

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
APPENDIX 1.
Cladistic matrix of Tanaka and Ichishima (2016) in nexus format.
Available in zipped format with Appendix 2 and Appendix 5.
APPENDIX 2.
Cladistic matrix of Tanaka and Ichishima (2016) in TNT format.
Available in zipped format with Appendix 1 and Appendix 5.
APPENDIX 3.
Morphological characters used in the phylogenetic analysis.
Terminology generally follows that of the cladistic papers cited, which in a few cases does not agree with the recommended uses of Mead and Fordyce (2009). For each character, references are given for the main past uses, with the relevant published character number given with a hatch # thus: Murakami et al. (2012a) #1.
Rostrum, Dental, and Mandibular
(1) Length of rostrum as percent skull length: moderately long, 50-55% (0); long, 55-60% (1); very long, >60% (2); medium, 50-40% (3); very short, 40-35% (4). (Murakami et al. (2012a, 2012b) #1; modified from Arnold and Heinsohn (1996) #8; Bianucci (2005) #1; Lambert (2008) #1; Tanaka and Fordyce (2014, 2015) #1).
(2) Premaxillae transverse proportion: transversely inflated almost entire length of rostrum (0); flat almost entire length of the rostrum (1). (Murakami et al. (2012a, 2012b) #2; Tanaka and Fordyce (2014, 2015) #2).
(3) Premaxillae mediolateral proportion: not compressed mediolaterally (0); compressed mediolaterally at anterior of rostrum (1). (Murakami et al. (2012a, 2012b) #3; Tanaka and Fordyce (2014, 2015) #3).
(4) Premaxillae at apex of rostrum: with lateral margins parallel or diverging (0); narrowing (1). (Murakami et al. (2012a, 2012b) #4; modified from Bianucci (2005) #2; Tanaka and Fordyce (2014, 2015) #4).
(5) Mesorostral groove constricted posteriorly, anterior to the nares and behind the level of the antorbital notch, then rapidly diverging anteriorly: absent (0); present (1). (modified from Murakami et al. (2012b) #279; Tanaka and Fordyce (2014, 2015) #7).
(6) Lateral margin of rostrum anterior to maxillary flange: concave (0); straight (1); convex (2); absent (3) (Murakami et al. (2012a, 2012b) #7; modified from Bianucci (2005) #3; Tanaka and Fordyce (2014, 2015) #8).
(7) Antorbital notch: absent or weakly developed (0); well developed (1). (Messenger and McGuire (1998) #1426; Fajardo-Mellor et al. (2006) #6; Murakami et al. (2012a, 2012b) #9; Tanaka and Fordyce (2014, 2015) #10).
(8) Width of premaxillae at mid-rostrum as percent greatest width of maxillae at level of postorbital processes: wide, >25% (0); medium, 25-15% (1); narrow, <15% (2). (Murakami et al. (2012a, 2012b) #10; modified from Aguirre-Fernandez et al. (2009) #4); Tanaka and Fordyce (2014, 2015) #11)
(9) Width of rostrum at mid-length as percent greatest width of maxillae at level of postorbital processes: wide, >35% (0); medium, 35-30% (1); narrow, <30% (2). (Murakami et al. (2012a, 2012b) #11; modified from Aguirre-Fernandez et al. (2009) #6; Tanaka and Fordyce (2014, 2015) #12).
(10) Anterior sinus fossa: absent (0); between anterior extremity of pterygoid sinus and posterior extremity of upper tooth row (1); between posterior extremity of upper tooth row and midpoint of rostrum (2); beyond midpoint of rostrum (3). (Murakami et al. (2012a, 2012b) #17; modified from Muizon (1988); Barnes (1990); Bianucci (2005) #13; Arnold and Heinsohn (1996) #21; Geisler and Sanders (2003) #157; Aguirre-Fernandez et al. (2009) #18; Geisler et al. (2011) (2012) #157; derived from Fraser and Purves (1960); Tanaka and Fordyce (2014, 2015) #18).
Teeth
(11) Teeth: conical, with or without accessory cusp (0); spatulate (1). (Murakami et al. (2012a, 2012b) #21; modified from Heyning (1989) #40 (1997) #72; Arnold and Heinsohn (1996) #25; Messenger and McGuire (1998) #1470; Geisler and Sanders (2003) #27 (2012) #27; Lambert (2008) #16; Geisler et al. (2011) #27; derived from Barnes (1984a) ; modified from Tanaka and Fordyce (2014, 2015) #21).
(12) Upper anterior "teeth": about same size as upper posterior teeth (0); clearly smaller than upper posterior teeth or absent (1). (Murakami et al. (2012a, 2012b) #22; modified from Tanaka and Fordyce (2014, 2015) #22).
(13) Greatest diameter of largest functional tooth as percent of greatest width of maxillae at the level of the postorbital processes: medium, 5-3% (0); small, <3% (1) (Murakami et al. (2012a, 2012b) #25; Aguirre-Fernandez et al. (2009) #15; modified from Tanaka and Fordyce (2014, 2015) #25).
Orbit
(14) Antorbital process shape in dorsal view: squared (0); rounded (1); tapered (2), reduced (3). (Bianucci (2005) #4; Murakami et al. (2012a, 2012b) #34; modified from Tanaka and Fordyce (2014, 2015) #34).
(15) Combined anteroposterior length of the lacrimal and jugal exposure that is posterior to antorbital notch: with skull in ventral view, exposure is small and combined length forms <50% of anteroposterior distance from antorbital notch to postorbital ridge (0); intermediate, forms between 50 and 62% of that distance (1); large, forms between 62 and 69% that distance (2); very large, forms >69% of that distance (3). (Murakami et al. in (2012a, 2012b) #42; modified from Geisler and Sanders (2003) #55; Geisler et al. (2012; 2011) #55; Tanaka and Fordyce (2014, 2015) #42).
(16) Dorsolateral edge of internal opening of infraorbital foramen: formed by maxilla (0); formed by maxilla and lacrimal and/or jugal (l); formed by lacrimal and/or jugal (2); formed by frontal (3). (Geisler and Sanders (2003) #57; Geisler et al. (2011) (2012) #57; Murakami et al. (2012a, 2012b) #43; derived from Miller (1923) ; Tanaka and Fordyce (2014, 2015) #43).
(17) Ventromedial edge of internal opening of infraorbital foramen: formed by maxilla (0); formed by maxilla and palatine and/or pterygoid (1); formed by palatine and/or pterygoid (2). (Geisler and Sanders (2003) #58; Geisler et al. (2012; 2011) #58; Murakami et al. (2012a, 2012b) #44; derived from Miller (1923); Tanaka and Fordyce (2014, 2015) #44).
(18) Direction of apex of postorbital process of frontal: projected posterolaterally and slightly ventrally (0); directed ventrally (1); not clear because of extremely reduced process (2). (modified from Murakami et al. (2012a, 2012b) #46; Geisler and Sanders (2003) #61; Geisler et al. (2012; 2011) #61; Tanaka and Fordyce (2014, 2015) #46).
(19) Shape of postorbital process of frontal: triangular, trapezoidal, or an anteroposteriorly widened falciform (0); dorsoventrally long falciform (1); robust, blunt descending posteriorly (2). (modified from Murakami et al. (2012a, 2012b) #47; Tanaka and Fordyce (2014, 2015) #47).
Facial Region
(20) Anterior dorsal infraorbital foramina: two (0); three or more (1). (Murakami et al. (2012a, 2012b) #49; modified from Barnes (1984b); Geisler and Sanders (2003) #64; Geisler et al. (2011) #64 (2012) #64; Tanaka and Fordyce (2014, 2015) #49).
(21) Width of premaxillae at antorbital notches as percent width of rostrum at antorbital notch: narrow, <49% (0); moderate, 50-64% (1); wide, >65% (2); antorbital notch absent (3). (Geisler and Sanders (2003) #66; Geisler et al. (2011)#66 (2012) #66; modified from Murakami et al. (2012a, 2012b) #51; Tanaka and Fordyce (2014, 2015) #51).
(22) Premaxillary foramen locating: medial (0); midpoint to lateral (1) absent (2). (modified from Murakami et al. (Murakami et al., 2014): Murakami et al. (2012b) #280; Tanaka and Fordyce (2014, 2015) #55).
(23) Lateral margin of the right premaxilla posterior to premaxillary foramen: widen posteriorly (0); straight (1). (Murakami et al. (2012b) #281; Tanaka and Fordyce (2014, 2015) #56).
(24) Posterolateral sulcus: deep (0); shallow or absent (1); presence of additional posterolateral sulcus (longitudinal striation) (2). (Murakami et al. (2012a, 2012b) #55; modified from Muizon (1984, 1988); Lambert (2008) #6; Geisler and Sanders (2003) #72; Geisler et al. (2011) (2012) #72; Tanaka and Fordyce (2014, 2015) #57).
(25) Posterior projections of premaxillae: both premaxillae extending posterior to anterior tip of nasals (0); neither premaxillae extending beyond external nares, and premaxillae displaced laterally by medial projection of maxilla (1); neither premaxillae extending posterior to external nares, and narrow posterior end of premaxillae adjacent to external nares (2); only right premaxilla extending beyond or in line with anterior-most portion of nasals (3). (Murakami et al. (2012a, 2012b) #76; modified from Muizon (1984); Barnes (1985a); Heyning (1989) #39, 42 (1997) #63, 71, 74; Arnold and Heinsohn,(1996) #35; Messenger and McGuire (1998) #1407, 1408; Fajardo-Mellor et al. (2006) #3; Lambert (2008) #5; Fordyce (1994) #27; Tanaka and Fordyce (2014, 2015) #58).
(26) Maxilla on dorsal surface of skull: does not contact supraoccipital posteriorly, maxilla separated by frontal and/or parietal (0); contact present (1). (Geisler and Sanders (2003) #129; Geisler et al. (2011) #129 (2012) #129, modified from Muizon (1991) (1994); Murakami et al. (2012a, 2012b) #60; Tanaka and Fordyce (2014, 2015) #62).
(27) Anterolateral corner of maxilla overlying supraorbital process of frontal: thin and equal in thickness to parts posteromedial (0); thickened with thinner maxilla in posteromedial direction (1). (Geisler and Sanders (2003) #78; Geisler et al. (2011) #78 (2012) #78; Murakami et al. (2012a, 2012b) #62; Tanaka and Fordyce (2014, 2015) #64).
(28) Maxillary crest on supraorbital process of maxilla: longitudinal ridges absent except at lateral edge of antorbital process (0); presence of longitudinal ridge except at lateral edge of antorbital process (1); longitudinal ridge present and joined with maxillary flange (2); presence of transversely compressed high crest, except at lateral edge of antorbital process (3); absent (4). (Murakami et al. (2012a, 2012b) #64; modified from Muizon (1984) (1987); Barnes (1985b); Messenger and McGuire (1998) #1420; Geisler and Sanders (2003) #79; Geisler et al. (2011) #79 (2012) #79; derived from Miller (1923); Tanaka and Fordyce (2014, 2015) #66).
(29) Fossa for inferior vestibule on maxilla lateral to external nares or lateral to premaxilla: absent (0); present (1). (Muizon (1988); Murakami et al. (2012a, 2012b) #68; derived from Curry (1992); Tanaka and Fordyce (2014, 2015) #70).
(30) Maxillary intrusion, anterior to external nares and encroaching the posteromedial or medial face of each premaxilla: absent (0); maxilla visible within opened mesorostral canal as small exposure medially (1); exposure of maxilla reaches dorsally to level of premaxilla and forms a square, rectangular to triangular plate (2); exposure of maxilla reaches dorsally and forms a small subcircular to polygonal ossicle (3). (Muizon (1984) (1988); Arnold and Heinsohn (1996) #24; Messenger and McGuire (1998) #1422; Murakami et al. (2012a, 2012b) #69; Tanaka and Fordyce (2014, 2015) #71).
(31) Premaxillary crest or posterior maxillary crest adjacent to nasal: absent (0); present (1). (transverse premaxillary crest, sensu Lambert (2005) #6; Murakami et al. (2012a, 2012b) #70; Tanaka and Fordyce (2014, 2015) #72).
(32) Premaxilla: not overhanging itself or maxilla laterally (0); overhanging itself or maxilla laterally, from anterior to midpoint of external nares (1). (Murakami et al. (2012a, 2012b) #71; Tanaka and Fordyce (2014, 2015) #73).
(33) Premaxillary sac fossa: smooth (0); rugose (1). (Messenger and McGuire (1998) #1551; Murakami et al. (2012a, 2012b) #72; Tanaka and Fordyce (2014, 2015) #74).
(34) Ratio of width of right premaxilla to width of left premaxilla in line with midpoint of external nares: 0.90-1.19 (0); 1.20-1.50 (1); 1.50> (2). (Murakami et al. (2012a, 2012b) #73; Tanaka and Fordyce (2014, 2015) #75).
(35) Ratio of greatest width of premaxillae to greatest width of maxillae at level of postorbital processes: ≥0.50 (0); 0.49-0.38 (1); <0.38 (2). (Murakami et al. (2012a, 2012b) #74; Tanaka and Fordyce (2014, 2015) #76).
(36) Premaxillary eminence: absent (0); present but low (1); present and high (2). (Lambert (2008) #4; Murakami et al. (2012a, 2012b) #75; modified from Muizon (1984); Barnes (1985a); Heyning (1989) #36 (1997) #68; Arnold and Heinsohn (1996) #12; Messenger and McGuire (1998) #1410; Geisler and Sanders (2003); #68; Fajardo-Mellor et al. (2006) #2; Geisler et al. (2011) #68 (2012) #69; derived from Flower (1867); Noble and Fraser (1971) ; Tanaka and Fordyce (2014, 2015) #77).
(37) Mesethmoid: not expanded posterodorsally (0); extended posterodorsally but narrow (1); expanded posterodorsally and visible in lateral view (2). (Murakami et al. (2012a, 2012b) #81; modified from Muizon (1984, 1988); Messenger and McGuire (1998) #1454; Bianucci (2005) #9; Tanaka and Fordyce (2014, 2015) #82).
Vertex and Area Adjacent to the Nares
(38) Nasals: lower than frontals (0); nearly same height as frontals (1); clearly higher than frontals (2). (Muizon (1988); Messenger and McGuire (1998) #1434; Geisler and Sanders (2003); #124; Geisler et al. (2011) #124 (2012) #124; Murakami et al. (2012a) #86; Tanaka and Fordyce (2014, 2015) #87).
(39) Nasal protuberance: absent (0); present (1). (Muizon (1988); Messenger and McGuire (1998) #1433; Fajardo-Mellor et al. (2006) #7; Lambert (2008) #8; Murakami et al. (2012a, 2012b) #87; Tanaka and Fordyce (2014, 2015) #88).
(40) Both nasals: straight anterior edges in one transverse plane (0); with point on midline and gap on each side between premaxilla and nasal (1); concave posteriorly on midline and gap on each side between premaxilla and nasal (2); concave posteriorly on midline (3). (Murakami et al. (2012a, 2012b) #88; modified from Geisler and Sanders (2003) #116; Geisler et al. (2011) #116 (2012) #116; derived from Moore (1968); Tanaka and Fordyce (2014, 2015) #89).
(41) Lateral edges of nasals: not overhanging or covering maxillae or premaxillae (0); overhanging or partly covering maxillae or premaxillae (1). (Murakami et al. (2012a, 2012b) #92; Tanaka and Fordyce (2014, 2015) #93).
(42) Nasal- frontal suture: anterior wedge (narial process) between frontal posterior ends of nasals (0); W or reversed U suture line (1). (Murakami et al. (2012a, 2012b) #93; modified from Muizon (1988); Geisler and Sanders (2003) #121; Geisler et al. (2011) #121 (2012) #121; Tanaka and Fordyce (2014, 2015) #94).
(43) Frontals posterior to nasals and between premaxillae: narrower than transverse width of nasals, maxillae expanded medially posterior to nasals (0); same as transverse width of nasals (1); wider than maximum transverse width across nasals (2). (Geisler and Sanders (2003) #125; Geisler et al. (2011) #125 (2012) #125; Murakami et al. (2012a, 2012b) #94; modified from Messenger and McGuire (1998) #1457; Tanaka and Fordyce (2014, 2015) #95).
(44) Frontal boss on vertex: absent (0); present (1). (Muizon (1984, 1988); Messenger and McGuire (1998) #1461; Fajardo-Mellor et al. (2006) #12; Murakami et al. (2012a, 2012b) #95; modified from Lambert (2008) #9; Tanaka and Fordyce (2014, 2015) #96).
(45) Nuchal crest: below frontals and/or nasals (0); at same level as frontals and/or nasals (1). (Murakami et al. (2012a, 2012b) #99; modified from Geisler and Sanders (2003) #128; derived from Moore (1968); Tanaka and Fordyce (2014, 2015) #100).
Temporal Fossae, Zygomatic Arch, and Occipitals
(46) Temporal fossa shape in lateral view: height lower than anteroposterior length (0); higher (1); lower and its posterior end is rounded (2). (Tanaka and Fordyce (2014, 2015) #281)
(47) Temporal fossa: not roofed over by lateral expansion of maxillae (0); roofed over by lateral expansion of maxillae (1). (Muizon (1988); Heyning (1989) #22 (1997) #54; Arnold and Heinsohn (1996) #39; Messenger and McGuire (1998) #1453; Murakami et al. (2012a, 2012b) #100; Tanaka and Fordyce (2014, 2015) #101).
(48) Parietals in dorsal view: completely absent in skull roof (0); visible only as triangular areas, dorsolateral to supraoccipital, with non-overlapping supraoccipital separated from and contacting parietals along irregular suture (1). (Geisler and Sanders (2003) #134; Geisler et al. (2011) #134 (2012) #134; Murakami et al. (2012a, 2012b) #104; derived from Whitmore and Sanders (1977); Barnes (1990); modified from Lambert (2005) #15; Tanaka and Fordyce (2014, 2015) #105).
(49) Interparietal: present (0); absent or fused and therefore not distinguishable from parietals and frontals (1). (Geisler and Sanders (2003) #135; Geisler et al. (2011) #135 (2012) #135; Murakami et al. (2012a) #105; Tanaka and Fordyce (2014, 2015) #106).
(50) Anterior zygomatic process end of squmosal in lateral view: taipered (0); squared (1). (Tanaka and Fordyce (2014, 2015) #282)
(51) Zygomatic process of squamosal: directed anterolaterally (0); directed anteriorly (1). (Sanders and Barnes, 2002; Geisler and Sanders (2003) #142; Geisler et al. (2011) #142 (2012) #142; Murakami et al. (2012a, 2012b) #108; Tanaka and Fordyce (2014, 2015) #109).
(52) Zygomatic process of squamosal in lateral view: part of dorsal face visible (0); entire dorsal surface of squamosal visible (1). (Murakami et al. (2012a, 2012b) #109; Tanaka and Fordyce (2014, 2015) #110).
(53) Emargination of posterior edge of zygomatic process by neck muscle fossa, skull in lateral view: deep emargination (0); shallow emargination (1). (Geisler and Sanders (2003) #144; Geisler et al. (2011) #144 (2012) #144; Murakami et al. (2012a, 2012b) #110; Tanaka and Fordyce (2014, 2015) #111).
(54) Ventral edge of zygomatic process of squamosal in lateral view: concave (0); almost straight (1); convex (2). (Geisler and Sanders (2003); #150; Geisler et al. (2011) #150 (2012) #150; Murakami et al. (2012a, 2012b) #112).
(55) Postglenoid process of squamosal: not reduced (0); greatly reduced (1). (Murakami et al. (2012a, 2012b) #113; Tanaka and Fordyce (2014, 2015) #114).
(56) Postglenoid process in lateral view: tapering ventrally (0); squared off ventrally (1); same as state 1 except very wide anteroposterior diameter of process (2). (Geisler and Sanders (2003) #151; Lambert (2005) #24; Geisler et al. (2011) #151 (2012) #151; Murakami et al. (2012a, 2012b) #114; derived from Muizon (1991) ; Tanaka and Fordyce (2014, 2015) #115).
(57) Relative ventral projections of postglenoid and post-tympanic processes of squamosal: postglenoid process more ventral or at same level as post-tympanic process (0); apex of postglenoid process dorsally higher than post-tympanic process (1). (Lambert (2005) #25; Murakami et al. (2012a). b #115; Tanaka and Fordyce (2014, 2015) #116).
(58) Dorsal condyloid fossa: present, situated anterodorsal to dorsal edge of condyle (0); present and forming deep pit (1). (Geisler and Sanders (2003) #156; Geisler et al. (2011) #156 (2012) #156; Murakami et al. (2012a, 2012b) #118; derived from Sanders and Barnes (2002); Tanaka and Fordyce (2014, 2015) #119).
Anterior Basicranium
(59) Lateral lamina of palatine relationship with orbit: does not form bony bridge “over” (= ventral to) orbit (0); does form bony bridge “over” (= ventral to) orbit (1). (Muizon (1984); Messenger and McGuire (1998) #1444; Murakami et al. (2012a, 2012b) #123; Tanaka and Fordyce (2014, 2015) #124).
(60) Pterygoids in anteroventral view: separated from each other by posteroventrally elongated palatines and/or vomer (0); contacting entire length of hamular process (1); contacting each other partially (2). (Murakami et al. (2012a, 2012b) #124; modified from Arnold and Heinsohn (1996) #5; Messenger and McGuire (1998) #1445; Fajardo-Mellor et al. (2006) #9; derived from Flower (1884); Barnes (1985a); Marsh et al. (1989) ; Tanaka and Fordyce (2014, 2015) #125).
(61) Lateral lamina of pterygoid: present and articulated with alisphenoid (0); partial, restricted to region lateral to hamular process (1). (Murakami et al. (2012a) #126; modified from Arnold and Heinsohn (1996) #121; Messenger and McGuire (1998) #1446; Geisler and Sanders (2003) #164; Lambert (2005) #32; Geisler et al. (2011) #164 (2012) #164; derived from Miller (1923); Kellogg (1936); Fraser and Purves (1960) ; Tanaka and Fordyce (2014, 2015) #127).
(62) Subtemporal crest: present (0); present but reduced, or absent (1). (modified from Geisler and Sanders (2003) #165; Geisler et al. (2011) #165 (2012) #165; Murakami et al. (2012a, 2012b) #127; Tanaka and Fordyce (2014, 2015) #128).
(63) Superior lamina of pterygoid: absent from sphenoidal region but present in orbital region (0); present and covers most of ventral exposure of alisphenoid (1); partially absent from orbital region (2); completely absent from orbital region (3). (Murakami et al. (2012a, 2012b) #128; modified from Arnold and Heinsohn (1996) #16; Geisler and Sanders (2003) #167; Geisler et al. (2011) #167 (2012) #167; derived from Miller (1923); Fraser and Purves (1960)) ; Tanaka and Fordyce (2014, 2015) #129.
(64) Depth of pterygoid sinus fossa in basicranium: deep, and extended dorsally into orbit (0); deep, excavated dorsally to level of cranial foramen oval (1). (modified from Fordyce (1994) #6; Lambert (2005) #30; Murakami et al. (2012a, 2012b) #130; Tanaka and Fordyce (2014, 2015) #131).
(65) Anterior level of pterygoid sinus fossa: interrupted posterior to, or the level of, antorbital notch (0); extending beyond the level of the antorbital notch (1). (Lambert (2005) #29; Murakami et al. (2012a, 2012b) #131; Tanaka and Fordyce (2014, 2015) #132).
(66) Fossa for preorbital lobe of pterygoid sinus in orbit: absent (0); present (1). (Fraser and Purves (1960); Arnold and Heinsohn (1996) #18; Murakami et al. (2012a, 2012b) #133; Tanaka and Fordyce (2014, 2015) #134).
(67) Dorsal development of fossa for preorbital lobe of pterygoid sinus toward the frontal-maxilla suture: absent (0); present (1). (Muizon (1984, 1988); Heyning (1989) #37 (1997) #69; Messenger and McGuire (1998) #1460; Arnold and Heinsohn (1996) #20; Lambert (2008) #13; Murakami et al. (2012a, 2012b) #134; modified from Fajardo-Mellor et al. (2006) #13; derived from Fraser and Purves (1960) ; Tanaka and Fordyce (2014, 2015) #135).
(68) Postorbital lobe of pterygoid sinus fossa: absent (0); present (1); large and deep (2). (Arnold and Heinsohn (1996) #18; Geisler and Sanders (2003) #170; Geisler et al. (2011) #170 (2012) #170; Murakami et al. (2012a, 2012b) #135; derived from Fraser and Purves (1960) ; Tanaka and Fordyce (2014, 2015) #136).
(69) Anteroposteriorly elongated pterygoid sinus fossa, at level of orbit, bordered by mediolaterally compressed subtemporal crest of frontal: absent (0); present (1). (Murakami et al. (2012a, 2012b) #136; Tanaka and Fordyce (2014, 2015) #137).
(70) Orbitosphenoid: not contacting lacrimal or lacrimojugal (0); contacting lacrimal or lacrimojugal (1). (Murakami et al. (2012a, 2012b) #137; Tanaka and Fordyce (2014, 2015) #138).
(71) Ratio of length of hamular process of pterygoid to cranium length: <0.30 (0); 0.30-0.44 (1); 0.45-0.59 (2); >0.60 (3). The length of the hamular process of the pterygoid is measured from anterior edge of the pterygoid to posterior edge of the hamular process. The cranium length is measured from anterior edge of the antorbital process to posterior edge of occipital condyles. (Murakami et al. (2012a, 2012b) #138; modified from Heyning (1989) #18 (1997) #50; Muizon (1991); Messenger and McGuire (1998) #1447; Lambert (2005) #31; Tanaka and Fordyce (2014, 2015) #139).
(72) Keel affecting ventral surfaces of hamular processes: absent (0); present (1). (Muizon (1988); Messenger and McGuire (1998) #1449; Bianucci (2005) #14; Murakami et al. (2012a, 2012b) #139; modified from Fajardo-Mellor et al. (2006) #10; Tanaka and Fordyce (2014, 2015) #140).
(73) Exposure of medial lamina of pterygoid hamuli in lateral view: complete or broad exposure due to extreme reduction of lateral lamina of pterygoid hamuli (0); no exposure due to a posterior extension of lateral lamina extending posterior to medial lamina (1); medial lamina of pterygoid hamuli exposing lateral lamina through ovoid window in lateral view (2). (Muizon (1988); Fajardo-Mellor et al. (2006) #11; Murakami et al. (2012a, 2012b) #140; derived from Noble and Fraser (1971); Tanaka and Fordyce (2014, 2015) #141).
(74) Shape of restricted area between postorbital ridge of frontal and subtemporal crest from ventral view: anteroposteriorly long elliptical (0); wide fan-shape (1); narrow fan-shape (2), rhombus (3). (Tanaka and Fordyce (2014, 2015) #280)
Posterior Basicranium
(75) Tympanosquamosal recess: very large, forming large fossa bordering entire medial edge of glenoid fossa (0); present and enlarged, forming triangular fossa medial and anteromedial to postglenoid process (1). (Geisler and Sanders (2003) #178; Geisler et al. (2011) #178 (2012) #178; Murakami et al. (2012a, 2012b) #143; modified from Lambert (2005) #35; derived from Fraser and Purves (1960), and Fordyce (2002); Tanaka and Fordyce (2014, 2015) #144).
(76) Fossa for the basisphenoidal sinus: absent (0); present (1). (Fraser and Purves (1960); Mead and Fordyce (2009); Murakami et al. (2012a, 2012b) #145; Tanaka and Fordyce (2014, 2015) #146).
(77) Posterior portion of periotic fossa of squamosal: fossa absent (0); fossa present but shallow (1); posteromedial portion contains large deep fossa (2). (Geisler and Sanders (2003) #187; Geisler et al. (2011) #187 (2012) #187; Murakami et al. (2012a, 2012b) #149 and #151; Tanaka and Fordyce (2014, 2015) #151).
(78) Length of zygomatic process of squamosal as percent of greatest width of maxilla at postorbital process: >31% (0); ≤30% (1). (Murakami et al. (2012a, 2012b) #152; modified from Heyning (1989) #33, 35, #65, 67; Geisler and Sanders (2003) #188; Geisler et al. (2011) #188 (2012) #188; Tanaka and Fordyce (2014, 2015) #152)
(79) Fossa for posterior sinus in exoccipital: absent or slightly concave (0); moderately concave (1); forming deep sack-like structure (2). (Murakami et al. (2012a, 2012b) #161; modified from Muizon (1991); Lambert (2005) #38; Tanaka and Fordyce (2014, 2015) #161).
(80) Occipital condyles; on pedicle (0); lacking pedicle, unified with occipital (1). (Tanaka and Fordyce (2014, 2015) #284)
Periotic
(81) Relative position of dorsal depth of stapedial muscle fossa and fenestra rotunda: ventral to, or in line with, dorsal edge of fenestra rotunda (0); well dorsal to fenestra rotunda (1). (Geisler and Sanders (2003) #223; Geisler et al. (2011) #223 (2012) #223; Murakami et al. (2012a, 2012b) #177; Tanaka and Fordyce (2014, 2015) #176).
(82) Aperture for cochlear aqueduct: smaller than aperture for vestibular aqueduct (0); approximately same size as aperture for vestibular aqueduct (1); much larger than aperture for vestibular aqueduct, with narrow posterior edge (2). (Geisler and Sanders (2003) #227; Geisler et al. (2011) #227 (2012) #227; Murakami et al. (2012a, 2012b) #181; modified from Muizon (1987); Fordyce (1994); Lambert (2005) #52; Tanaka and Fordyce (2014, 2015) #180).
(83) Bony connection between posterior process of periotic and squamosal/occipital bones: present (0); absent (ligamentous). (1). (Muizon (1984); Arnold and Heinsohn (1996) #34; Messenger and McGuire (1998) #1491; Murakami et al. (2012a, 2012b) #188; derived from Fraser and Purves (1960); Kasuya (1973); Heyning (1989) ; Tanaka and Fordyce (2014, 2015) #187).
(84) Posterior process of periotic in lateral view: ventrally bent (0); in same plane as body of periotic (1). (Bianucci (2005) #19; Murakami et al. (2012a, 2012b) #189; modified from Arnold and Heinsohn (1996) #28; Lambert (2005) #54; Tanaka and Fordyce (2014, 2015) #188).
(85) Angle between posterior process of periotic and long axis of pars cochlearis from dorsal or ventral views: >135° (0); ≤135° (1). (Murakami et al. (2012a, 2012b) #190; modified from Geisler and Sanders (2003) #246; Lambert (2005) #54; Geisler et al. (2011) #246 (2012) #246; derived from Kasuya (1973); Barnes (1990); Luo and Marsh (1996); Tanaka and Fordyce (2014, 2015) #189).
(86) Length of posterior process of periotic as percent length of pars cochlearis: long, ≥85% (0); short, ≤84% (1). (Murakami et al. (2012a, 2012b) #193; modified from Barnes (1990); Luo and Marsh (1996) #24; Geisler and Sanders (2003) #245; Geisler et al. (2011) #245 (2012) #245; Tanaka and Fordyce (2014, 2015) #193).
Tympanic Bulla
(87) Articulation of posterior process of tympanic bulla with squamosal: process contacting post-tympanic process of squamosal and posterior process of periotic (0); process contacting periotic only (1). (Muizon (1984); Fordyce (1994) #29; Arnold and Heinsohn (1996) #34; Messenger and McGuire (1998) #1481; Lambert (2005) #56; Murakami et al. (2012a, 2012b) #197; derived Kasuya (1973) ; Tanaka and Fordyce (2014, 2015) #197).
(88) Width of tympanic bulla as percentage of its length along its long axis: wide, ≥65% (0); narrow and long, ≤64% (1). (Geisler and Sanders (2003) #251; Bianucci (2005) #23; Geisler et al. (2011) #251 (2012) #251; Murakami et al. (2012a, 2012b) #198; derived from Kasuya (1973); Tanaka and Fordyce (2014, 2015) #198).
(89) Lateral furrow of tympanic bulla: shallow groove (0); absent (1); deep, well-defined groove (2). (Murakami et al. (2012a, 2012b) #200; modified from Muizon (1984, 1988); Arnold and Heinsohn(1996) #31; Messenger and McGuire (1998) #1485; Fajardo-Mellor et al. (2006) #17; Lambert (2008) #17; derived from Kasuya (1973) ; Tanaka and Fordyce (2014, 2015) #200).
(90) Dorsomedial edge of sigmoid process: expanded anteriorly to oppose lateral tuberosity of periotic (0); not articulating with squamosal or periotic (1) (Murakami et al. (2012a, 2012b) #202; modified from Geisler and Sanders (2003) #260; Geisler et al. (2011) #260 (2012) #260; modified from Luo and Marsh (1996) #10; Tanaka and Fordyce (2014, 2015) #202).
(91) Elliptical foramen of tympanic bulla: present (0); absent or close (1). (Geisler and Sanders (2003) #261; Geisler et al. (2011) #261 (2012) #261; Murakami et al. (2012a, 2012b) #204; derived from Kasuya (1973); Tanaka and Fordyce (2014, 2015) #204).
(92) Surface of posterior process of tympanic bulla: spiny or irregular edges (0); rounded and pachyostotic (1). (Muizon (1991); Messenger and McGuire (1998) #1483; Murakami et al. (2012a, 2012b) #206; derived from Kasuya (1973); Tanaka and Fordyce (2014, 2015) #206).
(93) Posterior edge of medial prominence of involucrum: approximately in line with posterior edge of lateral prominence (0); distinctly anterior to posterior edge of lateral prominence (1). (Muizon (1987); Geisler and Sanders (2003) #269; Geisler et al. (2011) #269 (2012) #269; Murakami et al. (2012a, 2012b) #209; derived from Kasuya (1973); Tanaka and Fordyce (2014, 2015) #209).
(94) Posterior end of ventromedial keel: not protruding and directed medially (0); protruding and directed medially (1). (Geisler and Sanders (2003) #275; Geisler et al. (2011) #275 (2012) #275; Murakami et al. (2012a, 2012b) #214; Tanaka and Fordyce (2014, 2015) #213).
Hyals
(95) Basihyal and thyrohyal shape: arched (0); angled (1). (Murakami et al. (2012a, 2012b) #216; modified from Bianucci (2005) #25; Tanaka and Fordyce (2014, 2015) #215).
Vertebrae
(96) Dorsal transverse process of atlas: developed dorsolaterally (0); absent or rudimentary obtuse angle (1). (Murakami et al. (2012a, 2012b) #217; modified from Muizon (1988); Barnes (1990); Tanaka and Fordyce (2014, 2015) #216).
(97) Roof of neural canal of atlas: arched (0); straight (1). (Murakami et al. (2012a, 2012b) #218; Tanaka and Fordyce (2014, 2015) #217).
(98) Postzygapophysis of axis in anterior view: appearing as crest, elongated dorsolaterally (0); appearing as rudimentary crest (1); not appearing (2). (Murakami et al. (2012a, 2012b) #219; Tanaka and Fordyce (2014, 2015) #218).
(99) Cervical vertebrae: unfused (0); atlas and axis fused (1); C1-C3 or C1-C4 fused (2); C1-C6 or C1-C7 fused (3); C2-C7 fused (4). (Murakami et al. (2012a, 2012b) #220; modified from Arnold and Heinsohn (1996) #9; Messenger and McGuire (1998) #1501; Geisler and Sanders (2003) #278, 279; Fajardo-Mellor et al. (2006) #18; Lambert (2008) #18; Geisler et al. (2011) #278, 279 (2012) #278, 279; derived from Allen (1923); Miller (1923); Fraser and Noble (1971); De Smet (1977); Rommel (1990) ; Tanaka and Fordyce (2014, 2015) #219).
(100) Length of cervicals (C1-C7) as percent of height of vertebral body plus neural canal of atlas: long, >150% (0); short, <150% (1). (Murakami et al. (2012a, 2012b) #221; Tanaka and Fordyce (2014, 2015) #220).
(101) Transverse processes of lumbar vertebrae: extend parallel to anterior and posterior borders (0); triangular (1). (Muizon (1984, 1985, 1988); Messenger and McGuire (1998) #1502; Geisler and Sanders (2003) #285; Geisler et al. (2011) #285 (2012) #285; Murakami et al. (2012a, 2012b) #224; Tanaka and Fordyce (2014, 2015) #223).
Scapula
(102) Anterodorsal part of scapula: rounded (0); almost rectilinear (1). (Murakami et al. (2012a, 2012b) #232; Tanaka and Fordyce (2014, 2015) #232).
(103) Ventral projection on anterior border of scapula: absent (0); present (1). (Fajardo-Mellor et al. (2006) #26; Murakami et al. (2012a, 2012b) #233; derived from Noble and Fraser (1971); Tanaka and Fordyce (2014, 2015) #233).
(104) Coracoid process of scapula: not expanded distally (0); expanded distally (1); notably reduced or absent (2). (Murakami et al. (2012a, 2012b) #237; modified from Muizon (1987, 1994); Messenger and McGuire (1998) #1504; Geisler and Sanders (2003) #292; Lambert (2005) #73; Bianucci (2005) #33; Geisler et al. (2011) #292; derived from True (1904); Tanaka and Fordyce (2014, 2015) #237).
(105) Coracoid process of scapula, with glenoid fossa: directed horizontally (0); directed anteroventrally (1). (modified from Murakami et al. (2012a, 2012b) #238; modified Barnes (1990); Tanaka and Fordyce (2014, 2015) #238).
(106) Acromion of scapula: narrow and not expanded distally (0); expanded distally (1). (Murakami et al. (2012a, 2012b) #239; modified from Bianucci (2005) #34; Tanaka and Fordyce (2014, 2015) #239).
(107) Acromion of scapula, when glenoid fossa direct ventrally: directed horizontally (0); directed anterodorsally (1); directed anteroventrally (2). (Murakami et al. (2012a, 2012b) #240; modified from Barnes (1990); Tanaka and Fordyce (2014, 2015) #240).
Forelimb (except scapula)
(108) Ratio of length of humerus to length of radius: long, >1.1 (0); short, <0.8 (1). (Murakami et al. (2012a, 2012b) #242; modified from Sanders and Barnes (2002); Geisler and Sanders (2003) #297; Geisler et al. (2012; 2011) #297; Tanaka and Fordyce (2014, 2015) #242).
(109) Prominent deltoid crest on anterior edge of humerus: present, forms greatest anteroposterior diameter along shaft (0); forming a knob-like tuberosity (1); tuberosity or crest absent (2). (Geisler and Sanders (2003) #294; Geisler et al. (2011) #294 (2012) #294; Murakami et al. (2012a, 2012b) #244; derived from Sanders and Barnes (2002); Tanaka and Fordyce (2014, 2015) #244).
(110) Radial and ulnar facets of humerus in lateral view: facets forming a semicircular articulation surface (0); facets forming an obtuse angle (1). (Barnes, Barnes (1990); Geisler and Sanders (2003) #296; Geisler et al. (2011) #296 (2012) #296; Murakami et al. (2012a, 2012b) #245; Tanaka and Fordyce (2014, 2015) #245).
(111) Olecranon process: present as a distinct process (0); present as a slightly raised proximal posterior edge (1); absent (2). (Messenger and McGuire (1998) #1507; Geisler and Sanders (2003) #296; Geisler et al. (2011) #284 (2012) #284; Murakami et al. (2012a, 2012b) #246; modified from Muizon (1984); Barnes (1990); Arnold and Heinsohn (1996) #10; Fajardo-Mellor et al. (2006) #28; derived from Howell (1927); Bianucci (2005) #37; Tanaka and Fordyce (2014, 2015) #246).
Added characters to Tanaka and Fordyce (2015)
(112) Mandible fossa of squamosal: wider than squamosal reces (0); narrower than squamosal recess (1).
(113) Anterior end of zygomatic process in ventral view: rounded (0); tapered (1).
(114) Most anterior maxillary foramen: anterior to or in line with antorbital notch (0); posterior to antorbital notch (1). (Ichishima and Kimura (2005); Colpaert et al. (2015) #3)
(115) Longitudinal sulcus through premaxillary eminence: 0, absent; 1, present. This additional sulcus is thought to be present in Brabocetus, at least on right side. (de Muizon (1984); Colpaert et al. (2015) #6)
(116) Dorsal extension of pterygoid sinus fossa between frontal and maxilla: absent or poorly developed (0); deep (1). (Lambert (2008); Fajardo‐Mellor et al. (2006); Colpaert et al. (2015) #13)
(117) Number of teeth in each upper tooth row: tooth counts >35 (0); tooth counts between 35 and 30 (1); minimum tooth counts 18-24 and maximum tooth counts 26-29 (2); minimum tooth counts 8-17 and maximum tooth counts 20-25 (3). (Fajardo‐Mellor et al. (2006); Lambert (2008); Colpaert et al. (2015) #15)
(118) Spinous process of axis: short, extends posteriorly only to about C4 (0); long, nearly contacts spinous process of C7 (1). (Fajardo‐Mellor et al. (2006); Colpaert et al. (2015) #19)
(119) Transverse foramina (vertebrarterial canal) of C4: complete (0); incomplete or absent (1). (Fajardo‐Mellor et al. (2006); Colpaert et al. (2015) #20)
(120) Transverse process of axis: well developed (0); weakly developed or absent (1). (Fajardo‐Mellor et al. (2006); Lambert (2008); Colpaert et al. (2015) #21)
(121) Metapophyses of spinous processes: well developed (0); absent or weakly developed (1). (Fajardo‐Mellor et al. (2006); Colpaert et al. (2015) #22)
(122) Acromion and coracoid processes of scapula: acromion longer than coracoid (0); acromion shorter than coracoid (1). (Fajardo‐Mellor et al. (2006);Colpaert et al. (2015) #23)
(123) Supraspinous fossa: broad (0); narrow (1). (Fajardo‐Mellor et al. (2006); Colpaert et al. (2015) #24)
REFERENCES
Aguirre-Fernández, G., Barnes, L.G., Aranda-Manteca, F.J., and Fernández-Rivera, J.R. 2009. Protoglobicephala mexicana, a new genus and species of Pliocene fossil dolphin (Cetacea; Odontoceti; Delphinidae) from the Gulf of California, Mexico. Boletin de la Sociedad Geologica Mexicana, 61(2):245-265.
Allen, G.M. 1923. The black finless porpoise, Meomeris. Bulletin of the Museum of Comparative Zoology, 65(7):233-256.
Arnold, P.W., and Heinsohn, G.E. 1996. Phylogenetic status of the Irrawaddy dolphin Orcaella brevirostris (Owen in Gray): a cladistic analysis. Memoirs of the Queensland Museum, 39(2):141-204.
Barnes, L.G. 1984a. Fossil odontocetes (Mammalia: Cetacea) from the Almejas Formation, Isla Cedros, Mexico. Paleobios, 42:1-46.
Barnes, L.G. 1984b. Whales, dolphins and porpoises; origin and evolution of the Cetacea. In Gingerich, P.D., and Badgle, C.E. (eds.), Mammals. Notes for a short course, University of Atennessee, Department of Geological Science.
Barnes, L.G. 1985a. Evolution, taxonomy and antitropical distributions of the porpoises (Phocoenidae, Mammalia). Marine Mammal Science, 1(2):149-165.
Barnes, L.G. 1985b. Fossil pontoporiid dolphins (Mammalia: Cetacea) from the Pacific coast of North America. Contributions to Science, Natural History Museum of Los Angeles County, 363:1-34.
Barnes, L.G. 1990. The fossil record and evolutionary relationships of the genus Tursiops, p. 3-26. In Leatherwood, S., and Reeves, R.R. (eds.), The bottlenose dolphin. Academic Press Inc, San Diego, New York.
Bianucci, G. 2005. Arimidelphis sorbinii a new small killer whale-like dolphin from the Pliocene of Marecchia River (Central eastern Italy) and a phylogenetic analysis of the Orcininae (Cetacea: Odontoceti). Rivista Italiana di Paleontologia e Stratigrafia, 111(2):329-344.
Colpaert, W., Bosselaers, M., and Lambert, O. 2015. Out of the Pacific: a second fossil porpoise from the Pliocene of the North Sea Basin. Acta Paleontologica Polonica, 60(1):1-10.
Curry, B.E. 1992. Facial anatomy and potential function of facial structures for sound production in the harbor porpoise ( Phocoena phocoena ) and Dall's porpoise ( Phocoenoides dalli ). Canadian Journal of Zoology, 70(11):2103-2114.
de Muizon, C. 1984. Les vertébrés fossiles de la Formation Pisco (Pérou). deuxiéme partie: les Odontocétes (Cetacea, Mammalia) du Pliocéne inférieur de Sud-Sacaco. Travaux de l’Institut Français d’Études Andines, 27:1-188.
de Muizon, C. 1985. Nouvelles données sur le diphylétisme des Dauphins de rivière (Odontoceti, Cetacea, Mammalia). Comptes rendus l'Academie des Sciences series 2,, 301:359-362.
de Muizon, C. 1987. The affinities of Notocetus vanbenedeni, an Early Miocene platanistoid (Cetacea, Mammalia) from Patagonia, southern Argentina. American Museum novitates, 2904:1-27.
de Muizon, C. 1988. Les relations phylogenetiques des Delphinida (Cetacea, mammalia). Annales de paléontologie, 74(4):159-227.
de Muizon, C. 1991. A new Ziphiidae (Cetacea) from the Early Miocene of Washington State (USA) and phylogenetic analysis of the major groups of odontocetes. Bulletin du Muséum National d'Histoire Naturelle, 12(3-4):279-326.
de Muizon, C. 1994. Are the squalodonts related to the platanistoids? Proceedings of the San Diego Society of Natural History, 29:135-146.
De Smet, W.M.A. 1977. The regions of the cetacean vertebral column volume 3, p. 59-80. In Harrison, R.J. (ed.), Functional anatomy of marine mammals. Academic Press, London.
Fajardo‐Mellor, L., Berta, A., Brownell, R.L., Boy, C.C., and Goodall, N.P. 2006. The phylogenetic relationships and biogeography of true porpoises (Mammalia: Phocoenidae) based on morphological data. Marine Mammal Science, 22(4):910-932.
Flower, W.H. 1867. Description of the skeleton of Inia geoffrensis and the skull of Pontoporia blainvillii, with remarks on the systematic position of these animals in the Order Cetacea. Transactions of the Zoological Society of London, 6(3):87-116.
Flower, W.H. 1884. On the characters and divisions of the Family Delphinidae. Proceedings of the Zoological Society of London, 1883:466-513.
Fordyce, R.E. 1994. Waipatia maerewhenua , new genus and new species (Waipatiidae, new family), an archaic Late Oligocene dolphin (Cetacea: Odontoceti: Platanistoidea) from New Zealand. Proceedings of the San Diego Society of Natural History, 29:147-176.
Fordyce, R.E. 2002. Simocetus rayi (Odontoceti: Simocetidae, new family): A bizarre new archaic Oligocene dolphin from the eastern North Pacific. Smithsonian Contributions to Paleobiology, 93:185-222.
Fraser, F.C., and Purves, P.E. 1960. Hearing in cetaceans: evolution of the accessory air sacs and the structure of the outer and middle ear in recent cetaceans. Bulletin of the British Museum of Natural History (Zoology) 7:1-140.
Geisler, J.H., Godfrey, S.J., and Lambert, O. 2012. A new genus and species of late Miocene inioid (Cetacea, Odontoceti) from the Meherrin River, North Carolina, USA. Journal of Vertebrate Paleontology, 32(1):198-211.
Geisler, J.H., McGowen, M.R., Yang, G., and Gatesy, J. 2011. A supermatrix analysis of genomic, morphological, and paleontological data from crown Cetacea. BMC Evolutionary Biology, 11:1-33.
Geisler, J.H., and Sanders, A.E. 2003. Morphological evidence for the phylogeny of Cetacea. Journal of Mammalian Evolution, 10(1/2):23-129.
Heyning, J.E. 1989. Comparative facial anatomy of beaked whales (Ziphiidae) and a systematic revision among the families of extant Odontoceti. Contributions in Science, Natural History Museum of Los Angeles County., 405:1-64.
Heyning, J.E. 1997. Sperm whale phylogeny revisited: analysis of the morphological evidence. Marine Mammal Science, 13(4):596-613.
Howell, A.B. 1927. Contribution to the anatomy of the Chinese finless porpoise Neomeris phocaenoides. Proceedings of The United States National Museum, 70(13):1-43.
Ichishima, H., and Kimura, M. 2005. Haborophocoena toyoshimai, a new Early Pliocene porpoise (Cetacea; Phocoenidae) from Hokkaido, Japan. Journal of Vertebrate Paleontology, 25(3):655-664.
Kasuya, T. 1973. Systematic consideration of recent toothed whales based on the morphology of tympano-periotic bone. Scientific Reports of the Whales Research Institute, Tokyo, 25:1-103.
Kellogg, R. 1936. A review of the Archaeoceti. Carnegie Institution of Washington publication, 482:1-366.
Lambert, O. 2005. Phylogenetic affinities of the long-snouted dolphin Eurhinodelphis (Cetacea, Odontoceti) from the Miocene of Antwerp, Belgium. Palaeontology, 48(3):653-679.
Lambert, O. 2008. A new porpoise (Cetacea, Odontoceti, Phocoenidae) from the Pliocene of the North Sea. Journal of Vertebrate Paleontology, 28(3):863-872.
Luo, Z., and Marsh, K. 1996. Petrosal (periotic) and inner ear of a Pliocene kogiine whale (Kogiinae, Odontoceti): implications on relationships and hearing evolution of toothed whales. Journal of Vertebrate Paleontology, 16(2):328-348.
Marsh, H., Lloze, R., Heinsohn, G.E., and Kasuya, T. 1989. Irrawaddy dolphin - Orcaella brevirostris (Gray, 1866), p. 101-118. In Ridgway, S.H., and Harrison, S. (eds.), Handbook of marine mammals. Volume 4: river dolphins and the larger toothed whales.
Mead, J.G., and Fordyce, R.E. 2009. The therian skull: a lexicon with emphasis on the odontocetes. Smithsonian Contributions to Zoology, 627:1-248.
Messenger, S.L., and McGuire, J.A. 1998. Morphology, molecules, and the phylogenetics of cetaceans. Systematic Biology, 47(1):90-124.
Miller, G.S. 1923. The telescoping of the cetacean skull. Smithsonian Miscellaneous Collections, 76(5):1-70.
Moore, J.C. 1968. Relationships among the living genera of beaked whales with classifications, diagnoses and keys. Fieldiana: Zoology, 53(4):509-598.
Murakami, M., Shimada, C., Hikida, Y., and Hirano, H. 2012a. A new basal porpoise, Pterophocaena nishinoi (Cetacea, Odontoceti, Delphinoidea), from the upper Miocene of Japan and its phylogenetic relationships. Journal of Vertebrate Paleontology, 32(5):1157-1171.
Murakami, M., Shimada, C., Hikida, Y., and Hirano, H. 2012b. Two new extinct basal phocoenids (Cetacea, Odontoceti, Delphinoidea), from the upper Miocene Koetoi Formation of Japan and their phylogenetic significance. Journal of Vertebrate Paleontology, 32(5):1172-1185.
Murakami, M., Shimada, C., Hikida, Y., Soeda, Y., and Hirano, H. 2014. Eodelphis kabatensis, a new name for the oldest true dolphin Stenella kabatensis Horikawa, 1977 (Cetacea, Odontoceti, Delphinidae), from the upper Miocene of Japan, and the phylogeny and paleobiogeography of Delphinoidea. Journal of Vertebrate Paleontology, 34(3):491-511.
Noble, B., and Fraser, F. 1971. Description of a skeleton and supplementary notes on the skull of a rare porpoise Phocoena sinus Norris & McFarland 1958. Journal of Natural History, 5(4):447-464.
Rommel, S. 1990. Osteology of the bottlenose dolphin, p. 29-49. In Leatherwood, S., and Reeves, R.R. (eds.), The bottlenose dolphin . Academic Press Inc, San Diego, New York.
Sanders, A.E., and Barnes, L.G. 2002. Paleontology of the late Oligocene Ashley and Chandler Bridge formations of South Carolina, 2: Micromysticetus rothauseni, a primitive cetotheriid mysticete (Mammalia: Cetacea). Smithsonian Contributions to Paleobiology (93):271-293.
Tanaka, Y., and Fordyce, R.E. 2014. Fossil dolphin Otekaikea marplesi (latest Oligocene, New Zealand) expands the morphological and taxonomic diversity of Oligocene cetaceans. PLoS ONE, 9(9):e107972.
Tanaka, Y., and Fordyce, R.E. 2015. A new Oligo-Miocene dolphin from New Zealand: Otekaikea huata expands diversity of the early Platanistoidea. Palaeontologia Electronica, 18(2.23A):1-71.
True, F.W. 1904. The whalebone whales of the western North Atlantic compared with those occurring in European waters with some observations on the species of the North Pacific. Smithson Contribution to Knowledge, 33:1-332.
Whitmore, F.C., and Sanders, A.E. 1977. Review of the Oligocene Cetacea. Systematic Zoology, 25(4):304-320.
APPENDIX 4.
List of modifications to the original codings and character descriptioons of Tanaka and Fordyce (2015). The changed codings were originally coded by Murakami (2012b). The initial number refers to the character number in Appendix 3. Character numbers are identified in parentheses.
(26) Maxilla on dorsal surface of skull: does not contact supraoccipital posteriorly, maxilla separated by frontal and/or parietal (0); contact present (1).
Haborophocoena toyoshimai ? to 0
(46) Temporal fossa shape in lateral view: height lower than anteroposterior length (0); higher (1); lower and its posterior end is rounded (2).
Haborophocoena minutus ? to 0
Haborophocoena toyoshiimai 0 to 1
(56) Postglenoid process in lateral view: tapering ventrally (0); squared off ventrally (1); same as state 1 except very wide anteroposterior diameter of process (2).
Haborophocoena toyoshimai 0 to 1
(84) Posterior process of periotic in lateral view: ventrally bent (0); in same plane as body of periotic (1).
Haborodelphis toyoshimai 0 to 1
Miophocaena nishinoi ? to 0
Semirostrum ceruttii 1 to 0
APPENDIX 5.
Treefile of the analysis of Tanaka and Ichishima (2016).
Available in zipped format with Appendix 1 and Appendix 2.
 Yoshihiro Tanaka. Numata Fossil Museum, 2-7-49, Minami 1, Numata Town, Hokkaido, 078-2225 Japan, yoshihiro.tanaka@otago.ac.nz and Hokkaido University Museum, Kita 10, Nishi 8, Kita-ku, Sapporo, Hokkaido 060-0810 Japan
Yoshihiro Tanaka. Numata Fossil Museum, 2-7-49, Minami 1, Numata Town, Hokkaido, 078-2225 Japan, yoshihiro.tanaka@otago.ac.nz and Hokkaido University Museum, Kita 10, Nishi 8, Kita-ku, Sapporo, Hokkaido 060-0810 Japan
Yoshi Tanaka received his Ph.D. from the University of Otago, New Zealand. He is a curator at Numata Fossil Museum, Hokkaido, Japan and also a researcher at Hokkaido University Museum. He works on the morphology and phylogeny of marine mammals (dolphins, porpoises, whales and walruses) from Japan and New Zealand. This paper is a part of his research project at Numata Town.

 Hiroto Ichishima. Fukui Prefectural Dinosaur Museum, Terao 51-11, Muroko, Katsuyama, Fukui 911-8601, Japan, hiroto.ichishima@dinosaur.pref.fukui.jp
Hiroto Ichishima. Fukui Prefectural Dinosaur Museum, Terao 51-11, Muroko, Katsuyama, Fukui 911-8601, Japan, hiroto.ichishima@dinosaur.pref.fukui.jp
Hiroto Ichishima received his Ph.D. from the University of Otago, New Zealand. He is currently a senior curator at Fukui Dinosaur Museum in Japan. He has worked on fossil cetaceans from Hokkaido, the northernmost island of Japan. His research interest includes vertebrate paleontology, whale anatomy, and evolution.
FIGURE 1. 1, Locality maps, 2, stratigraphic sections of the locality based on previous studies which are mentioned in the text. This figure is modified from Tanaka (2016).
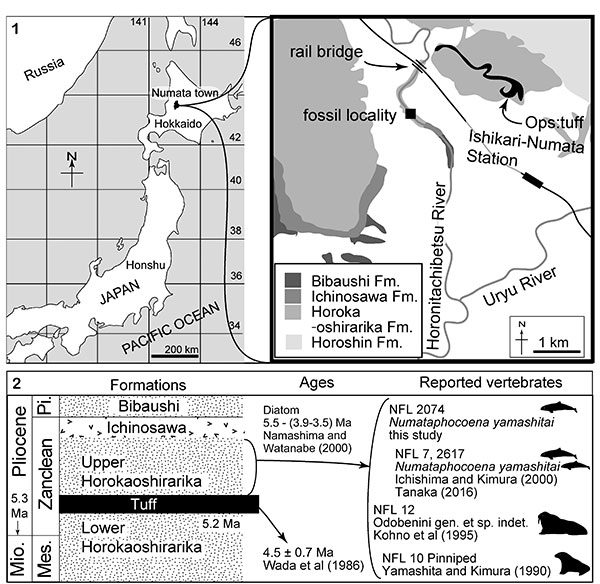
FIGURE 2. The skull, NFL 2074, referred specimen of Numataphocoena yamashitai in dorsal view. 1, photo, 2, line art.
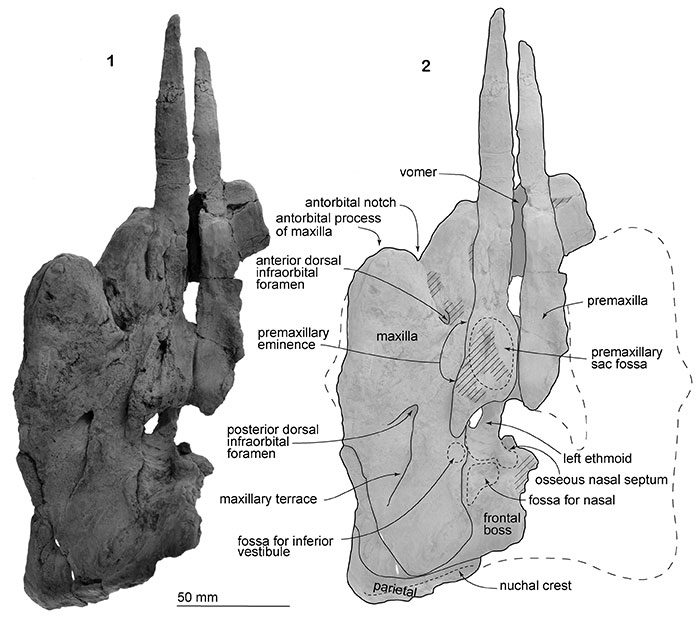
FIGURE 3. The skull, NFL 2074, referred specimen of Numataphocoena yamashitai in ventral view. 1, photo, 2, line art.
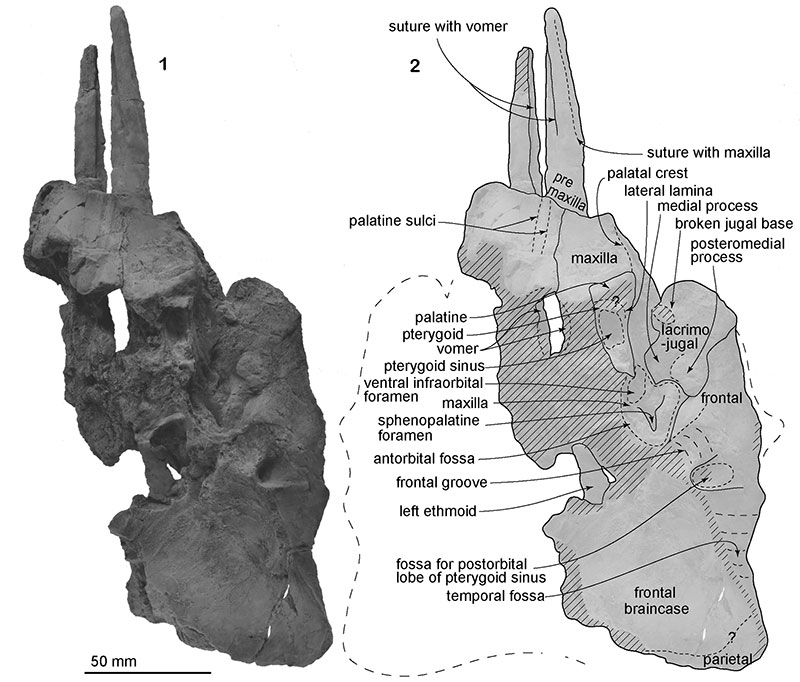
FIGURE 4. The skull, NFL 2074, referred specimen of Numataphocoena yamashitai in lateral view. 1, photo, 2, line art.
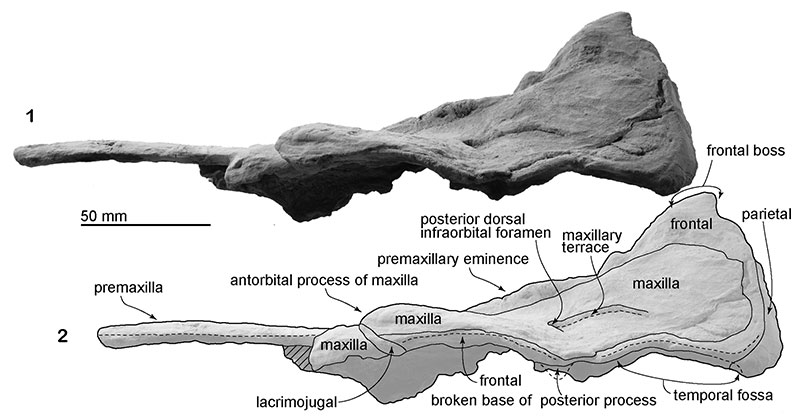
FIGURE 5. The skull, NFL 2074, referred specimen of Numataphocoena yamashitai in anterior view. 1, photo, 2, line art.
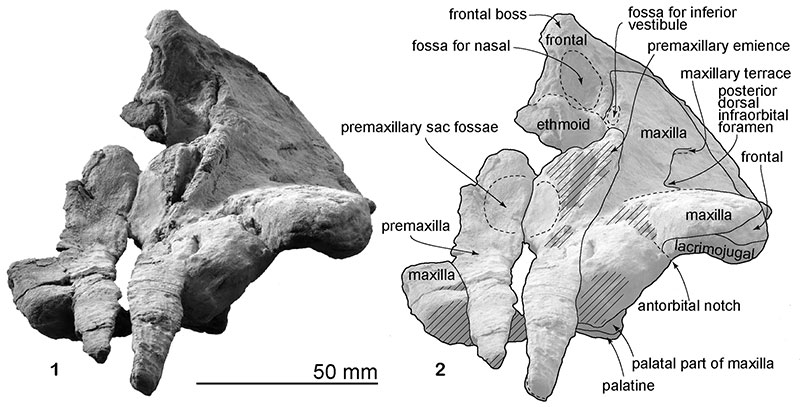
FIGURE 6. Life restoration of Numataphocoena yamashitai. Art work by Tatsuya Shinmura (Ashoro Museum of Paleontology).

FIGURE 7. A cladogram showing relationships of the Phocoenidae. Numbers represent branch lengths.
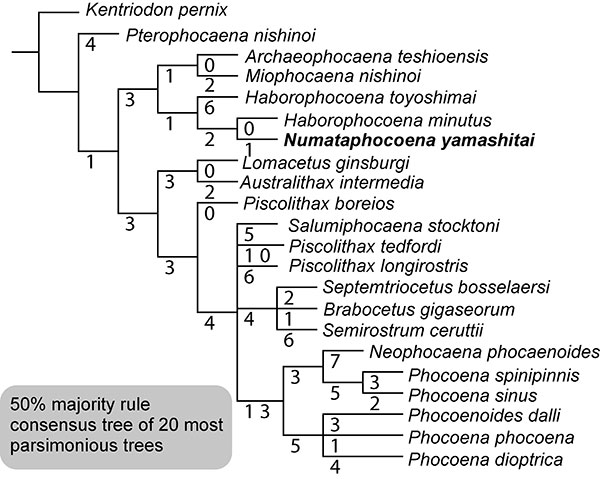
FIGURE 8. The maxillary terrace from the posterior dorsal infraorbital foramen on the maxilla. 1 and 2, NFL 7, the type of Numataphocoena yamashitai. 3 and 4, NFL 2074, the referred specimen.
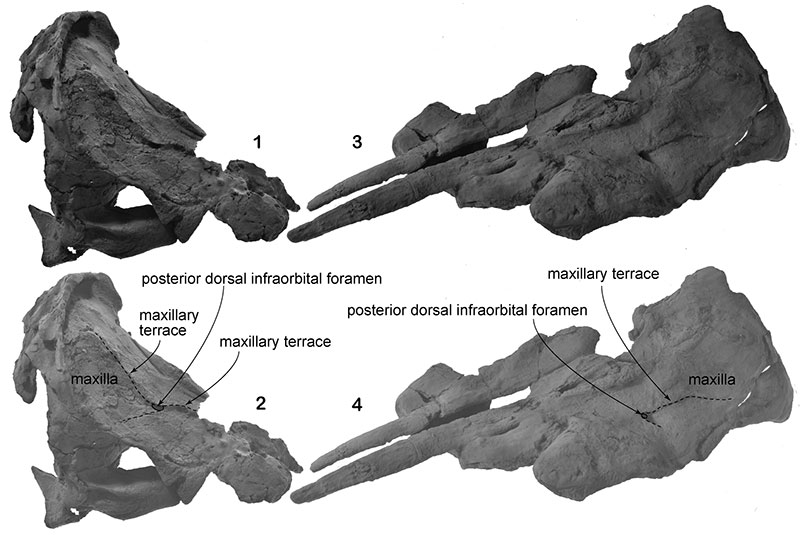
FIGURE 9. Geochronological distributions and postulated phylogeny of the Phocoenidae.
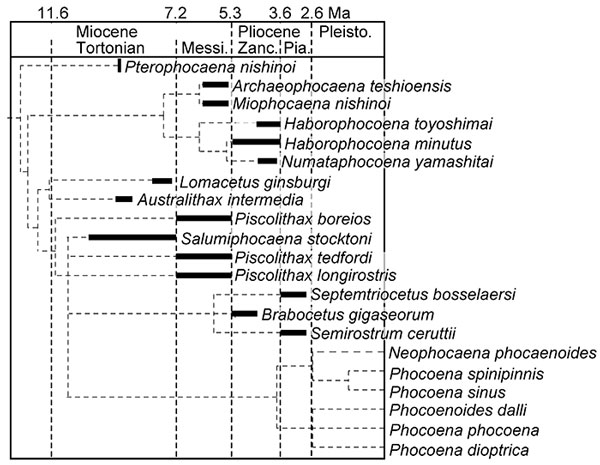
TABLE 1. Measurements in mm of NFL 2074, referred specimen of Numataphocoena yamashitai: skull. Dimensions follow Fordyce et al. (2002). Measurements are rounded to the nearest 0.5 mm. For skull and mandible, distances are either horizontal or vertical. * shows a measurement, which is only of the left side (the measurement points to the median line).
| Skull | NFL 2074 | NFL 7 |
| total length, from the most anterior point to posterior of occipital condyles | 260.0+ | 199.0+ |
| length of rostrum | 111.5+ | - |
| width of rostrum at the base | 43.5* | - |
| cranial length | 142.5+ | 199.0+ |
| width of premaxillae at a line across posterior limits of antorbital notches | 44.0* | - |
| maximum width of premaxillae about the level with mid-orbit | 25.0* | 56.0+ |
| postorbital width, across apices of postorbital processes | 85.5* | - |
| maximum width across narial aperture | 19.5* | - |
A new skull of the fossil porpoise Numataphocoena yamashitai (Cetacea: Phocoenidae) from the upper part of the Horokaoshirarika Formation (lower Pliocene), Numata Town, Hokkaido, Japan, and its phylogenetic position
Plain Language Abstract
An early Pliocene (about 4.5 to 3.5 million years ago) porpoise, Numataphocoena yamashitai from Hokkaido, northern Japan is known from only the holotype, a fairly wellpreserved skeleton with an incomplete skull and a referred earbone. A new skull referred to Numataphocoena yamashitai found from almost the same locality as the holotype is interesting because it provides information about the skull shape and adds a diagnostic character of the species. Numataphocoena yamashitai differs from other porpoises in having the characteristic feature in the maxilla associated with the posterior dorsal infraorbital foramen. A cladistic analysis places Numataphocoena yamashitai with fossil porpoises, such as Haborophocoena toyoshimai and Haborophocoena minutus from more or less the same age and locality.
Resumen en Español
Un cráneo nuevo de la marsopa fósil Numataphocoena yamashitai (Cetacea: Phocoenidae) de la parte superior de la Formación Horokaoshirarika (Plioceno inferior), ciudad de Numata, Hokkaido, Japón, y su posición filogenética
Una marsopa de edad pliocena temprana, Numataphocoena yamashitai de Hokkaido, Japón, se conoce a partir del holotipo, un esqueleto relativamente bien preservado con un cráneo incomplete y un hueso del oído referido. Un nuevo cráneo referido a Numataphocoena yamashitai hallado en casi la misma localidad que el holotipo es interesante porque expande el conocimiento acerca de la morfología craneana y mejora la diagnosis del taxon. Numataphocoena yamashitai difiere de otros focénidos por tener el rasgo característico en el maxilar asociado con el foramen infraorbital dorsal posterior, una parte anterior del meato acústico interno más angosta y aguda, y un proceso anterior del periótico robusto. Un nuevo análisis cladístico posiciona a Numataphocoena yamashitai adyacente a Haborophocoena toyoshimai y Haborophocoena minutus, entre un clado de focénidos de ramificación temprana, todos los cuales son cercanos cronológicamente y geográficamente. El nuevo cráneo es probablemente de un individuo más joven porque es de aproximadamente un 80% del tamaño del cráneo del holotipo y muestra suturas cerradas pero no fusionadas. Nuestra descripción de este espécimen ayuda a entender la variación intraespecífica de la especie extinta Numataphocoena yamashitai.
Palabras clave: cráneo; Phocoenidae; filogenia; características del maxilar; ontogenia; variación intraespecífica
Traducción: Diana Elizabeth Fernández
Résumé en Français
text
Translator: Kenny J. Travouillon or Antoine Souron
Deutsche Zusammenfassung
Ein neuer Schädel des fossilen Schweinswals Numataphocoena yamashitai (Cetacea: Phocoenidae) aus dem oberen Teil der Horokaoshirarika Formation (unteres Pliozän), Numata, Hokkaido, Japan und seine phylogenetische Position
Der frühpliozäne Schweinswal, Numataphocoena yamashitai from Hokkaido, Japan, ist durch einen Holoty bekannt, der aus einem gut erhaltenen Skelett mit unvollständigem Schädel und dem dazugehörigen Gehörknochen besteht. Ein neuer Schädel von Numataphocoena yamashitai aus nahezu der gleichen Fundstelle wie der Holotyp ist interessant, da dadurch die Kenntnis über die Schädelmorphologie erweitert und die Taxon-Diagnose verbessert wird. Numataphocoena yamashitai unterscheidet sich von anderen Phocoeniden durch die Assoziation des charakteristischen Merkmals im Oberkiefer mit dem posterioren dorsalen Infraorbitalforamen, den schmaleren und scharfkantigeren inneren Gehörgang und einen robusten anterioren Fortsatz im Perioticum. Eine neue cladistische Analyse stellt Numataphocoena yamashitai neben Haborophocoena toyoshimai und Haborophocoena minutus imnitten einer Klade von frühen abzweigenden Phocoendien von denen alle chronologisch und geographisch nahe beieinander stehen. Der neue Schädel gehört möglicherweise einem Jungtier, da seine Größe nur ungefähr 80% von der des Holotyps beträgt und er zeigt geschlossene aber nicht verschmolzene Suturen. Unsere Beschreibung trägt zum Verständnis der intraspezifischen Variation der ausgestorbenen Art Numataphocoena yamashitai bei.
Schlüsselwörter: Schädel; Phocoenidae; Phylogenie; Maxillarstufen; Ontogenie; intraspezifische Variation
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024