

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
APPENDIX 1
List of characters used to on the phylogenetic analysis performed in this work. Characters 111 to 113 are proposed in this manuscript, the others are from Keivany and Nelson (2006, appendix 2);
1. Shape of nasal: 0) Simple, flat; 1) With anterior lamina; 2) Absent.
2. Lateral ethmoid: 0) Not extended to the orbit; 1) Extended to the orbit.
3. Vomer location: 0) Between ethmoids; 1) Superficially anteriorly.
4. Lachrymal: 0) Short; 1) Elongated.
5. Infraorbitals: 0) Discontinuous; 1) Continuous.
6. Frontal lateral postorbital process: 0) Absent; 1) Present.
7. Frontal: 0) Does not meet paresphenoid lateral process; 1) Meeting paresphenoid lateral process.
8. Parietal: 0) Present; 1) Absent.
9. Intercalar: 0) Present; 1) Absent.
10. Prootic and exoccipital: 0) Connected; 1) Widely separated by pterotic.
11. Basisphenoid: 0) Present; 1) Absent.
12. Pterosphenoid: 0) Present; 1) Absent.
13. Paresphenoid: 0) Narrow; 1) Expanded between the lateral ethmoids.
14. Paresphenoid shaft: 0) Divided posteriorly; 1) Entire.
15. Occipital condyle: 0) Concave; 1) Convex.
16. Exoccipital elongated posterior process: 0) Absent; 1) Present.
17. Spina occipitalis: 0) Present; 1) Absent.
18. Teeth on the jaws: 0) Present; 1) Absent.
19. Premaxillary ascending process: 0) Well developed; 1) Absent or reduced.
20. Premaxillary articular process: 0) Present; 1) Absent.
21. Premaxillary postmaxillary process: 0) Absent; 1) Present.
22. Maxillary shaft: 0) Expanded ventrally; 1) Uniform.
23. Maxillary posterior process: 0) Present; 1) With an anterior lamina; 1) Absent.
24. Palatine: 0) Separated; 1) United at the midline and to vomer.
25. Palatine head: 0) Cylindrical; 1) Conical.
26. A separatedentopterygoid: 0) Present; 1) Absent.
27. Metapterygoid: 0) Separate; 1) Fused to hyomandibular or sympletic.
28. Metapterygoid: 0) Posterior; 1) Anterior to the orbit.
29. Quadrate shaft: 0) Lacks a deep dorsal flange; 1) Bears a deep dorsal flange.
30. Length of the quadrate shaft: 0) About the same that quadrate body; 1) Much longer than quadrate body.
31. Articulation of quadrate and lower jaw: 0) Below the orbit; 1) In front of the orbit.
32. Sympletic: 0) Lacking a dorsal or ventral flange; 1) Bearing a dorsal or ventral flange; 2) Bifurcated anteriorly.
33. Hyomandibular: 0) Lacking a large medial lamina; 1) Bearing a large medial lamina.
34. Hyomandibular: 0) Bearing a lateral lamina; 1) Lacking a lateral lamina.
35. Hyomandibular: 0) Bearing distinct cranial condyles; 1) Lacking distinct cranial condyles.
36. Preopercle: 0) Without socket at articulation with interhyal; 1) With socket at articulation with interhyal.
37. Ventral and dorsal arms of the preopercle: 0) Different sized; 1) Equal sized.
38. Preopercular canal: 0) Enclose; 1) Open.
39. Ascending limb of subopercle: 0) Present; 1) Absent.
40. Subopercle and interopercle: 0) Close together; 1) Widely separating from each other.
41. Interopercle: 0) Short; 1) Elongated.
42. Posteroventral expansion of the interopercle: 0) Absent; 1) Present.
43. Basihyal: 0) Small; 1) Elongated; 2) With enlarged anterior cartilage.
44. Urohyal: 0) Without ventrolateral flanges; 1) With ventrolateral flanges.
45. Urohyal blade: 0) Entire; 1) Incised posteriorly.
46. Hypohyals: 0) Overlap anterior ceratohyal; 1) Articulate with posterior ceratohyal.
47. Anterior ceratohyal: 0) Abruptly becoming narrower; 1) Even or gradually becoming narrower anteriorly.
48. Struts that connects anterior and posterior ceratohyals: 0) Present; 1) Absent.
49. Interhyal: 0) Cylindrical and free; 1) Round and sutured to the posterior ceratohyal.
50. Branchiostegal rays: 0) More than four; 1) Four or fewer.
51. Gill membrane: 0) Free from isthmus; 1) United to isthmus.
52. Ossified basibranchials: 0) Three; 1) Less than three.
53. A fourth cartilaginous basibranchials: 0) Present; 1) Absent.
54. Hypobranchials: 0) Three; 1) Fewer than three.
55. Epibranchial 1: 0) With uncinated process; 1) Without uncinated process.
56. Epibranchial 1: 0) Separate to pharyngeal tooth plate 2; 1) Attached to pharyngeal tooth plate 2.
57. Uncinated process of epibranchial 3 and 4: 0) Present; 1) Absent.
58. Epibranchial 3 and 4: 0) Not associated with their processes or heads; 1) Associated with their processes or heads.
59. Epibranchial 4: 0) Normal; 1) Enlarged; 2) Absent.
60. Epibranchial 4 head: 0) Larger than three; 1) Normal.
61. Pharyngobranchial 1: 0) Present; 1) Absent.
62. Pharyngeal tooth plate 2: 0) Present; 1) Absent.
63. Pharyngeal tooth plate 4: 0) Present; 1) Absent.
64. Interarcual cartilage: 0) Absent; 1) Present.
65. Gill filaments: 0) Normal; 1) Lophobranch with skeleton fused basally.
66. Tooth plates on the branchial arches: 0) Present. 1) Absent.
67. Posttemporal dorsal process: 0) Tightly attached with short ligament to epioccipital; 1) Ossified to cranium.
68. Posttemporal sensory canal: 0) Absent; 1) Present.
69. Extrascapular: 0) Present; 1) Absent.
70. Supracleithrum: 0) Normal; 1) Reduced or absent.
71. Pointed anterodorsal ramus of the cleithrum: 0) Present; 1) Absent.
72. Cleithrum: 0) Entire; 1) Divided into two struts ventrally.
73. Cleithrum: 0) Without posteromedial extension to coracoids; 1) With posteromedial extension to coracoids.
74. Postcleithrum number: 0) Two; 1) One; 2) Absent.
75. Scapular foramen: 0) Complete; 1) Incomplete.
76. Scapula and first actinost: 0) Separate; 1) Fused together.
77. Coracoid ventral flange: 0) Present; 1) Absent.
78. Ectocoracoid: 0) Absent; 1) Present.
79. Shape of actinost: 0) Normal; 1) Strongly hourglass-shaped.
80. Size of actinosts: 0) Different; 1) Similar.
81. Position of actinosts: 0) Normal; 1) Rotated laterally.
82. Anteroventral spike of actinost 4: 0) Absent; 1) Present.
83. Actinost 4 and coracoids sutured to each other: 0) Absent; 1) Present.
84. Pelvic spines: 0) Present; 1) Absent.
85. Pelvic plates: 0) Separated; 1) Joined by a suture.
86. Pelvic posterior process: 0) Present; 1) Absent.
87. Pelvic plate: 0) Lacking anterior process; 1) Bearing an anterior process.
88. Dorsal spines: 0) Present; 1) Absent.
89. Dorsal posterior basals: 0) Absent; 1) Present.
90. Dorsal spine distal pterygiophores: 0) Not expanding; 1) Expanding.
91. Number of anal spines: 0) More than one; 1) One; 2) Absent.
92. Dorsal part of occipital condyle: 0) Made of exoccipitals and the first centrum bearing a dorsal facet that articulate with the exoccipital condyles; 1) Occipital condyle consisting only of basioccipital and the first vertebra lacks the dorsal facet, but bears lateral facets that articulate with exoccipital; 2) The first vertebra directly connected only to the basioccipital.
93. Anterior centra: 0) Separate; 1) Sutured together.
94. Transverse process on the first two vertebrae: 0) Present; 1) Absent.
95. Anterior vertebrae: 0) Normal; 1) Longer than posterior one.
96. Anterior vertebrae: 0) Lack lateral process to scutes; 1) Bear lateral processes to scutes.
97. Neural arch of precaudal vertebra: 0) Not pierced by foramina; 1) Pierced by foramina.
98. Neurohypophyses: 0) Present; 1) Absent.
99. Number of supraneural bones: 0) Three; 1) One or two; 2) Absent.
100. Pleural ribs: 0) Present; 1) Absent.
101. Epineurals and epipleurals: 0) Both present; 1) Only epineurals present; 2) Both absent.
102. Baudelot’s ligament: 0) Originates on the first vertebra; 1) Originates on exoccipital; 2) Absent.
103. Bony scutes: 0) Absent; 1) Present.
104. Parhypural: 0) Autogenous; 1) Fused to centrum or hypurals.
105. Condition of hypurals: 0) Autogenous; 1) All fused to centrum.
106. Number of hypural plates: 0) Three or more; 1) Two; 2) One.
107. Condition of hypurapophysis: 0) Distinct; 1) Indistinct.
108. Length of neural spine of preural 2: 0) Shorter than other preneurals; 1) The same size as other preneurals.
109. Condition of the uroneural 1: 0) Autogenous; 1) Fused.
110. Caudal cartilage: 0) Present; 1) Absent.
111. Interspace of dorsal spines: 0) Separate; 1) Close together.
112. Snout: 0) Nude; 1) Covered with dermal scutes or scales.
113. Condition of postcleithrum: 0) Not posteriorly elongate (it does not reach the level of the posterior tip of the postcleithrum); 1) Posteriorly elongate (it reaches the posterior level of the postcleithrum).
APPENDIX 2
List of states of characters considered to perform the phylogenetic analysis of the present paper (Figure 6). Polymorphic states are coded as * = 0/1, # = 0/2, @ = 1/2.
Atheriniformes:
0000000000000000010110000100001000000000020000000000000001010110100010*0***00000000000000100000*000000**000*0-00.
Elassomatiformes:
0000000000100100000000000100000000000000000000010000000100000000000010000000000?000011000001000000000000010010-00.
Synbranchiformes:
0100000010000000100*00100*00000*00*01110000*0000001**01*0*0*010*000*001012001000000????*00#00000000*0100001*01-00.
Hypoptychidae:
01000100001011001*01111001000001010001000010001101?01011000100100000111012000000010????10?20000010000101121111-00.
Aulorhynchidae:
11011110*01*01000001*1101110011201*00100101000100100*011000100*00101*10112101101010010101112000*10**11111@1111-00.
Gasterosteidae:
11010110001101000001011011100112000001001110000001100011000100100000010012101101011010101112000010100111121111000.
Pegasidae:
11011000101111000110011??110000110101101010000100011111?100101101010010002100001011011110?20011001011111121011-00.
Solenostomidae:
201100011110110111100011001111111010011111?01011010111????2?00101000010102100011000101110?20011001011111121011-00.
Syngnathidae:
211101111110110011100110001111111011001110010110111111111?2?01101010011012100011000????10?20000111011111121011-00.
Indostomidae:
010101?110100001101101100110001000000111001000010000001110001100101001101111000101010110?120000101110111121011-00.
Aulostomidae:
011100011110100101100000010111110010010010110100011110011?2?00010101011111111111000101110?00111000010100011110001.
Fistulariidae:
211100001110100101100010001111110010010010111110001110111?2?00010011010111101111000101110?20111000011101121101-01.
Macrorhamphosidae:
11110001111011100110011000111110101101001010101110000010010100000010011001010001111010100020001100111111121111-00.
Centriscidae: 0111000011101110011001100011111010110100101011111010101101011000001001101101000 1111001100?20111111011110121111-00.
Eekaulostomus cuevasae gen. and sp. nov.:
?11???0????01?????0??0??0???1?1?011?01??10????????????????????????????0011???1?????1???0??0???????????1???????111.
APPENDIX 3
Data matrix considered to perform the phylogenetic analyzes on the present work in NEXUS file format. The characters 1-110 and compared taxa are retrieved from Keivany and Nelson (2006). The characters 111-113 and Eekaulostomus cuevasae gen. and sp. nov. are added in this manuscript. Polymorphic states are represented by numbers separated by a slash (/) symbol. This zipped file is available for download.
TABLE 1. Measurements and body proportions of IGM 4716, holotype and single specimen known of Eekaulostomus cuevasae gen. and sp. nov.
| Measurements | ||
| in mm | as % of SL | |
| Total length | 79.64 | - |
| Standard length (SL) | 72.76 | - |
| Head length | 26.73 | 36.73 |
| Postorbital length | 51.50 | 70.78 |
| Orbital length | 3.95 | 5.42 |
| Preorbital length | 17.17 | 23.59 |
| Prepelvic length | 33.49 | 46.02 |
| Predorsal length | 58.85 | 80.88 |
| Dorsal fin length | 5.44 | 7.48 |
| Preanal length | 56.98 | 78.31 |
| Anal fin length | 5.01 | 6.88 |
| Head height | 8.51 | 11.69 |
| Maximum body height | 12.14 | 16.68 |
| Caudal peduncle height | 5.02 | 6.89 |
| Postdorsal length | 7.15 | 9.83 |
| Postanal length | 9.33 | 12.82 |
TABLE 2. Comparison between results of phylogenetic analyses on Gasterosteiformes published by Keivany and Nelson (2006, figure 1), results of the same essay obtained in this work, and obtained after the inclusion of Eekaulostomus cuevasae gen. and sp. nov. and characters 111 to 113. The topology of trees in these three analyses is similar (see Figure 6) except for node J after the inclusion of E. cuevasae. Synapormorphies are underlined and characters with different positions throughout these comparisons are in bold.
| Published by Keivany and Nelson (2006). | Replay of Keivany and Nelson, (2006), in this work. | Present work. | |
| 308 steps, CI=0.5 | 271steps, CI = 0.43 | 279 steps; CI = 0.43 | |
| A
|
43-1, 47-1, 70-1, 80-1, 82-1, 91-2, 104-1, 105-1, 106-1 | 6-1, 11-1, 14-1, 20-1, 22-1, 43-1, 47-1, 50-1, 60-1, 63-1, 70-1, 82-1, 87-1, 90-1, 91-2, 97-1, 104-1, 105-1, 106-1, 108-1 | 6-1, 11-1, 14-1, 20-1, 22-1, 43-1, 47-1, 50-1, 60-1, 63-1, 70-1, 82-1, 87-1, 90-1, 91-2, 97-1, 104-1, 105-1, 106-1, 108-1 |
| B | - | 4-1, 27-1, 31-1, 75-1, 80-1, 99-1, 103-1 | 4-1, 27-1, 31-1, 75-1, 80-1, 99-1, 103-1 |
| C | 7-1, 25-1, 32-2, 41-1, 78-1, 89-1, 91-1, 92-2 | 1-1, 7-1, 12-1, 25-1 , 30-1, 32-2, 41-1, 71-0, 77-1, 78-1, 85-1, 89-1, 91-1, 92-2 | 1-1, 7-1, 12-1, 25-1 , 30-1, 32-2, 41-1, 77-1, 78-1, 85-1, 89-1, 91-1, 92-2 |
| D | 9-1, 19-1, 57-1, 86-1, 98-1, 100-1 | 8-1, 9-1, 19-1, 40-1, 50-0, 57-1, 62-1, 65-1, 67-1, 84-1, 86-1, 97-0, 98-1, 100-1, 108-0 | 8-1, 9-1, 19-1, 40-1, 50-0, 57-1, 62-1, 65-1, 67-1, 84-1, 86-1, 97-0, 98-1, 100-1, 108-0 |
| E | - | 6-0, 13-1, 18-1, 20-0, 33-1, 35-1, 43-0, 51-1, 52-1, 53-1, 54-1, 88-1, 90-0, 94-1, 95-1, 99-0, 101-1 | 6-0, 13-1, 18-1, 20-0, 33-1, 35-1, 43-0, 51-1, 52-1, 53-1, 54-1, 88-1, 90-0, 94-1, 95-1, 99-0, 101-1 |
| F | 3-1, 10-1, 28-1, 29-1, 30-1, 41-1 | 3-1, 10-1, 26-0, 28-1, 29-1, 30-1, 41-1, 45-1, 46-1, 58-1 , 59-2, 62-0, 79-1, 82-0 | 3-1, 10-1, 26-0, 28-1, 29-1, 30-1, 41-1, 45-1, 46-1, 58-1, 59-2, 62-0, 79-1, 82-0 |
| G | 1-2, 39-1 | 1-2, 17-1, 39-1, 50-1 | |
| H | - | 40-0, 43-1, 54-0, 63-0, 65-0, 74-1, 76-1, 88-0, 93-1, 98-0, 108-1 | 40-0, 43-1, 54-0, 63-0, 65-0, 74-1, 76-1, 88-0 93-1, 98-0, 108-1 |
| I | 15-1, 36-1, 49-1, 81-1, 83-1, 96-1 | 15-1, 32-0, 36-1, 48-1, 49-1, 52-0, 57-0, 59-0, 75-0, 79-0, 81-1 , 82-1, 83-1, 84-0, 88-0, 96-1 | 15-1, 32-0, 36-1, 48-1, 49-1, 52-0, 57-0, 59-0, 71-1, 75-0, 79-0, 81-1, 82-1, 83-1, 84-0, 96-1 |
| J | - | - | 14-0, 16-1, 22-0, 33-0, 44-1, 64-1, 68-1, 77-1, 78-1, 113-1 |
| K | 16-1, 44-1, 64-1, 68-1, 72-1, 77-1, 78-1 | 14-0, 16-1, 22-0, 33-0, 44-1, 64-1, 68-1, 72-1, 77-1, 78-1, 103-0 | 72-1, 103-0 |
| Hypoptychidae | 13-1, 21-1, 34-1, 48-1, 53-1, 69-1, 88-1 | 13-1, 17-1, 21-1, 34-1, 48-1, 53-1, 69-1, 88-1 | 13-1, 17-1, 21-1, 34-1, 48-1, 53-1, 69-1, 71-1, 88-1 |
| Aulorhynchidae | - | 5-1, 34-1, 66-1, 68-1, 72-1, 101-1 | |
| Gasterosteidae | - | 42-1, 47-0, 51-1, 83-1 | 42-1, 47-0, 51-1, 83-1 |
| Indostomidae | 16-1, 39-1, 48-1, 61-1, 74-1, 76-1, 96-1 | 14-0, 16-1, 17-1, 32-0, 39-1, 47-0, 48-1, 60-0, 61-1, 63-0, 74-1, 76-1, 96-1 | 14-0, 16-1, 17-1, 32-0, 39-1, 47-0, 48-1, 60-0, 61-1, 63-0, 71-1, 74-1, 76-1, 96-1 |
| Pegasidae | 1-1, 5-1, 12-1, 37-1, 42-1, 83-1 | 1-1, 5-1, 8-0, 12-1, 31-0, 37-1, 42-1, 71-0, 73-0, 83-1, 84-0, 85-1 | 1-1, 5-1, 8-0, 12-1, 31-0, 37-1, 42-1, 73-0, 83-1, 84-0, 85-1 |
| Solenostomidae | - | 2-0, 16-1, 22-0, 24-1 , 42-1, 46-0, 48-1, 51-0, 67-0, 71-0, 72-1, 73-0 | 2-0, 16-1, 22-0, 24-1 , 42-1, 46-0, 48-1, 51-0, 67-0, 72-1, 73-0 |
| Syngnathidae | - | 6-1, 7-1, 36-1, 38-0, 44-1, 45-0, 49-1, 62-1, 94-0, 95-0, 96-1, 97-1 | 6-1, 7-1, 36-1, 38-0, 44-1, 45-0, 49-1, 62-1, 71-1, 94-0, 95-0, 96-1, 97-1 |
| Eekaulostomus | - | - | 19-0, 34-1, 91-0, 111-1, 112-1 |
| Aulostomidae | - | 23-0, 26-1, 27-0, 45-0, 47-0, 50-1, 55-0, 66-1, 67-0, 101-0, 104-0, 105-0, 106-1, 110-0 | 23-0, 26-1, 27-0, 45-0, 47-0, 50-1, 55-0, 66-1, 67-0, 71-1, 101-0, 104-0, 105-0, 106-1, 110-0 |
| Fistulariidae | - | 1-2, 8-0, 71-0, 76-0, 109-0 | 1-2, 8-0,76-0, 88-1, 109-0 |
| Macrorhamphosidae | - | 1-1, 46-0, 51-0, 53-0, 56-0, 73-0, 85-1, 86-0, 93-0, 94-0, 99-1 | 1-1, 46-0, 51-0, 53-0, 56-0, 73-0, 85-1, 86-0, 93-0, 94-0, 99-1 |
| Centriscidae | - | 8-0, 61-1, 97-1, 98-1, 104-0 | 8-0, 61-1, 97-1, 98-1, 104-0 |
TABLE 3. Morphological comparison between Eekaulostomus, and both recent and some extinct members of the superfamily Aulostomoidea (Wheeler, 1955; Fritzsche, 1976, 1981; Blot, 1980; among others). Abbreviations: Abd, abdominal.
| Eekaulostomus | Macroaulostomus | Extant Aulostomus spp. | Jungersenichthys | Aulostomoides | Eoaulostomus | Synhypuralis | Extant Fistularia spp. | |||||
| Dorsal fin | II + 8 | ? + (15)? | VIII-XIII + 21-26 | - | - | III +16-21 | II or III +14-15 | - +14-20 | ||||
| Dorsal spines | Stout | ? | Thin, large | - | - | ? | ? | No | ||||
| Anal fin | II + 8 | ? + 14? | III + 23-27 | - | - | III + 16-20 | II-III + 12-13 | 0 + 14-19 | ||||
| Anal fin spines | Stout | - | Thin | - | - | ? | ? | - | ||||
| Pectoral fin rays | 15 | - | ? | - | - | 15 | 15-16 | 13-18 | ||||
| Pelvic fin | Thoracic | Anterior? | Abd | - | Abd | Abd | ? | Abd | ||||
| Pelvic fin rays | 6 | 6 | 5 | - | - | 6 | 6 | 6 | ||||
| Caudal fin formula | iv+I+7 -- 5+I+iii |
iii+I+5 -- 5+I+iii |
? | (15)? | - | iv+I+5 -- 5+I+iv |
-- 5+I+iii |
0+I+6 -- 6+I+0 |
||||
| Caudal filament | No | No | No | No | ? | Yes | Yes | Yes | ||||
FIGURE 1. Map of the Belisario Domínguez quarry, near Palenque, Chiapas, southeastern Mexico. Yellow areas into the map show the Paleocene Tenejapa-Lacadón geological unit.
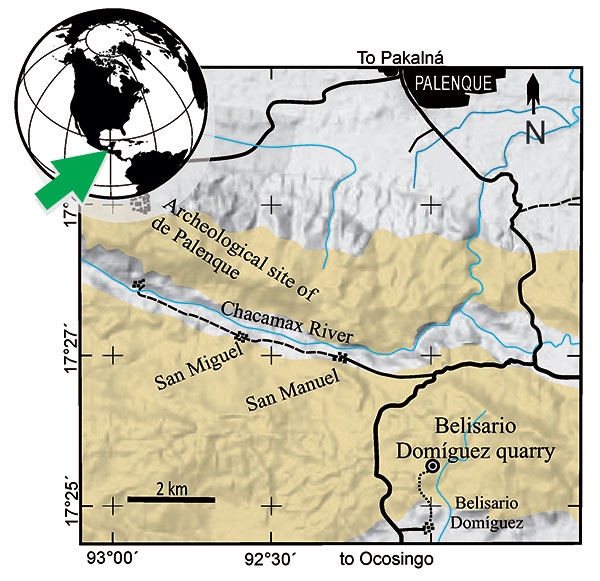
FIGURE 2. Eekaulostomus cuevasae gen. and sp. nov. from the Danian marine deposits of the Belisario Domínguez quarry, near Palenque, Chiapas, southeastern Mexico. 1, General view of IGM 4716, holotype and single specimen known. 2, Idealized line drawing of the same specimen.

FIGURE 3. Head and anterior part of the trunk of Eekaulostomus cuevasae gen. and sp. nov. from the Belisario Domínguez quarry. 1, General view of the head of specimen IGM 4716. 2, Line drawing of 1. Abbreviations: aa, anguloarticular; br, branchiostegals; cl, cleithrum; den, dentary; eccor, ectocoracoid; ecp, ectopterygoid; fr, frontal; hy, hyomandibula; iop, interopercle; la, lachrymal; le, lateral ethmoid; mes, mesethmoid; mx, maxilla; op, opercle; pa, parasphenoid; pal, palatine; pmx, premaxilla; pop, preopercle; pvfr, pelvic fin rays; q, quadrate; sc, scutes; sop, subopercle; sy, symplectic; v, vertebral centra; vo, vomer (the rays of pelvic fin are seen below the cleithrum).
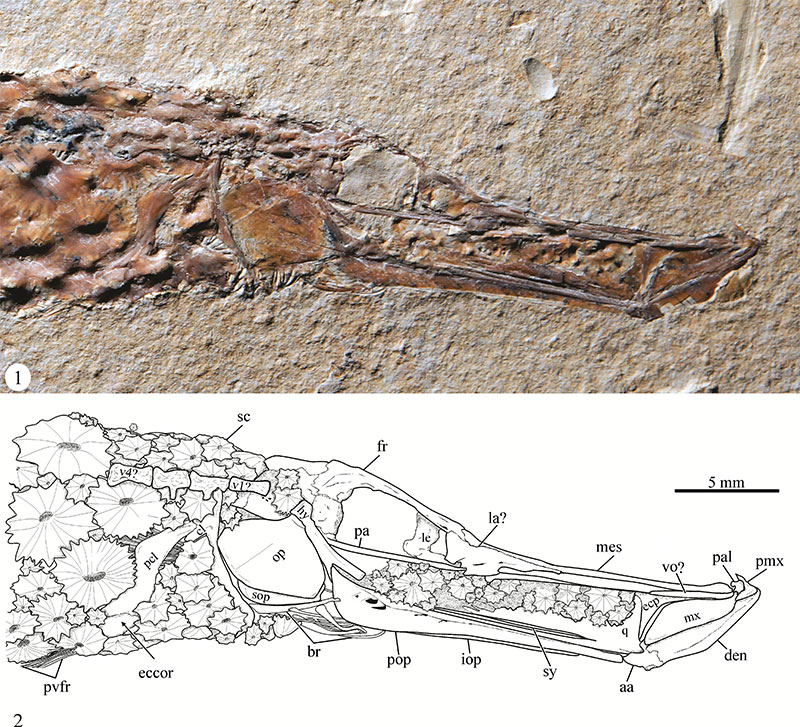
FIGURE 4. Unpaired and caudal fins of Eekaulostomus cuevasae gen. and sp. nov. from the Belisario Domínguez quarry. 1, General view of the head of specimen IGM 4716. 2, Idealized line drawing of 1. Abbreviations: afs, anal fin spines; afr, anal fin rays; dfr, dorsal fin rays; dfs, dorsal fin spines; pr, procurrent rays; black arrows enclose the principal rays of caudal fin.
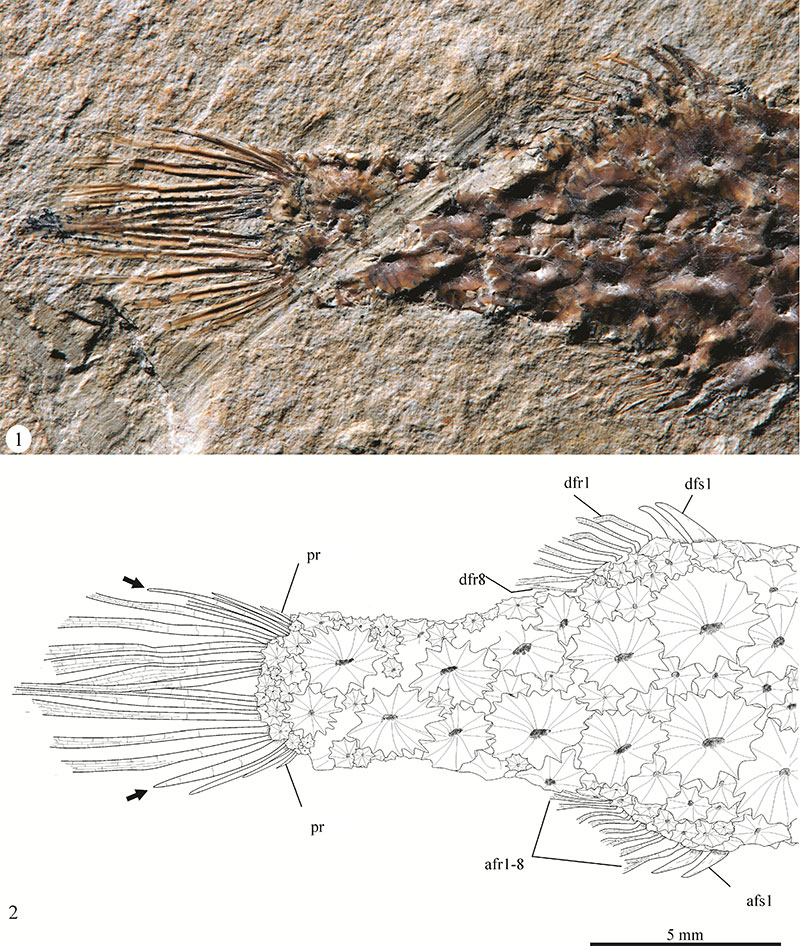
FIGURE 5. Scutes of Eekaulostomus cuevasae gen. and sp. nov. from the Belisario Domínguez quarry. 1, Close-up of the head of specimen IGM 4716, showing a scute with the external surface exposed. 2, Idealized line drawing of 1. 3, Close-up of the caudal region of the specimen IGM 4716, showing a scute with the internal surface exposed.
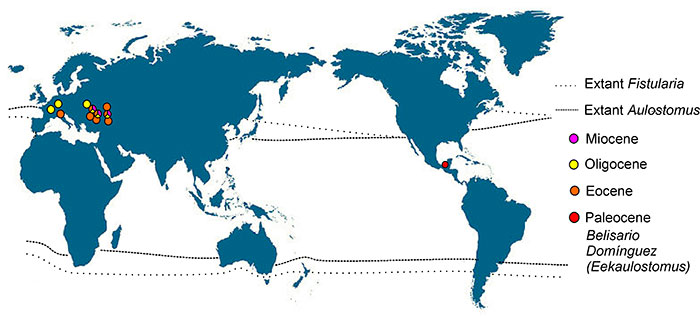
FIGURE 6. Phylogenetic relationships of Eekaulostomus cuevasae gen. and sp. nov. and the superfamily Aulostomoidea within the order Gasterosteiformes (modified from Keivany and Nelson, 2006) (check the characters supporting the nodes in Table 2; number near the nodes show the bootstrap values obtained above 50%).
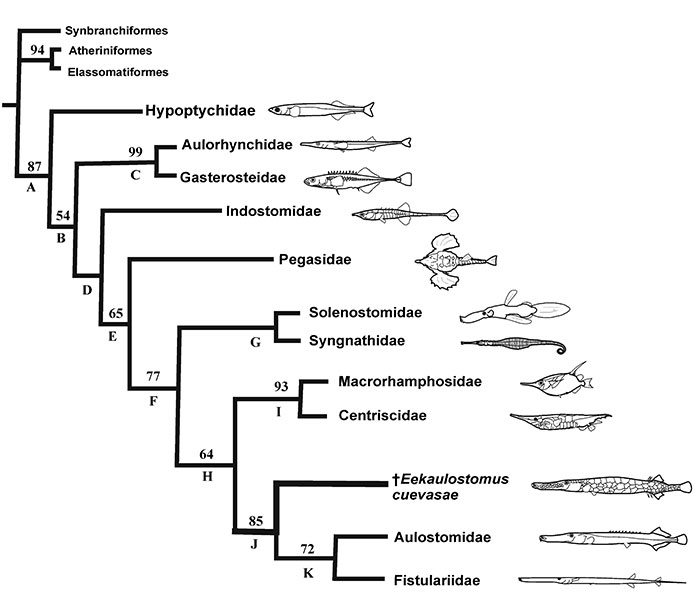
FIGURE 7. Geographical and temporal distribution of fossil and recent members of the superfamily Aulostomoidea, including Eekaulostomus cuevasae gen. and sp. nov. from the Belisario Domínguez quarry, near Palenque, Chiapas, southeastern Mexico.
 Kleyton Magno Cantalice. Departamento de Paleontología, Instituto de Geología, Universidad Nacional Autónoma de México; Circuito de la Investigación, Ciudad Universitaria, Coyoacán, Ciudad de México, 04510, México. kleytonbio@yahoo.com.br
Kleyton Magno Cantalice. Departamento de Paleontología, Instituto de Geología, Universidad Nacional Autónoma de México; Circuito de la Investigación, Ciudad Universitaria, Coyoacán, Ciudad de México, 04510, México. kleytonbio@yahoo.com.br
Kleyton Cantalice recently has obtained his Ph.D. in Biosciences at Universidade do Estado do Rio de Janeiro (UERJ). His field research is focused on taxonomy and systematic of fossil and extant acanthomorphs fishes to understand the origin and evolution of derived forms on salt and freshwater enviroments, principally along America. His studies are based on anatomical comparisons, phylogenetic systematic, biogeography and biochronology. Currently, he is a Postdoctoral student, working on a project about the phylogenetic and biogeographic implications of Paleocene acanthomorph fishes from Chiapas, Mexico.

 Jesús Alvarado-Ortega. Departamento de Paleontología, Instituto de Geología, Universidad Nacional Autónoma de México; Circuito de la Investigación, Ciudad Universitaria, Coyoacán, Ciudad de México, 04510, México. alvarado@geologia.unam.mx
Jesús Alvarado-Ortega. Departamento de Paleontología, Instituto de Geología, Universidad Nacional Autónoma de México; Circuito de la Investigación, Ciudad Universitaria, Coyoacán, Ciudad de México, 04510, México. alvarado@geologia.unam.mx
Born in 1966 in the city of Zacatecas. In 1998, I got the Bachelors degree (Biology) in the Sciences Faculty at Universidad Nacional Autónoma de México (UNAM). In 2005, I got my Doctoral degree in the Geological Institute (IGM), UNAM. During 2005-2007, I had a Posdoctoral position in the Biological Institute at Universidade do Estado do Rio de Janeiro, Brazil. Always I worked on Mexican fossil fishes under my advisor and friend Dr. Shelton P. Applegate. Since 2007, I´m in charge of the Tlayúa project as full-time Researcher at IGM, UNAM. The main interest of my work is focused on the taxonomical characterization of Mexican fossil vertebrates and the understood of phylogenetic and biogeographic patterns and process outlined by these fossils.
Eekaulostomus cuevasae gen. and sp. nov., an ancient armored trumpetfish (Aulostomoidea) from Danian (Paleocene) marine deposits of Belisario Domínguez, Chiapas, southeastern Mexico
Plain Language Abstract
The fossil fish Eekaulostomus cuevasae is named and described in this work. This fossil discovered in Paleocene marine strata of the Belisario Domínguez quarry, near Palenque town, Chiapas, Mexico. This fossil species is the oldest relative of the extant trumpetfish and cornetfish, and it is the first fossil representative of this group in America. The name of this fish refers to its most peculiar feature; its body is totally armored with star-shaped scutes.
Resumen en Español
Eekaulostomus cuevasae gen. and sp. nov., un antiguo pez trompeta acorazado (Aulostomoidea) de los depósitos marinos del Daniense (Paleoceno) de Belisario Domínguez, Chiapas, sureste de México
En este trabajo se describe e identifica a Eekaulostomus cuevasae gen. y sp. nov. como un miembro nuevo de la a superfamilia Aulostomoidea. El único ejemplar conocido de esta especie forma parte de una asociación fosilífera recientemente colectada en los sedimentos marinos del Paleoceno temprano pertenecientes a la unidad geológica Tenejapa-Lacandón, mismos que son explotados en la cantera Belisario Domínguez, en las cercanías del pueblo de Palenque, Chiapas, en el sureste de México. E. cuevasae representa el pez aulostomoido más antiguo hasta ahora conocido y el primer fósil de esta superfamilia encontrado en América. E. cuevasae se diferencia de otros aulostomoidos por presentar dos espinas antes de los radios blandos tanto en la aleta dorsal como en la anal, por tener todo el cuerpo y parte del hocico cubierto con escudos en forma de estrella, y también por el número relativamente elevado de radios principal en la aleta caudal. El reconocimiento de E. cuevasae como grupo basal de Aulostomoidea incrementa la distribución temporal y geográfica de esta superfamilia hasta el Daniano dentro de la región del Caribe, cuando gran parte de Chiapas permanecía bajo el mar después del evento de extinción masiva del límite Cretácico-Terciario. Este hallazgo también nos da evidencias que sugieren la pertenencia de Aulostomoidea dentro del orden Gasterosteiformes, en donde la presencia de los escudos cubriendo el cuerpo y de espinas en las aletas impares son características recurrentes.
Palabras clave: Nueva especie; Aulostomoidea; Fósil; Paleoceno; Chiapas; México
Traducción: authors
Resumo in Portuguese
No presente trabalho está a descrição de Eekaulostomus cuevasae gen. et sp. nov., identificada como um novo representante da superfamília Aulostomoidea. O único exemplar conhecido desta espécie faz parte de uma assembleia fóssil recém descoberta em sedimentos paleocênicos marinhos da unidade geológica Tenejapa-Lacandón, explorados na pedreira Belisario Domínguez, próximo à cidade de Palenque, no estado de Chiapas, localizada na região sudeste do México. E. cuevasae representa o mais antigo aulostomoideo conhecido e o primeiro registro fóssil da superfamília encontrado no continente Americano. Esta espécie se distingue dos demais aulostomoideos pela presença de dois espinhos anteriores aos raios das nadadeiras dorsal e anal; pela presença de cobertura corporal composta por escamas rígidas e em forma de estrela sobre todo o corpo e na parte anterior da cabeça; e por maior número de raios presentes na nadadeira caudal. O reconhecimento de E. cuevasae como um aulostomoideo basal aumenta a distribuição geográfica do grupo até o Caribe e temporal para a idade daniano, época em que grade parte de Chiapas estava submersa logo após o evento de extinção massiva do Cretáceo e Terciário. Estas descobertas também indicam fortes evidencias suportando a adesão ao aulostomoidos, dentro da ordem Gasterosteiformes, os quais a presença de cobertura corporal rígida e espinhos fortes nas nadadeiras impares são características recorrentes.
Translator: author
Résumé en Français
text
Translator: Kenny J. Travouillon or Antoine Souron
Deutsche Zusammenfassung
Eekaulostomus cuevasae gen. und sp. nov., ein ursprünglicher gepanzerter Trompetenfisch (Aulostomoidea) aus marinen Ablagerungen von Belisario Domínguez (Danian, Paläozän), Chiapas, südöstliches Mexiko
Eekaulostomus cuevasae gen. and sp. nov. ist hier beschrieben und identifiziert als ein neues Mitglied der Superfamilie Aulostomoidea. Das einzelne Stück dieser Art ist Teil einer neuen Fossilassemblage aus den marinen Sedimenten der frühpaläozänen geologischen Einheit Tenejapa-Lacandón, gefunden im Belisario Domínguez Steinbruch, nahe Palenque (Chiapas, südöstliches Mexiko). E. cuevasae repräsentiert den bis jetzt ältesten Aulostomoiden und die erste fossile Art dieser Superfamilie aus Amerika. E. cuevasae unterscheidet sich von anderen Aulostomoiden durch das Vorhandensein von zwei Stacheln vor den Weichstrahlen der Dorsal-und Afterflossen, den sternförmigen Schuppen auf dem gesamten Körper und auf Teilen der Schnauze und in der relativ großen Anzahl von Hauptstrahlen in der Schwanzflosse. Das Erkennen von E. cuevasae als Stammgruppe der Aulostomoidea vergrößert die zeitliche und geographische Verbreitung dieser Superfamilie bis in das Danium und innerhalb der Karibik, als ein großer Teil von Chiapas nach dem Kreide-Tertiär Massenaussterben unter Wasser lag. Dieser Fund legt ebenso die Zugehörigkeit der Aulostomoidea innerhalb der Ordnung Gasterosteiformes nahe, bei der die den Rumpf bedeckenden Schuppen und die robusten Stacheln der unpaaren Flossen ein wiederkehrendes Merkmal ist
Schlüsselwörter: Neue Art; Aulostomoidea; Fossil; Paläozän; Chiapas; Mexiko
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
Eekaulostomus cuevasae gen. and sp. nov., un antiguo pez trompeta acorazado (Aulostomoidea) de los depósitos marinos del Daniense (Paleoceno) de Belisario Domínguez, Chiapas, sureste de México
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024