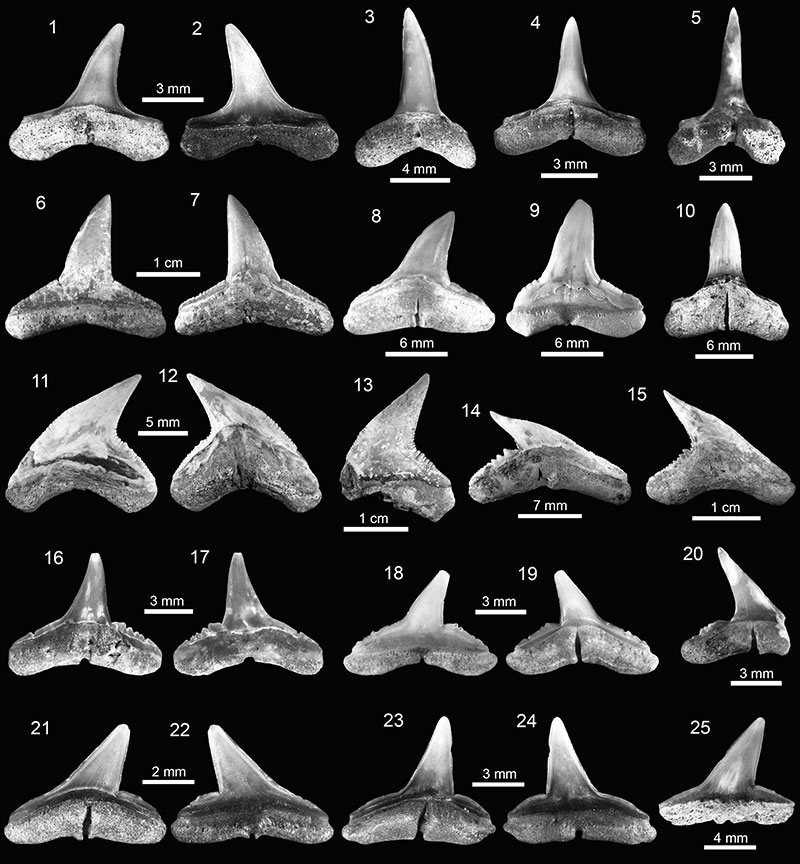
FIGURE 8. Carcharhiniformes and Rajiformes of the Cantaure Formation. 1-5. Sphyrna cf. † S. laevissima (1-3, 5: AMU-CURS-964 and 4: AMU-CURS-963). 6-11. Rhynchobatus sp. (6, 9: AMU-CURS-968; 7-8, 10: AMU-CURS-969 and 11: AMU-CURS-970). 12-20. Pristis sp. (rostral denticle 12-14: AMU-CURS-242; buccal teeth 15-20: AMU-CURS-967). Jaw position: upper (1-2) and indet. (3-11, 15-20). View: labial (1, 3), lingual (2, 4-5, 11), occlusal (6-8, 15, 17, 19), profile (18, 20), basal (9-10, 16), posterior (12) and dorsal (13-14).
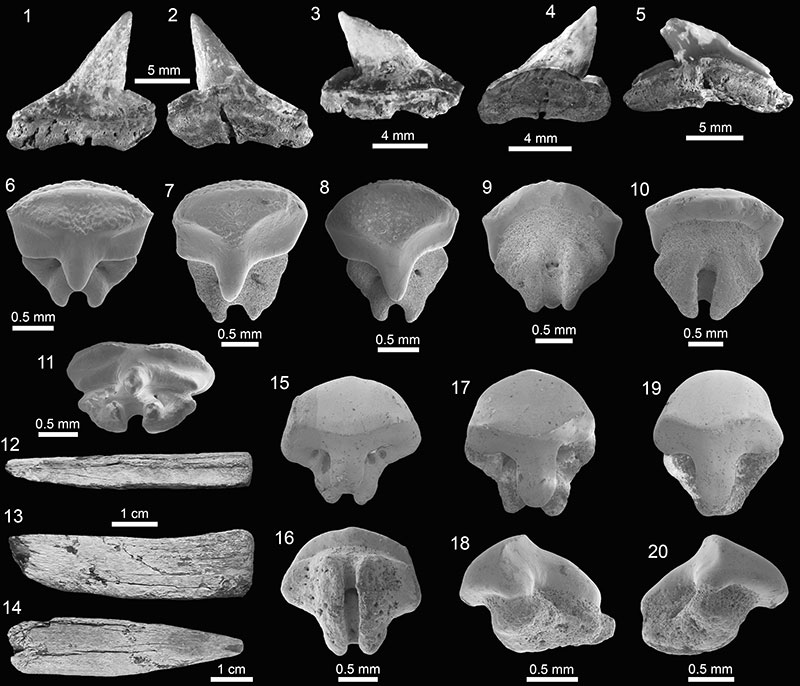
FIGURE 9. Myliobatiformes of the Cantaure Formation. 1-10. Dasyatis sp. (1-2, 7-8: AMU-CURS-930; 3-4: AMU-CURS-932; 5-6: AMU-CURS-933 and 9-10: AMU-CURS-931). 11-17. cf. Pteroplatytrygon sp. (AMU-CURS-926). 18-21. cf. Taeniurops sp. (AMU-CURS-925). 22-27. Dasyatidae indet. (22-24: AMU-CURS-936 and 25-27: AMU-CURS-937). Jaw position: indet. (1-27). View: labial (4, 9, 11, 15, 21), lingual (2, 16), occlusal (1, 5, 13, 18, 22, 25), profile (3, 6-7, 10, 12, 17, 19, 24, 27) and basal (8, 14, 20, 23, 26).

FIGURE 10. Myliobatiformes of the Cantaure Formation. 1-3. Aetobatus sp. (1-3: AMU-CURS-716). 4-14. Aetomylaeus sp. (AMU-CURS-750). 15-29. Rhinoptera sp. (15-18: AMU-CURS-979; 19-21, 22-23, 24-25, 28-29: AMU-CURS-977 and 26-27: AMU-CURS-981). Jaw position: indet. (1-29). View: labial (11, 18), lingual (5, 12, 17, 21-22, 24, 28), occlusal (1, 4, 8, 10, 15, 19, 23, 26), profile (3, 13-14) and basal (2, 6, 7, 9, 16, 20, 25, 27, 29).

FIGURE 11. Mobulids of the Cantaure Formation. 1-11. † Mobula fragilis (1-3, 10-11: AMU-CURS-947; 4-6: AMU-CURS-948 and 7-9: AMU-CURS-949). 12-18. Mobula cf. † M. loupianensis (AMU-CURS-946). 19-23. Mobula sp. (AMU-CURS-927). 24-27. † Plinthicus stenodon (AMU-CURS-715). Jaw position: indet. (1-27). View: labial (1, 4, 7, 10, 14-15, 17, 22, 24), lingual (3, 6, 20, 26), occlusal (2, 5, 8, 16, 19, 25), profile (11-12, 18, 23, 27) and basal (9, 13, 21).
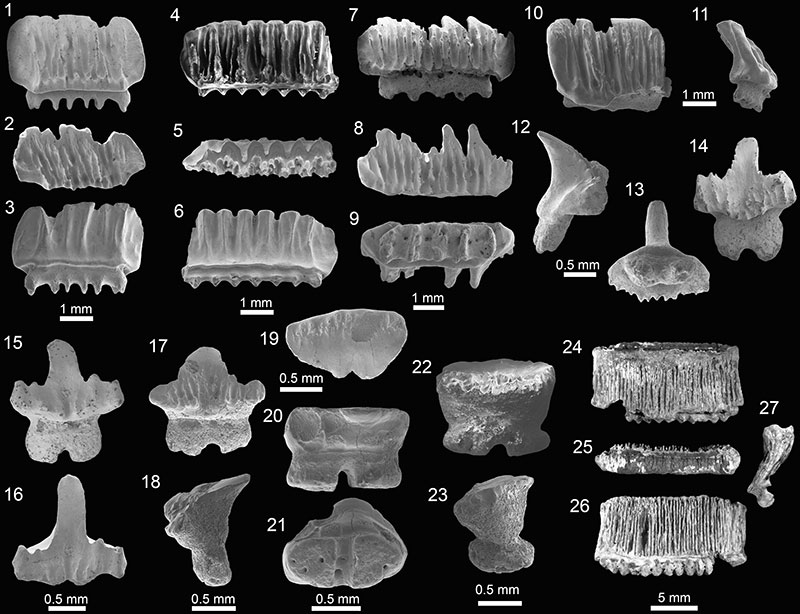
FIGURE 12. Myliobatiformes and other chondrichthyans of the Cantaure Formation. 1-6. Myliobatiformes indet. (AMU-CURS-507). 7-8. vertebra of chondrichthyan Indet. (AMU-CURS-928). 9-10. tooth of † Galeocerdo aduncus in association with barnacles of the family Balanidae (AMU-CURS-720). Jaw position: indet. (9). View: lingual (9), dorsal (1, 4, 5-6), ventral (2-3) and indet. (7-8).

FIGURE 13.Bathymetric ranges estimated for Cantaure Formation taxa and their relative abundance. Arrows indicate that the bathymetric range is greater; question mark symbol indicates uncertainty about the extent of the range. A dashed red line indicates that there is no information for the taxa. The gray shadow indicates the more probably depth range for this assemblage. In the relative abundance graphic the y-axis is shortened between 2 and 3 to improve the values visualization; note the greater abundance of shallow water taxa. The extinct species including P. benedenii, C. megalodon, P. contortus, and P. stenodon, and taxa without clear identification, including Carcharhinus spp., Carcharhinidae indet., Myliobatiformes indet. and Chondrichthyes indet., were removed from the analysis.
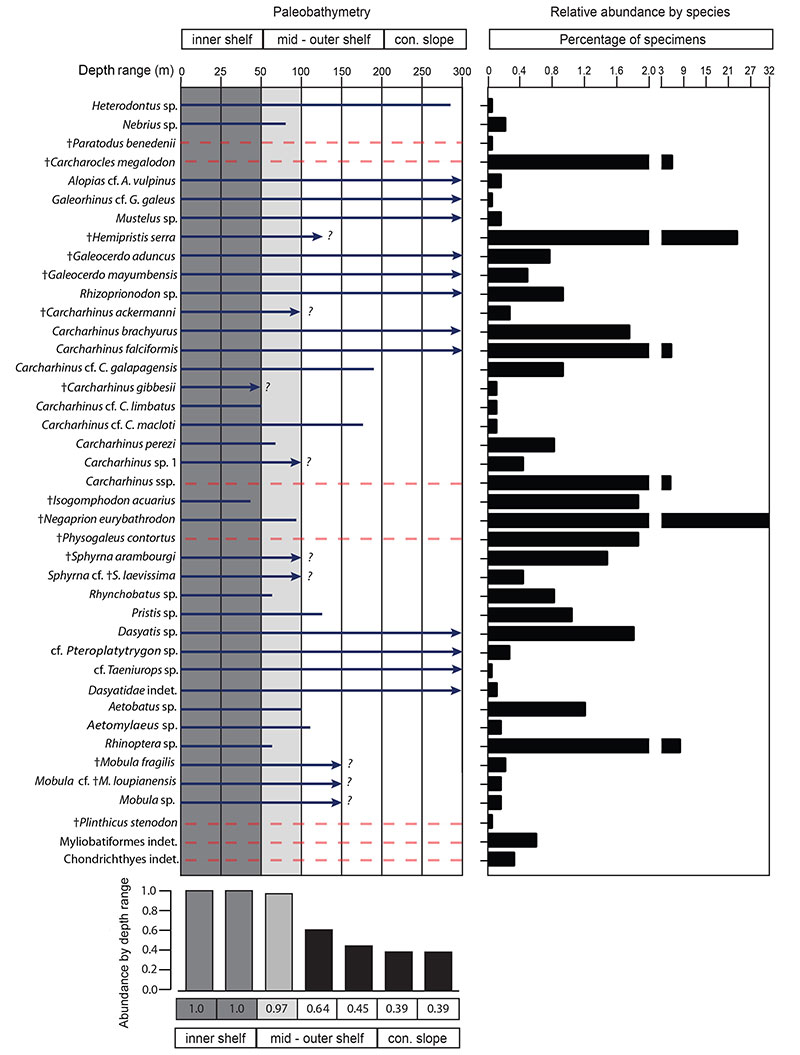
FIGURE 14. Dietary preference composition of the elasmobranchs from the Cantaure Formation. Percentages do not include Carcharhinus spp., Myliobatiformes indet. and Chondrichthyes indet.
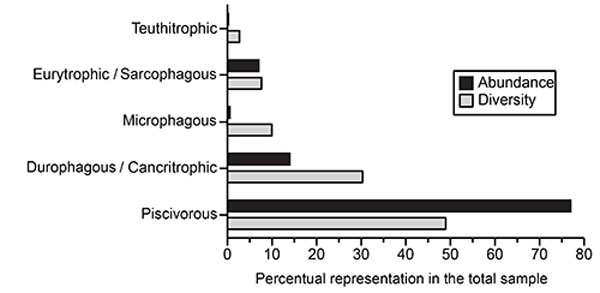
FIGURE 15. Early Neogene chondrichthyan diversity in the Americas. Tropical America: Barbados (Ba), Brazil (Br), Colombia (Co), Costa Rica (Cs), Cuba (Cu), The Grenadines (Gr), Panamá (Pa), Perú (Pu), Trinidad (Tr) and Venezuela (Ve). North America: United States of America (US). Southern South America: Argentina (Ar) and Chile (Ch). Aquitanian (Anq), Burdigalian (Brd), Langhian (Lngh); early Miocene without differentiation (EM), early-Middle Miocene (E-MM). Formation(Fm.), early (E). Note: to references see “Paleodiversity significance” (see Discusion).
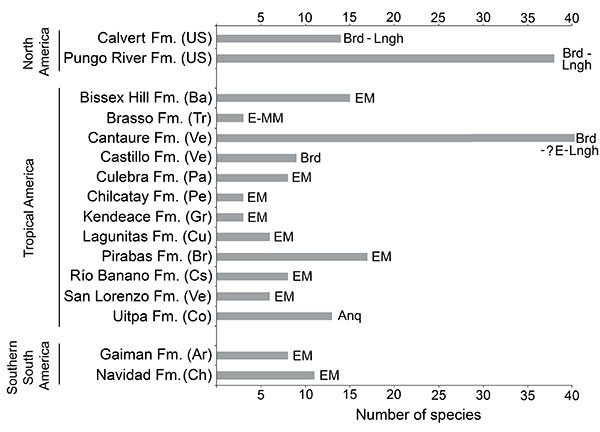
FIGURE 16. Paleobathymetric interpretation of the Cantaure Formation using several proxies. A, Foraminifera (Díaz de Gamero, 1974), B, corals (Griffiths et al., 2013), C, mollusks (Jung, 1965; Landau et al., in press), D, crustaceans (Aguilera et al., 2010), E, sharks (this study), F, teleosteans (Nolf and Aguilera, 1998, Aguilera and Rodrigues de Aguilera, 2001).
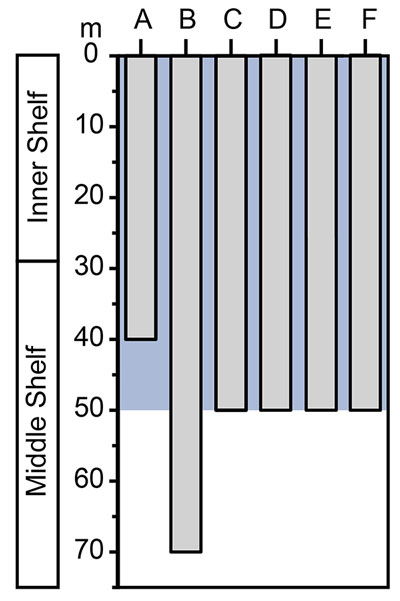
TABLE 1. Elasmobranchii paleodiversity of the Cantaure Formation.
| Superorder | Order | Family | Genus | Taxon |
| Galeomorphii | Heterodontiformes | Heterodontidae | Heterodontus | Heterodontus sp. |
| Orectolobiformes | Ginglymostomatidae | Nebrius | Nebrius sp. | |
| Lamniformes | †Otodontidae | †Paratodus | †Paratodus benedenii (Le Hon, 1871) | |
| †Carcharocles | †Carcharocles megalodon (Agassiz, 1843) | |||
| Alopiidae | Alopias | Alopias cf. A. vulpinus (Bonnaterre, 1788) | ||
| Carcharhiniformes | Triakidae | Galeorhinus | Galeorhinus cf. G. galeus (Linnaeus, 1758) | |
| Mustelus | Mustelus sp. | |||
| Hemigaleidae | Hemipristis | †Hemipristis serra (Agassiz, 1835) | ||
| Carcharhinidae | Galeocerdo | †Galeocerdo aduncus Agassiz, 1843 | ||
| †Galeocerdo mayumbensis Dartevelle and Casier, 1943 | ||||
| Rhizoprionodon | Rhizoprionodon sp. | |||
| Carcharhinus | †Carcharhinus ackermannii Santos & Travassos, 1960 | |||
| Carcharhinus brachyurus (Günther, 1870) | ||||
| Carcharhinus falciformis (Müller and Henle, 1839) | ||||
| Carcharhinus cf. C. galapagensis (Snodgrass and Heller, 1905) | ||||
| †Carcharhinus gibbesii (Woodward, 1889) | ||||
| Carcharhinus cf. C. limbatus (Müller and Henle, 1839) | ||||
| Carcharhinus cf. C. macloti (Müller and Henle, 1839) | ||||
| Carcharhinus perezi (Poey, 1876) | ||||
| Carcharhinus sp. 1 | ||||
| Carcharhinus spp. | ||||
| Isogomphodon | †Isogomphodon acuarius (Probst, 1879) | |||
| Negaprion | †Negaprion eurybathrodon (Blake, 1862) | |||
| †Physogaleus | †Physogaleus contortus (Gibbes, 1849) | |||
| Gen. indet. | sp. indet. | |||
| Sphyrnidae | Sphyrna | †Sphyrna arambourgi Cappetta, 1970 | ||
| Sphyrna cf. †S. laevissima (Cope, 1867) | ||||
| Batomorphii | Rajiformes | Rhynchobatidae | Rhynchobatus | Rynchobatus sp. |
| Pristidae | Pristis | Pristis sp. | ||
| Myliobatiformes | Dasyatidae | Dasyatis | Dasyatis sp. | |
| cf. Pteroplatytrygon | cf. Pteroplatytrygon sp. | |||
| cf. Taeniurops | cf. Taeniurops sp. | |||
| Dasyatidae Indet. | ||||
| Myliobatidae | Aetobatus | Aetobatus sp. | ||
| Aetomylaeus | Aetomylaeus sp. | |||
| Rhinopteridae | Rhinoptera | Rhinoptera sp. | ||
| Mobulidae | Mobula | †Mobula fragilis (Cappetta, 1970) | ||
| Mobula cf. †M. loupianensis (Cappetta, 1970) | ||||
| Mobula sp. | ||||
| †Plinthicus | †Plinthicus stenodon Cope, 1869 | |||
| Myliobatiformes indet. | ||||
| Chondrichthyes indet. |
APPENDIX 1.
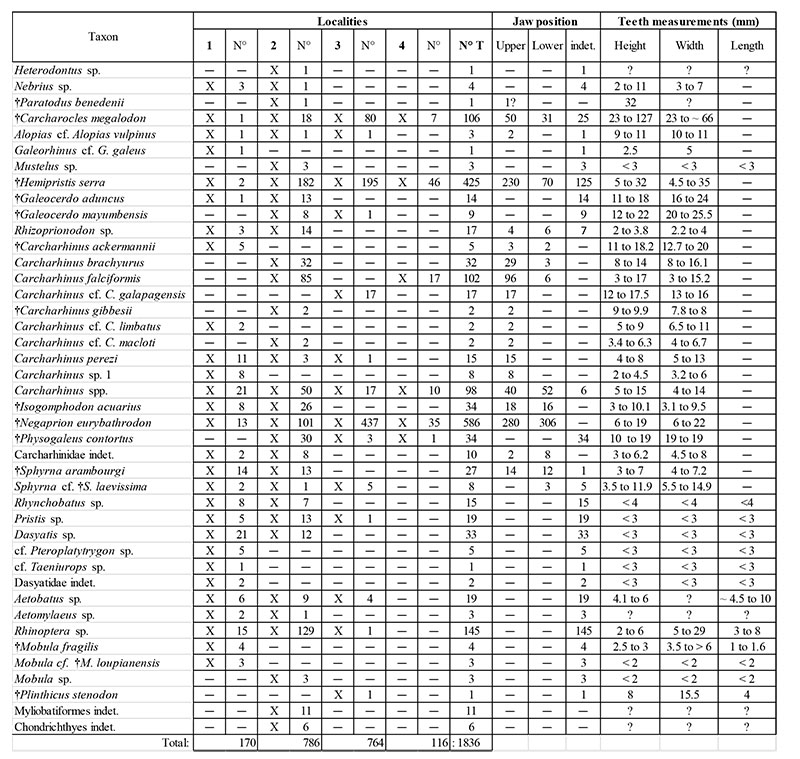
Elasmobranchii of the Cantaure Formation and their record per localities, jaw position and tooth measurements. Abbreviations: number of specimens (N°), total number of specimens (N° T) and indeterminate (indet.).
APPENDIX 2.
Referred fossil specimens and collection numbers.


APPENDIX 3.
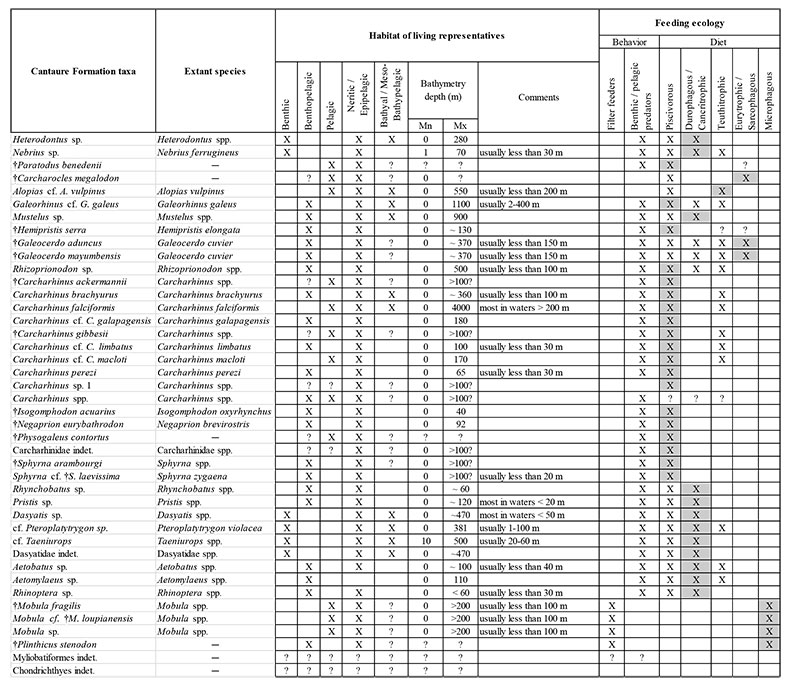
Bathymetric and feeding preferences of the Cantaure Formation elasmobranch taxa, based on the biology of their extant relatives. The gray shadow indicates the feeding preferences. Abbreviations: minimun (Mn), maximun (Mx) and meters (m).
APPENDIX 4.
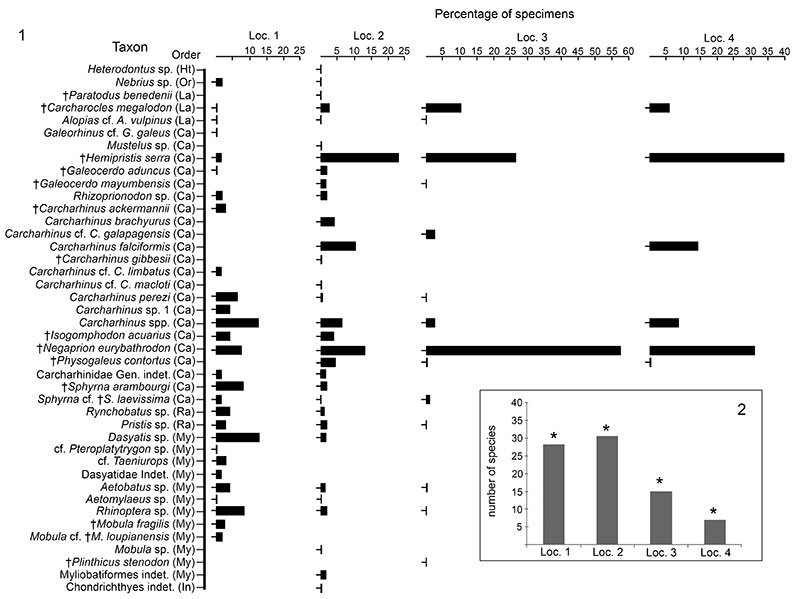
Relative abundance of elasmobranchs in the Cantaure Formation. 1. Abundance percentage per locality per taxon. 2. Total abundance per locality. Ht: Heterodontiformes; Or: Orectolobiformes; La: Lamniformes; Ca: Carcharhiniformes; Ra: Rajiformes; My: Myliobatiformes; In: Indeterminate; Loc.: locality; †: extint taxon. *: Percentage does not include Carcharhinus spp., Myliobatiformes indet. and Chondrichthyes indet.
An Early Neogene Elasmobranch fauna from the southern Caribbean (Western Venezuela)
Plain Language Abstract
A total of 39 species of fossil sharks and rays are documented from sediments of the Cantaure Formation located in North Western Venezuela. The age of the fossils spans from about 20 to 16 million years ago. The most common sharks found in the Cantaure Formation were fish-eaters, and their environment is interpreted as shallow-water with a high productivity also sustaining a high diversity of fish and invertebrates. The large shark "Megalodon" was found in sediments relatively old (~16.5 million years ago).
Resumen en Español
Una fauna de elasmobranquios del Neógeno temprano del sur del Caribe (Venezuela Occidental)
La Formación Cantaure (Burdigaliense / Langhiense temprano?) se localiza en la cuenca de Falcón, al noroeste de Venezuela, e incluye una de las faunas neógenas más diversas de teleósteos y de invertebrados bentónicos en América Tropical. Las preferencias paleoambientales de los miembros de esta fauna, así como los datos aportados por las reconstrucciones paleogeográficas publicadas, sugieren que la Formación Cantaure se depositó en un ambiente acuático somero altamente productivo, asociado con afloramiento (upwelling) en la costa. Se documentó una paleodiversidad de 39 especies de tiburones y rayas, entre ellos 15 taxones no documentados anteriormente para Venezuela y seis no documentados anteriormente para la América tropical. Se realizó un análisis batimétrico de la asociación fósil basado en la distribución de los condrictios actuales estrechamente relacionados (corelatos) con los taxones fósiles y se discute el papel ecológico y el valor estratigráfico de estos. Nuestros resultados apoyan la hipótesis de que la Formación Cantaure se depositó en un ambiente de plataforma interna-media insular. La fauna de elasmobranquios se caracteriza por el predominio de los tiburones bentopelágicos con preferencias piscívoras (por ejemplo, †Paratodus, Galeorhinus, Hemipristis, Rhizoprionodon, Carcharhinus, Isogomphodon, Negaprion, †Physogaleus y Sphyrna) seguidos por los durófagos/consumidores de crustáceos (por ejemplo, Heterodontus, Nebrius, Mustelus, Rhynchobatus, Pristis, Dasyatis, cf. Pteroplatytrygon, cf. Taeniurops, Aetobatus, Aetomylaeus y Rhinoptera). También se han documentado especies filtradoras (por ejemplo, Mobula y †Plinthicus), euritróficas/sarcófagas (por ejemplo, †Carcharocles y Galeocerdo) y devoradores de cefalópodos (por ejemplo, Alopias). Los dientes de Carcharocles megalodon encontrados en los sedimentos burdigalienses de la Formación Cantaure son indicativos de que esta especie estaba ya presente en el Mioceno temprano. Algunos taxones (Nebrius, Carcharhinus cf. C. macloti y Rhynchobatus) están ausentes de la fauna actual del Caribe y del Atlántico Occidental, pero estaban presentes en la región antes del cierre de la Vía marítima de América Central.
Palabras clave: Mioceno Temprano; Burdigaliense; Formación Cantaure; tiburones; rayas; Megalodon; paleoambiente
Traducción: Enrique Peñalver (Sociedad Española de Paleontología)
Résumé en Français
Une faune d'élasmobranches du Néogène ancien du sud des Caraïbes (ouest du Venezuela)
La formation de Cantaure (datée du Burdigalien au Langhien ancien?) est située dans le bassin de Falcón, nord-ouest du Venezuela, et inclut une des faunes de téléostéens et d'invertébrés benthiques les plus diverses du Néogène d'Amérique tropicale. Les préférences paléoenvironnementales des membres de cette faune et les reconstitutions paléogéographiques publiées suggèrent que la formation de Cantaure a été déposée dans un environnement d'eau peu profonde à haute productivité, associé à des remontées d'eau profonde (« upwelling ») près des côtes. Nous documentons une paléodiversité de 39 espèces de requins et de raies, dont 15 taxons jamais signalés jusqu'à présent pour le Venezuela, et six pour l'Amérique tropicale. Nous avons effectué une analyse bathymétrique de l'assemblage fossile basée sur la distribution des espèces actuelles de chondrichthyens étroitement apparentées aux taxons fossiles et nous discutons le rôle écologique et les implications stratigraphiques de cette analyse. Nos résultats soutiennent l'hypothèse que la formation de Cantaure a été déposée dans un environnement insulaire de plateforme interne et moyenne. La faune d'élasmobranches est caractérisée par une prédominance de requins benthopélagiques piscivores (e.g., †Paratodus, Galeorhinus, Hemipristis, Rhizoprionodon, Carcharhinus, Isogomphodon, Negaprion, †Physogaleus, et Sphyrna) suivis par des durophages et des spécialistes des crustacés (e.g., Heterodontus, Nebrius, Mustelus, Rhynchobatus, Pristis, Dasyatis, cf. Pteroplatytrygon, cf. Taeniurops, Aetobatus, Aetomylaeus, et Rhinoptera). Des espèces filtreuses (e.g., Mobula et †Plinthicus), eurytrophes/carnivores (e.g., †Carcharocles et Galeocerdo), et se nourrissant de céphalopodes (e.g., Alopias) ont aussi été trouvées. Des dents de Carcharocles megalodon trouvées dans les sédiments burdigaliens de la formation de Cantaure soutiennent la présence de cette espèce dès le Miocène ancien. Certains taxons (Nebrius, Carcharhinus cf. C. macloti, et Rhynchobatus) sont absents de la faune actuelle des Caraïbes et de l'Atlantique occidental, mais elles étaient présentes dans la région avant la fermeture de la mer d'Amérique centrale.
Mots-clés : Miocène ancien ; Burdigalien ; formation de Cantaure ; requins ; raies ; mégalodon ; paléoenvironnement
Translator:Antoine Souron
Deutsche Zusammenfassung
Eine früh-neogene Knorpelfisch-Fauna aus der südlichen Karibik (westliches Venezuela)
Die Cantaure Formation (Burdigalium bis ?frühes Langhium) liegt im Falcón Becken, nordwestliches Venezuela, und beinhaltet eine der diversesten neogenen Teleostier-und benthonischen Invertebratenfaunen des Tropischen Amerikas. Sowohl die Paläoumwelt-Präferenzen der Mitglieder dieser Fauna, als auch publizierte paläogeographische Rekonstruktionen legen nahe, dass die Cantaure Formation in einem hochproduktiven Flachwassermilieu, das mit küstennahem Upwelling assoziiert war, abgelagert wurde. Wir dokumentierten eine Paläodiversität von 39 Hai-und Rochenarten, die 15 bisher unveröffentlichte Taxa für Venezuela und sechs für das Tropische Amerika umfassen. Basierend auf der Verteilung von heutigen Knorpelfischen, die nahe mit den Fossilen Arten verwandt sind, führten wir eine bathymetrische Analyse der Fossilassemblage durch und diskutieren die ökologische Rolle und stratigraphische Signifikanz. Unsere Ergebnisse unterstützen die Hypothese, dass die Cantaure Formation in einem insularen inneren-mittleren Schelfmilieu abgelagert wurde. Die Knorpelfisch-Fauna wird dominiert von benthopelagischen Haien mit piscivoren Fressgewohnheiten (z.B., †Paratodus, Galeorhinus, Hemipristis, Rhizoprionodon, Carcharhinus, Isogomphodon, Negaprion, †Physogaleus und Sphyrna) gefolgt von durophagen/cancritrophischen Arten (z.B., Heterodontus, Nebrius, Mustelus, Rhynchobatus, Pristis, Dasyatis, cf. Pteroplatytrygon, cf. Taeniurops, Aetobatus, Aetomylaeus und Rhinoptera). Filterer (z.B., Mobula and †Plinthicus), eurytrophische/sarcophage (z.B., †Carcharocles und Galeocerdo) und teuthitrophische (z.B., Alopias) Arten wurden ebenfalls gefunden. Zähne von Carcharocles megalodon aus Sedimenten des Burdigaliums der Cantaure Formation unterstützen das Vorkommen dieser Art bereits im frühen Miozän. Einige Taxa (Nebrius, Carcharhinus cf. C. macloti und Rhynchobatus) kommen in der heutigen Karibik und westlichen atlantischen Fauna nicht vor, waren jedoch in der Region vor der Schließung des zentralamerikanischen Seewegs präsent.
Schlüsselwörter: Frühes Miozän; Burdigalium; Cantaure Formation; Haie; Rochen; Megalodon; Paläoumwelt
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024