

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
FIGURE 1. Schematic geological map with location of the investigated section (1). The study section is located at Portella Colla (2) in the Madonie Mountains, Sicily (3).
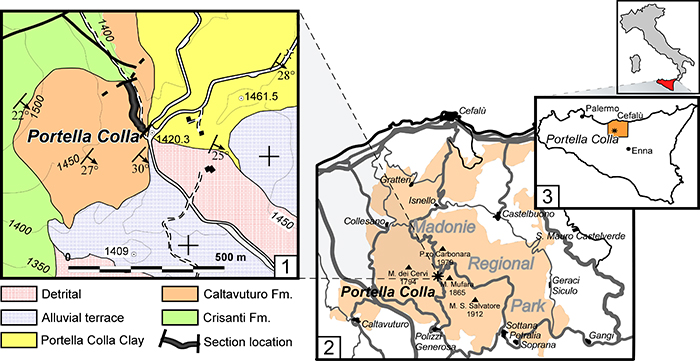
FIGURE 2. Biostratigraphic interpretation of the investigated section. According to Benedetti (2010), the distribution of the main larger foraminifers in the displaced layers allows the recognition of four biozones in the Paleogene succession. The presence of SBZ 18 (late Bartonian) at the base of Paleogene succession is uncertain. The E/O boundary is here redefined by the study of poorly-preserved nannofossils occurring only in some levels, and following the biozonal schemes of Martini (1971), and Catanzariti et al. (1997).
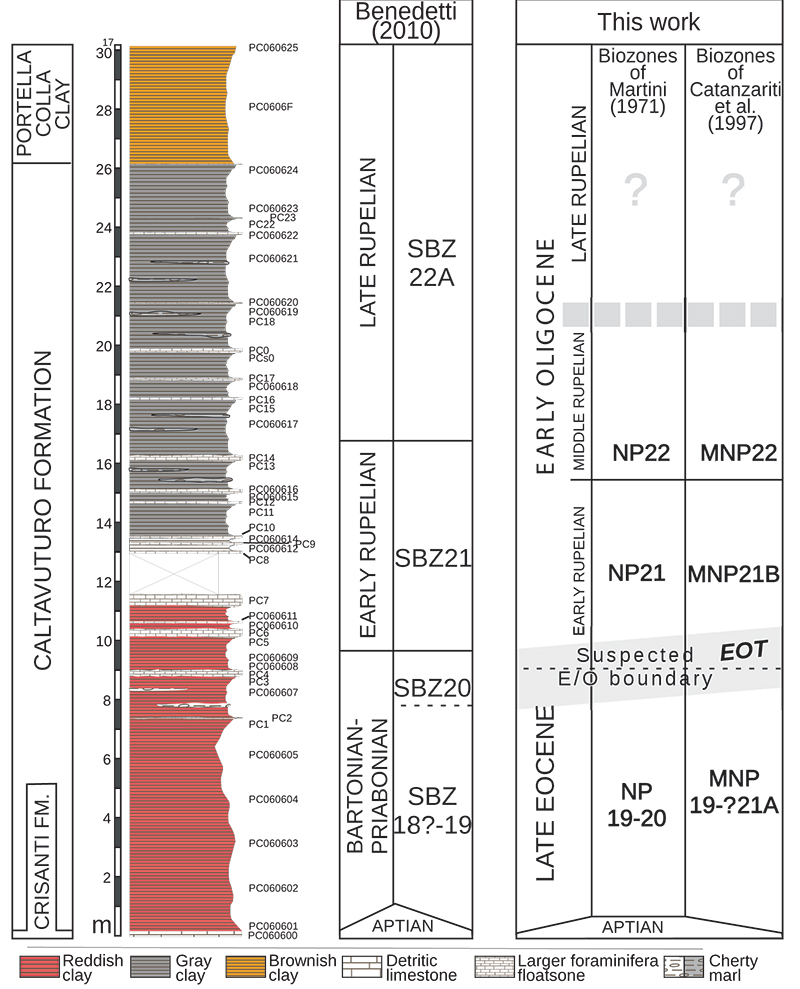
FIGURE 3. Representative forms of foraminifers belonging to the six different functional morphogroups (redrawn after Jones and Charnock, 1985, figure 2).
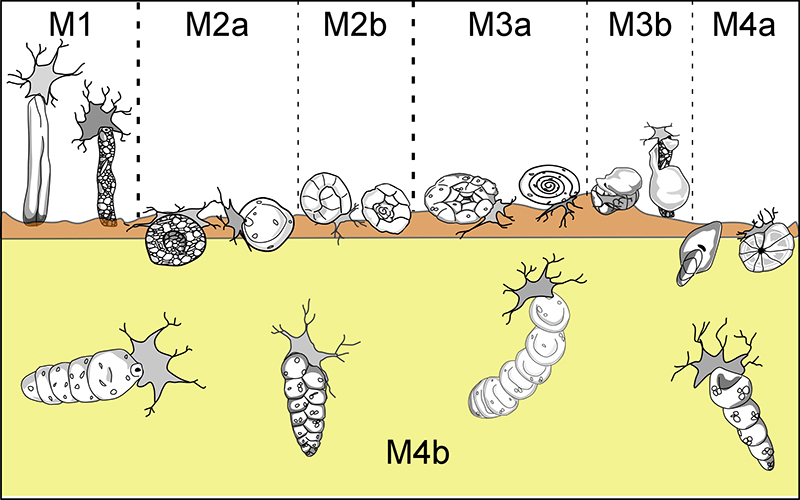
FIGURE 4. Summarizing scheme of the DWAF functional morphogroups and morphotypes described in this work (redrawn and modified after van den Akker et al., 2000, figure 6).
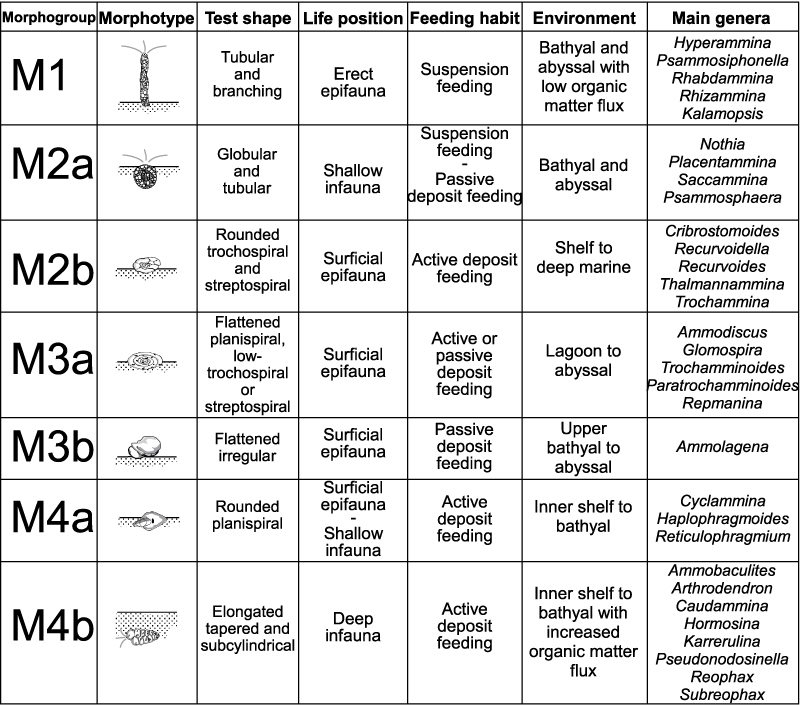
FIGURE 5. Results of the micropaleontological analysis along the investigated section cropping out at Portell Colla (Sicily). Vertical distribution of individuals counted per sample, number of taxa, Fisher’s Alpha diversity, species richness, evenness index, Simpson’s index, and percentage of CaCO 3 within the samples. The three boxes represent the relative abundances of agglutinated and calcareous hyaline taxa, the DWAF morphogroups, and the recognized assemblages.
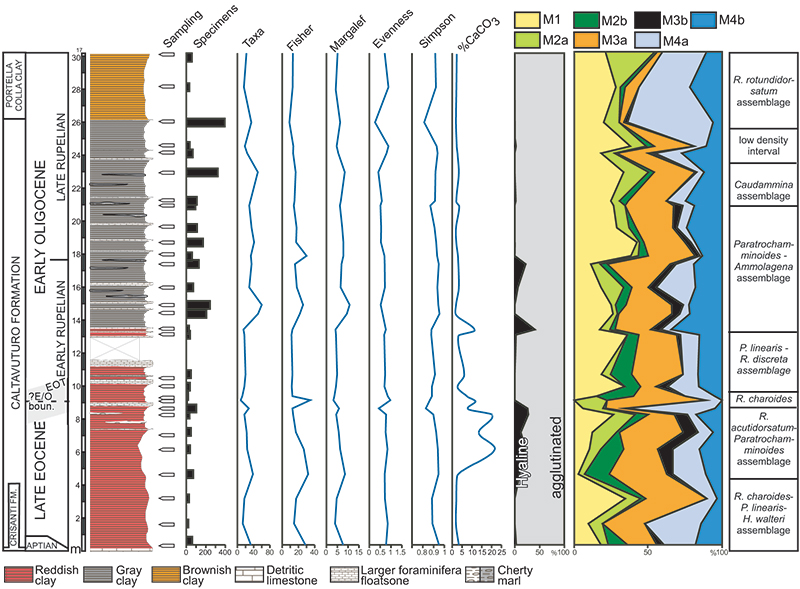
FIGURE 6. Stratigraphic distribution (in percentage) of selected DWAF taxa (solid line). The abundances have been exaggerated (dashed lines) in order to show also the minimal fluctuations.
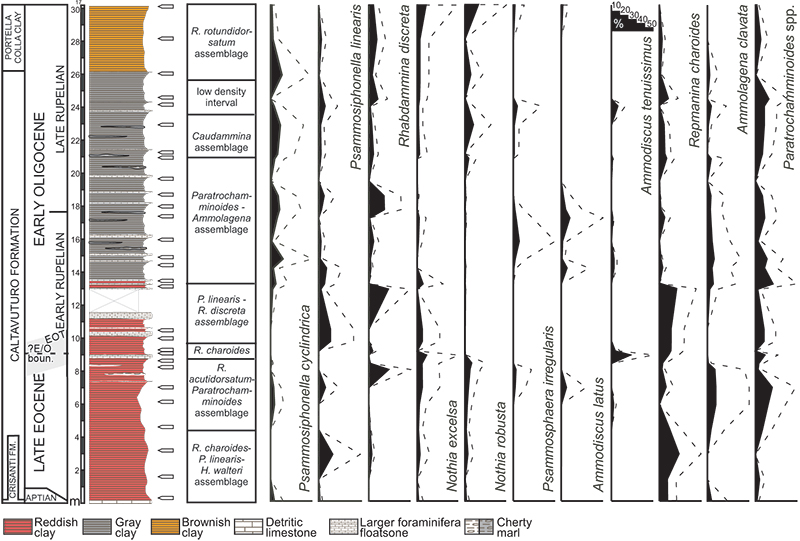
FIGURE 7. Stratigraphic distribution (in percentage) of selected DWAF taxa (solid line). The abundances have been exaggerated (dashed lines) in order to show also the minimal fluctuations.
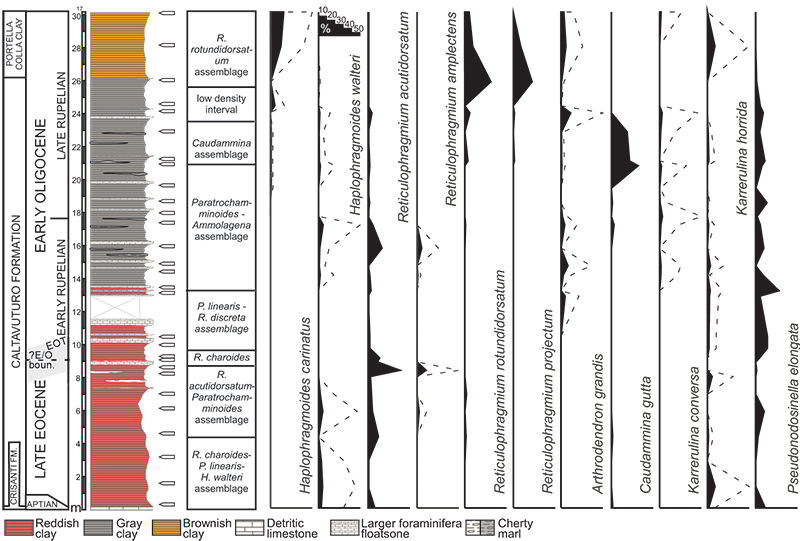
FIGURE 8. Environmental interpretation of the investigated succession. Block diagrams schematically show the presence/absence of main taxa and the faunal density.
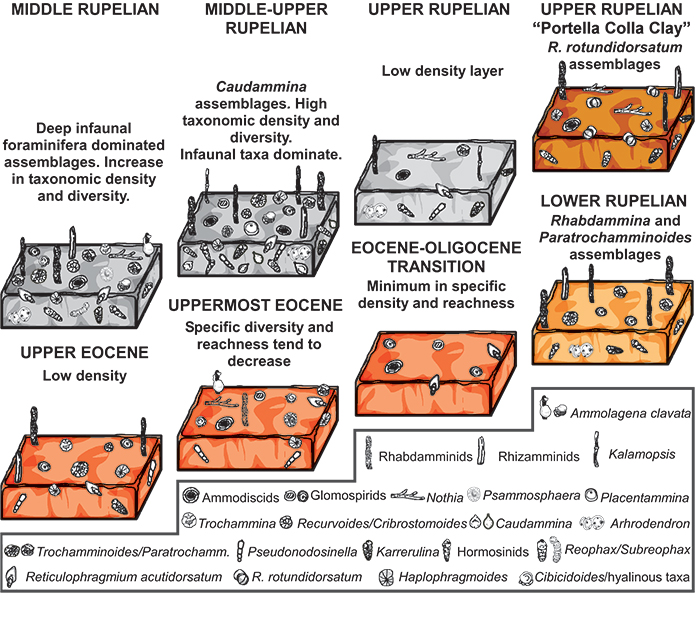
FIGURE 9. Schematic zonal schemes of late Eocene-lower Oligocene successions described from different regions (redrawn and modified after Kaminski and Gradstein, 2005). The most common DWAF taxa are added.
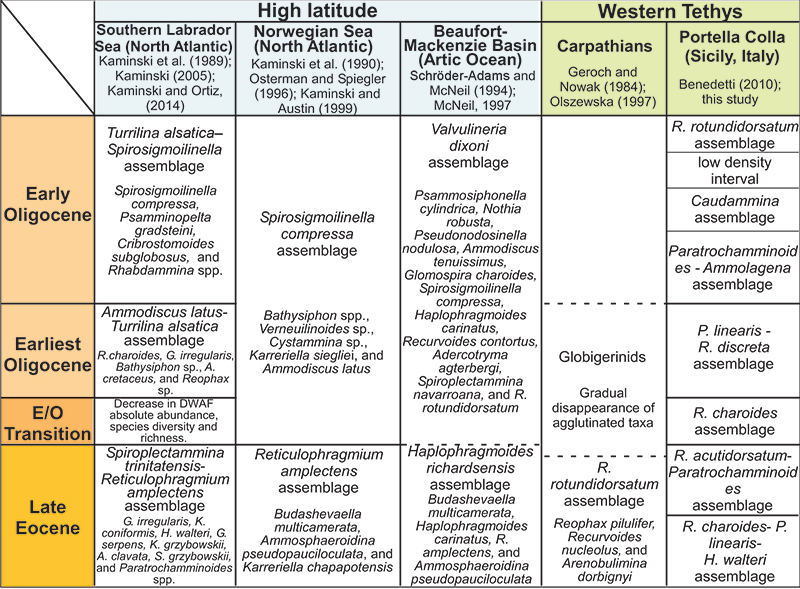
FIGURE 10. Depth distribution of some selected and frequent taxa according to data presented by van Morkhoven et al. (1986), Kaminski and Gradstein (2005) and Waśkowa (2014).
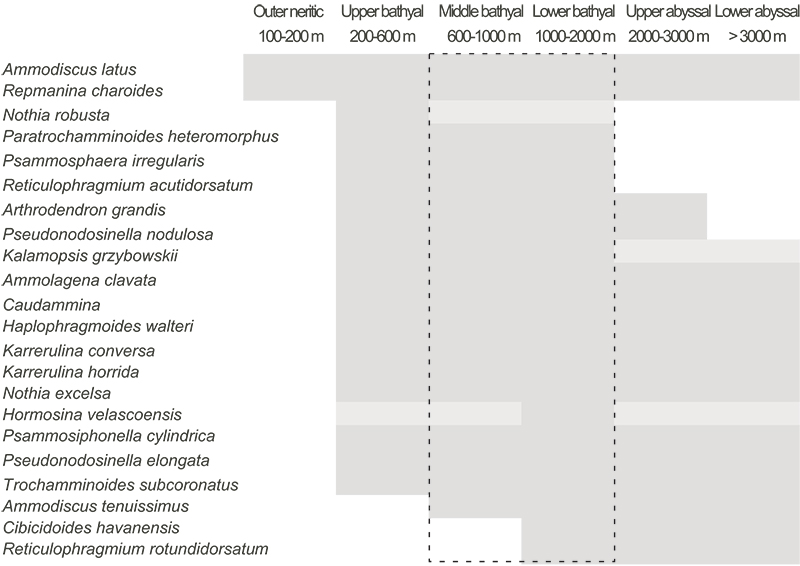
FIGURE 11. Location of the section FO; the reddish clay are interbedded by quartzose volcanic layers marked by the metallic labels.
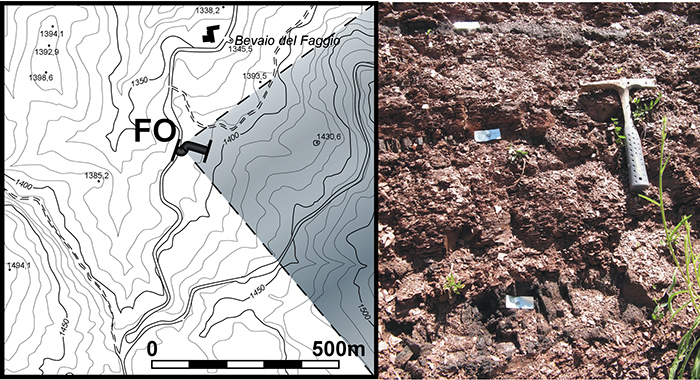
FIGURE 12. Scanning electron micrographs of Astrorhizida ( 1-13) and Saccamminidae (14-15) foraminifers from the Caltavuturo Formation cropping out at Portella Colla. 1-2, Rhabdammina discreta Brady, 1884, PCs0 (1) and PC060618 (2). 3,Rhabdammina eocenica Cushman and Hanna, 1927, PC060601. 4-5,Psammosiphonella cylindrica (Glaessner, 1937), PC11 (4) and PCs0 (5). 6-7,Psammosiphonella linearis (Brady, 1879), PC060618 (6) and PC11 (7). 8,Bathysiphon sp., MM12. 9,Nothia excelsa (Grzybowski, 1898), PC060603. 10,Nothia robusta (Grzybowski, 1898) with Ammolagena clavata (Jones and Parker, 1860), PC060621. 11, Nothia cf. latissima (Grzybowski, 1898), PC060624. 12-13, Rhizammina indivisa Brady, 1884, PC060621. 14-15,Psammosphaera irregularis (Grzybowski, 1896), PC13 (14) and PC22 (15). 16,Placentammna placenta (Grzybowski, 1898), PCs0. 17,Saccammina grzybowskii (Schubert, 1902), PC060621. Scale bar equals 0.5 mm.
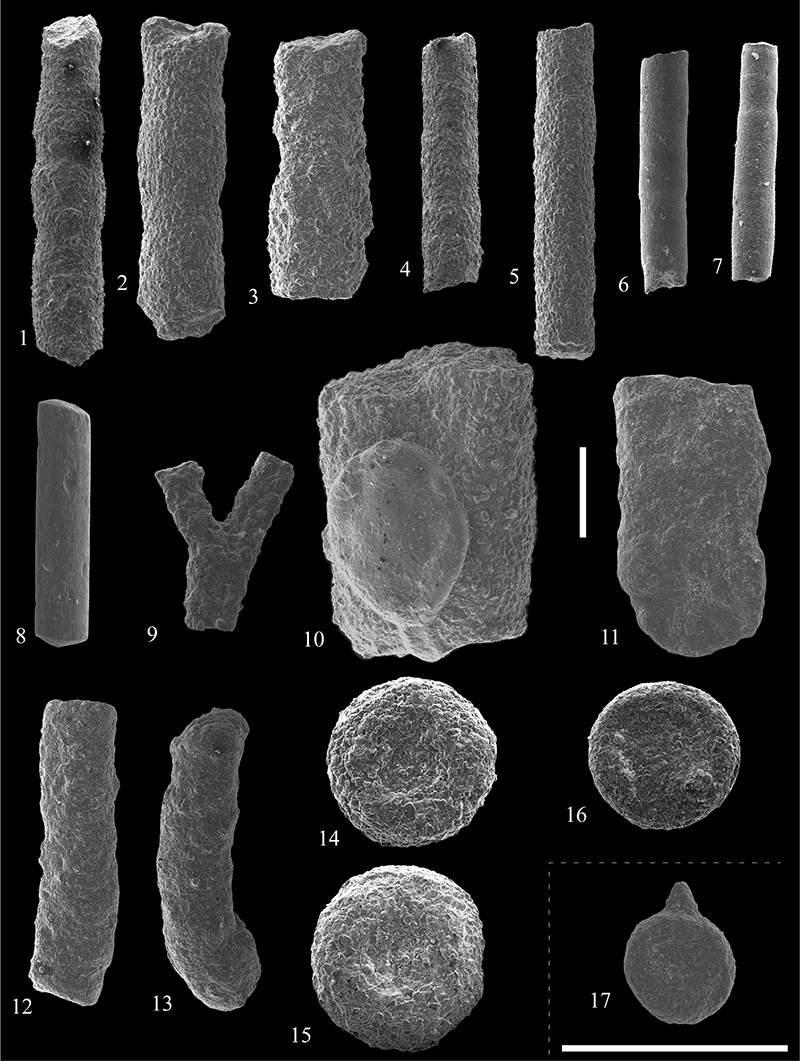
FIGURE 13. Scanning electron micrographs of Psammosphaeridae (1-4) and Ammodiscidae (5-27) from the Caltavuturo Formation cropping out at Portella Colla. 1,Psammosphaera cf. laevigata White, 1928, PCs0. 2,Psammosphaera sp. 1, PC13. 3, Psammosphaera sp. 2, PC060615. 4,Psammosphaera sp. 3, PC060625. 5,Ammodiscus cretaceus (Reuss, 1845), PC22. 6,Hyperammina sp., PC060625. 7-8,Ammodiscus incertus (d’Orbigny, 1839), PC060603 (7) and PC13 (8). 9-11,Ammodiscus tenuisimus (Gümbel, 1862), PC22 (9), PC060615 ( 10) and PC060615 (11). 12-13,Ammodiscus latus Grzybowski, 1898, PC11 (12) and PC11 (13). 14,Ammodiscus peruvianus Berry, 1928, PC060617. 15-16,Ammodiscus cf. latus Grzybowski, 1898, PC11. 17, Ammodiscus sp. 1, PC11. 18,Ammodiscus sp. 2, PC15. 19,Ammodiscus sp. 3, PC060603. 20,Annectina biedai Gradstein and Kaminski, 1997, PC060618. 21,Annectina cf. grzybowski (Jurkiewicz, 1960), PC060617. 22,Glomospira sp. 2, PC15. 23,Glomospira sp. 1, PC11. 24, Glomospira sp. 3, PC13. 25-26. Glomospira serpens (Grzybowski, 1898), PC060617 (25) and PC060618 (26). 27,Glomospira irregularis (Grzybowski, 1898), PC060618. Scale bar equals 0.5 mm.
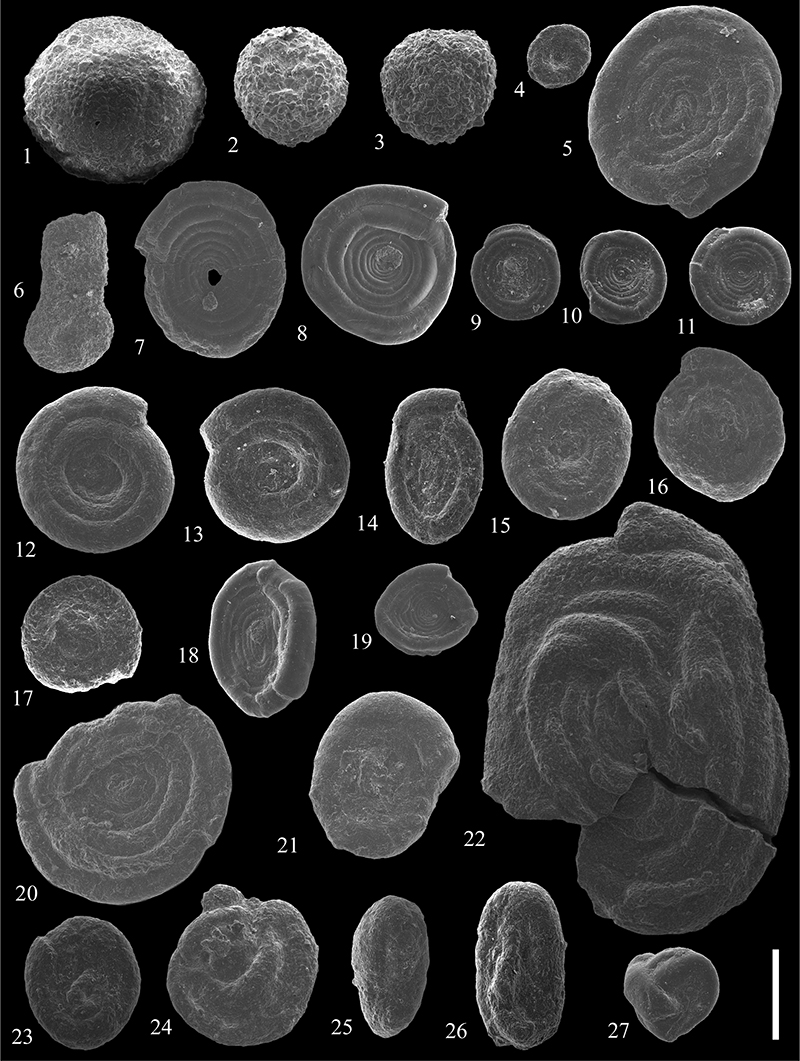
FIGURE 14. Scanning electron micrographs of Hormosinelloidea from the Caltavuturo Formation cropping out at Portella Colla. 1, Ammolagena clavata (Jones and Parker, 1860) on a specimen of Psammosiphonella cylindrica (Glaessner, 1937), PCs0. 2,Ammolagena clavata (Jones and Parker, 1860), PC060607. 3-7,Caudammina gutta Benedetti and Pignatti, 2009, PC18. 8,Subreophax cf. guttifer (Brady, 1881), PC15. 9,Subreophax cf. pseudoscalaris (Samuel, 1977), PCs0. 10,Subreophax scalaris (Grzybowski, 1896), PC11. 11-12,Subreophax splendidus (Grzybowski, 1898), PC060624 (11) and PC11 ( 12). 13,Arthrodendron subnodosiformis (Grzybowski, 1898), PC060615. 14,Arthrodendron grandis (Grzybowski, 1898), PC050515. 15-19, Kalamopsis grzybowskii (Dylążanka, 1923), PC18 (15), PC11 (16), PC18 (17), PC060621 (18) and PC060621 (19). Scale bar equals 0.5 mm.

FIGURE 15. Scanning electron micrographs of Usbekistaniinae from the Caltavuturo Formation cropping out at Portella Colla. 1,Glomospira extendens Emiliani, 1954, PC11. 2,Glomospira gordialis (Jones and Parker, 1860), PC060604. 3-16,Repmanina charoides (Jones and Parker, 1860), PC060604 (3), PC060601 (4), PC060604 ( 5), PC060603 (6), PC060611 (7), PC060601 (8), PC060603 (9), PC060621 (10), PC060621 (11), PC060617 (12), PCs0 (13), PCs0 (14), PC060604 (15) and PCs0 (16). Scale bar equals 0.5 mm.
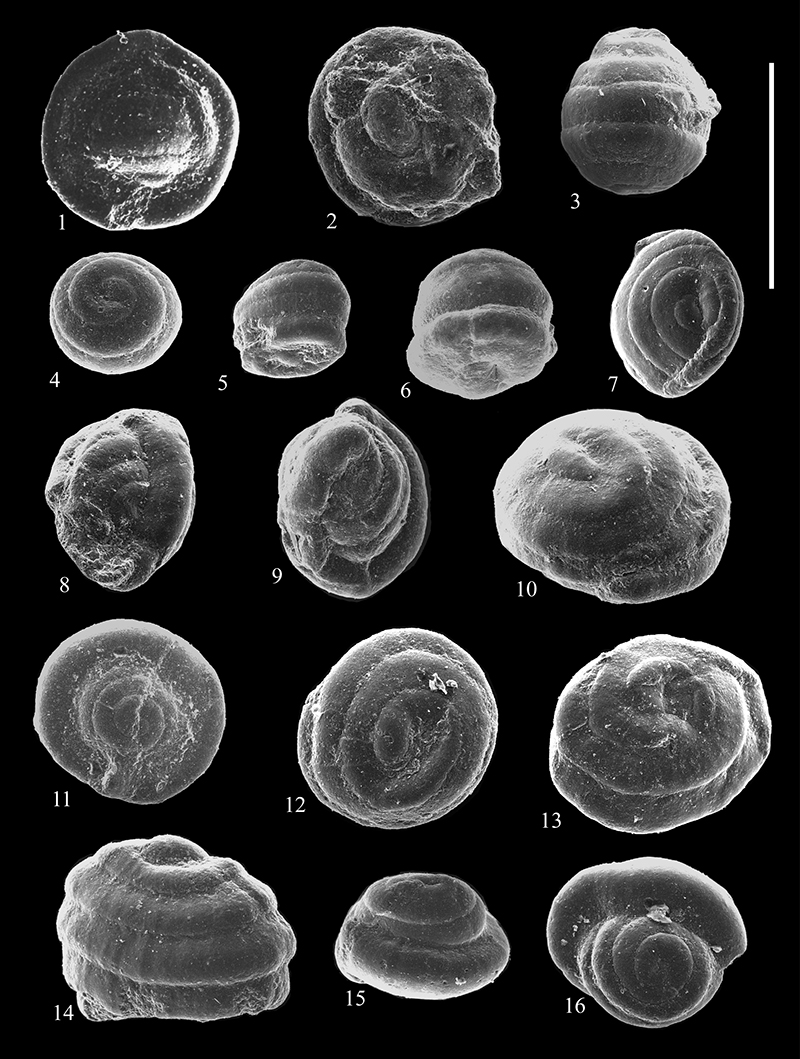
FIGURE 16. Scanning electron micrographs of Reophacidae (1-3), Hormosinidae (4-13), and Lituotubidae (14-20) from the Caltavuturo Formation cropping out at Portella Colla. 1,Reophax duplex Grzybowski, 1896, PC060624. 2,Reophax pilulifer Brady, 1884, PC13. 3,Reophax sp. 1, PC060615. 4-5,Hormosina velascoensis (Cushman, 1926), PC060617 (4) and PC11 (5). 6,Hormosina trinitatensis Cushman and Renz, 1946, PC11. 7,Hormosina sp. 1, PC060624. 8,Hormosina sp. 2, PC060615. 9-11,Pseudonodosinella elongata Grzybowski, 1898, PC11 (9), PC060601 (10) and PCs0 (11). 12-13,Pseudonodosinella nodulosa Brady, 1879, PC11 (12) and PC060617 ( 13). 14,Paratrochamminoides acervulatus (Grzybowski, 1896), PC13. 15,Paratrochamminoides deflexiformis (Noth, 1912), PC060615. 16,Lituotuba lituiformis (Brady, 1879), PC060615. 17,Paratrochamminoides sp. 1, PC060601. 18,Paratrochamminoides draco (Grzybowski, 1901), PC11. 19,Paratrochamminoides mitratus (Grzybowski, 1901), PCs0. 20,Conglophragmium deforme (Grzybowski, 1898), PC060623. Scale bar equals 0.5 mm.
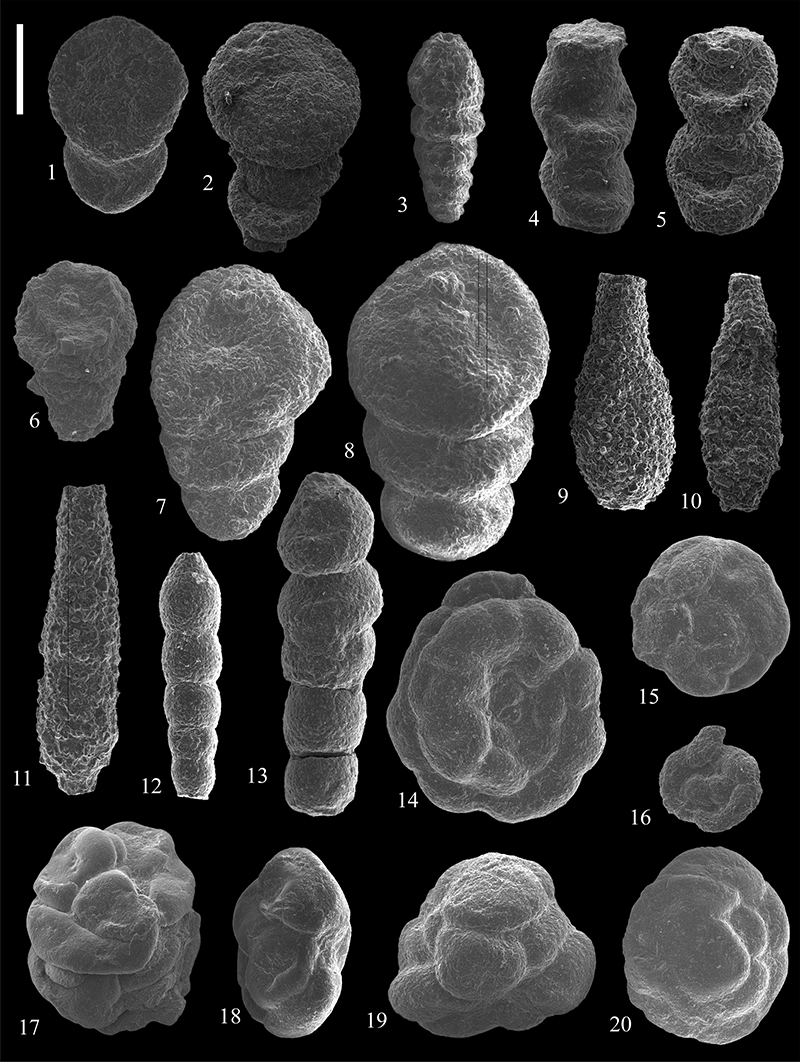
FIGURE 17. Scanning electron micrographs of Lituotubidae (1-3, 7, 9, 11-12) and Trochamminoidae (4-6, 8, 10) from the Caltavuturo Formation cropping out at Portella Colla. 1-2, Paratrochamminoides heteromorphus (Grzybowski, 1898), MM12 (1) and MM14 (2). 3,Conglophragmium irregulare (White, 1928), PCs0. 4,Trochamminoides coronatus (Brady, 1879), MM14. 5,Trochamminoides grzybowskii Kaminski and Geroch, 1992, MM1. 6,Trochamminoides subcoronatus (Grzybowski, 1896), MM1. 7,Paratrochamminoides cf. olszewskii (Grzybowski, 1898), PC060615. 8,Trochamminoides dubius (Grzybowski, 1901), PC060611. 9,Paratrochamminoides olszewskii (Grzybowski, 1898), PC11. 10,Trochamminoides dubius (Grzybowski, 1901), PC060611. 11,Paratrochamminoides aff. olszewskii (Grzybowski, 1898), PC060615. 12,Paratrochamminoides aff. gorayskii (Grzybowski, 1898), PC060615. Scale bars equal 0.5 mm.
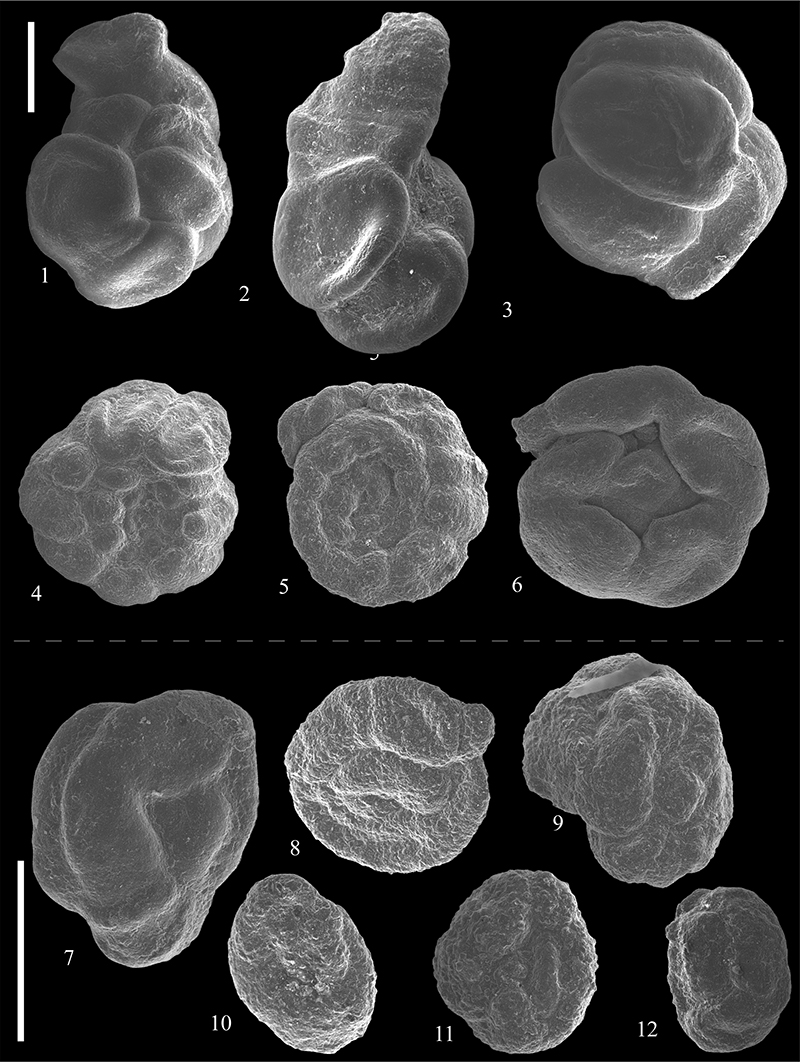
FIGURE 18. Scanning electron micrographs of Trochamminoidae ( 1-10) and Haplophragmoididae (11-27) from the Caltavuturo Formation cropping out at Portella Colla. 1,Trochamminoides septatus (Grzybowski, 1898), PC060604. 2,Trochamminoides proteus (Karrer, 1866), PC060621. 3, Trochamminoides cf. proteus (Karrer, 1866), PC060624. 4,Trochamminoides velascoensis Cushman, 1926, PC11. 5,Trochamminoides intermedius (Grzybowski, 1896), PC060621. 6,Trochamminoides variolarius (Grzybowski, 1898), MM14. 7,Trochamminoides sp. 1, PC060603. 8,Trochamminoides sp. 2, PC060604. 9, Trochamminoides sp. 3, PC060618. 10,Trochamminoides sp. 4, PC18. 11-13,Haplophragmoides carinatus Cushman and Renz, 1941, PC060621 (11), PC060621 (12) and PC060624 (1314-16,Haplophragmoides walteri (Grzybowski, 1898), PC18 (14), PC11 (15), PC5 (16). 17, Haplophragmoides eggeri Cushman, 1926, PC060611. 18, Haplophragmoides excavatus Cushman and Waters, 1927, PC060601. 19,Haplophragmoides cf. kirki Wickenden, 1932, PC060602. 20,Haplophragmoides porrectus Maslakova, 1955, PCs0. 21, Haplophragmoides cf. walteri (Grzybowski, 1898), PC060621. 22,Haplophragmoides horridus (Grzybowski, 1901), PC060601. 23, Haplophragmoides cf. horridus (Grzybowski, 1901), PC060604. 24,Haplophragmoides cf. porrectus Maslakova, 1955, PC060615. 25-26,Haplophragmoides sp. 1, PC060602 (25) and PC060602 (26). 27,Haplophragmoides sp. 2, PC060603. Scale bar equals 0.5 mm.
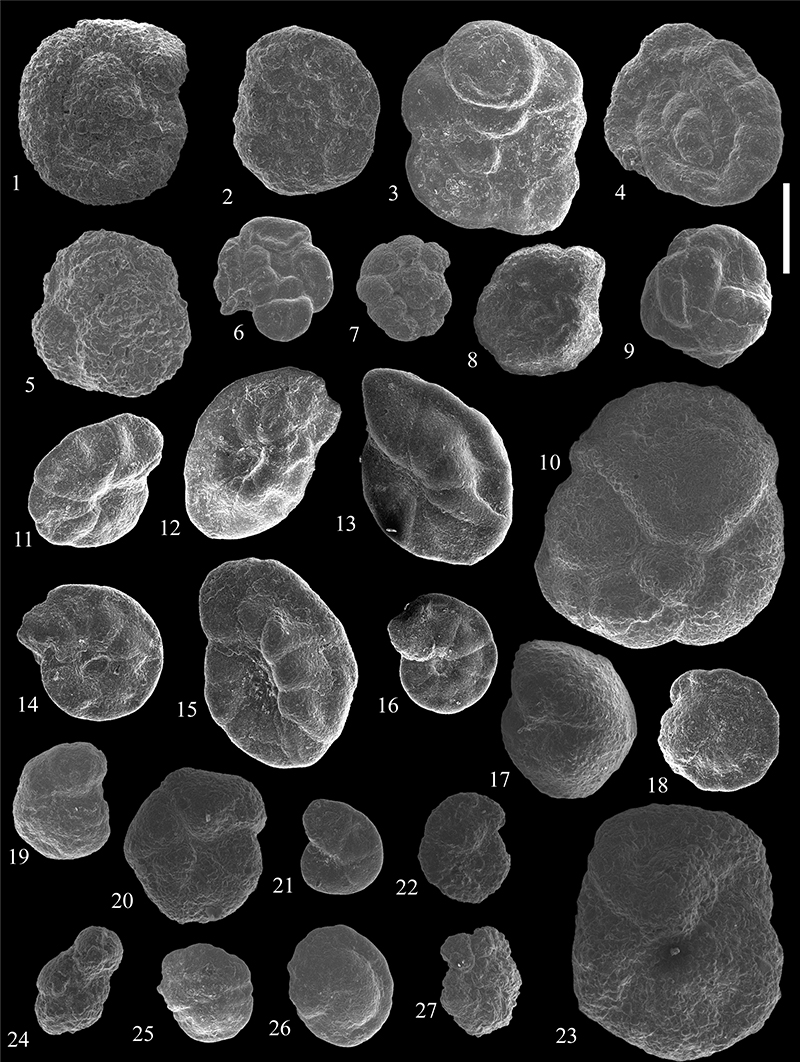
FIGURE 19. Scanning electron micrographs of Haplophragmoididae ( 1-6), Sphaeramminidae (7-10) Lituolidae (11-14), and Recurvoidea (15-25) from the Caltavuturo Formation cropping out at Portella Colla. 1-2,Haplophragmoides cf. latissimusuturalis Smith, 1971, PC3. 3,Haplophragmoides sp. 3, PC060615. 4Haplophragmoides sp. 4, PC060601. 5, Haplophragmoides sp. 5, PC060615. 6,Haplophragmoides sp. 6, PC060621. 7-8,Praesphaerammina subgaleata (Vašiček, 1947), PC11 (7) and PC1 (8). 9-10, Ammosphaeroidina pseudopauciloculata (Mjatliuk, 1966), PC060604 (9) and PC060604 (10). 11,Ammobaculites agglutinans (d’Orbigny, 1846), PC060615. 12,Ammobaculites sp. 1, PC060607. 13,Ammobaculites sp. 3, PC15. 14, Ammobaculites sp. 2, PC060625. 15-16,Budashevaella multicamerata (Voloshinova and Budasheva, 1961), PC060604. 17, Cribrostomoides subglobosus (Cushman, 1910), PC060604. 18, Recurvoidella lamella (Grzybowski, 1898), PC060601. 19, Recurvoides anormis Mjatliuk, 1970, PC5. 20,Recurvoides sp. 1, PC060601. 21,Recurvoides sp. 2, PC060623. 22,Recurvoides nucleolus (Grzybowski, 1898), MM3. 23, Recurvoides walteri (Grzybowski, 1898), PC5. 24,Trochammina bifaciata Friedberg, 1901, PC060604. 25,Thalmannammina subturbinata (Grzybowski, 1898), MM14. Scale bar equals 0.5 mm.

FIGURE 20. Scanning electron micrographs of Trochamminoidea ( 1-11, 13), Ataxophragmiidae (12, 17, 20-21), and Textulariina ( 14-16, 18-19, 22-23) from the Caltavuturo Formation cropping out at Portella Colla. 1,Trochammina sp. 1, PC060601. 2,Trochammina sp. 2, PC060601. 3,Trochammina sp. 3, PC060601. 4,Trochammina sp. 4, PC060604. 5, Trochammina sp. 5, PC060605. 6-10,Karrerulina conversa (Grzybowski, 1901), PC060615 (6), PC018 (7), PC11 (8), PC060615 (9) and PCs0 ( 10). 11, Karrerulina horrida (Mjatliuk, 1970), PC060617. 12,Remesella varians (Glaessner, 1937), PC060601. 13,Gaudryina sp., PC1. 14,Eggerella compressa (Andreae, 1884), PC13. 15,Eggerella sp. 1, PC3. 16,Eggerella sp. 2, PC3. 17,Arenobulimina sp., PC060602. 18,Siphotextularia sp. 1, PC060621. 19,Siphotextularia sp. 2, PC060621. 20,Tetraxiella subtilissima PCs0. 21, Gravellina sp., PC060618. 22,Valvulina flexilis Cushman and Renz, 1941, PC060604. 23,Valvulina sp., PC060621. Scale bars equal 0.5 mm.
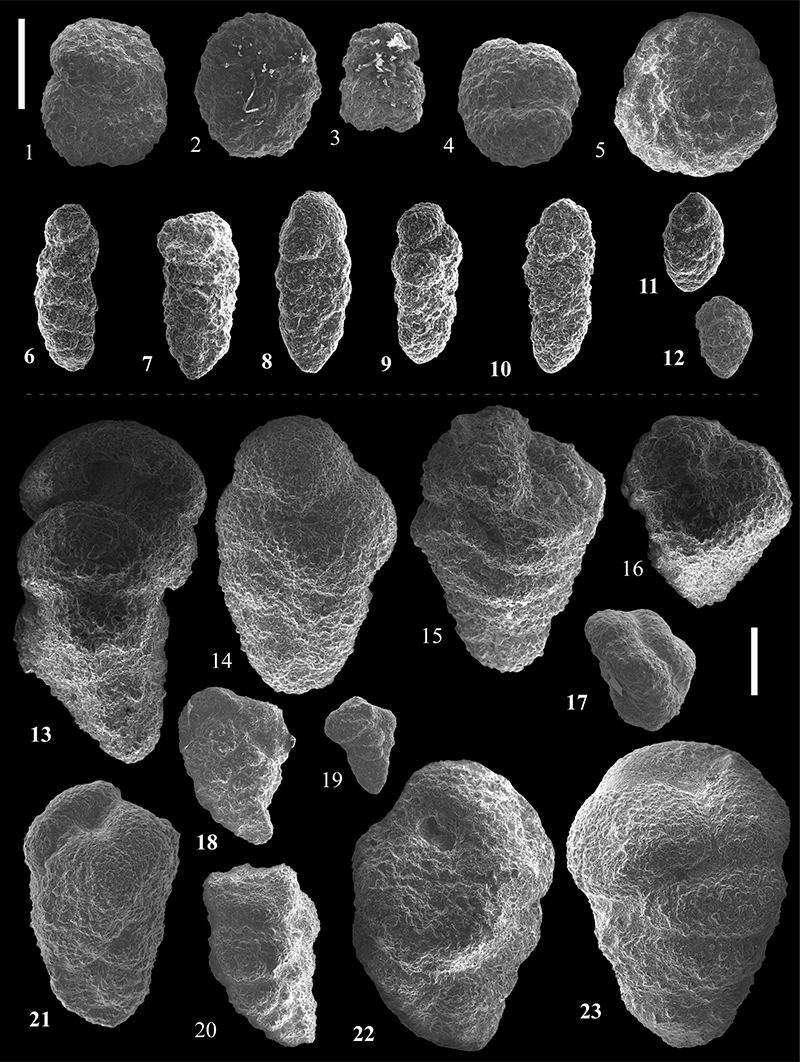
FIGURE 21. Scanning electron micrographs of Cyclamminidae from the Caltavuturo Formation cropping out at Portella Colla. 1-3, Reticulophragmium acutidorsatum (Hantken, 1868), PC3. 4-7, Reticulophragmium amplectens (Grzybowski, 1898), PC3. 8-10, Reticulophragmium rotundidorsatum (Hantken, 1875), PC060624. 11-12,Cyclammina cancellata Brady, 1879, PC22. 13-14, Reticulophragmium projectum Schröder-Adams and McNeil, 1994, PC060624. 15, Cyclammina placenta (Reuss, 1851), PC11. Scale bar equals 0.5 mm.
 Andrea Benedetti. Dipartimento di Scienze della Terra, “Sapienza” Università di Roma, P.le A. Moro 5, 00185, Italy; GIRMM -- Informal Group of Micropaleontological and Malacological Researches. andrea.benedetti@uniroma1.it
Andrea Benedetti. Dipartimento di Scienze della Terra, “Sapienza” Università di Roma, P.le A. Moro 5, 00185, Italy; GIRMM -- Informal Group of Micropaleontological and Malacological Researches. andrea.benedetti@uniroma1.it
Andrea Benedetti is a micropaleontologist and an independent contractor at the University of Rome "La Sapienza", where he got his undergraduate and postgraduate degrees. His main interests are focused on systematic, ecology, biostratigraphy and evolution of larger foraminifers in the present and past, and on the deep water foraminifers from Paleogene sedimentary successions of Sicily (Southern Italy). During the last three years he focused his scientific attention on the biometry and morphology of Larger Foraminifers with speciel regards on the orbitoidiform Nephrolepidina and the nummulitid Heterostegina.
Taxonomic list and results of the counts of the foraminiferal tests (available in PDF format).
Eocene/Oligocene deep-water agglutinated foraminifers (DWAF) assemblages from the Madonie Mountains (Sicily, Southern Italy)
Plain Language Abstract
In this study, 29 samples from a 30 m thick sedimentary succession of the Caltavuturo Formation cropping out at Portella Colla (Madonie Mts.) were investigated to characterize the benthic foraminiferal assemblages. The stratigraphic section spans from the late Eocene to the early Oligocene (Rupelian), and agglutinated foraminifers dominate along the whole section, whereas calcareous-walled species are rare and poorly preserved except for the resedimented larger foraminifers occurring in numerous turbiditic layers. Detailed systematic analysis of 138 agglutinated taxa has revealed a faunal turnover in proximity to the Eocene/Oligocene transition and the establishment of a low diversity assemblage dominated by opportunistic species in an oligotrophic environment. During the Oligocene the organic matter flux increases and the oxygen content in the seafloor sediments decreases. The occurrence of new and poorly known species is discussed.
Resumen en Español
Asociaciones de foraminíferos aglutinados de aguas profundas (DWAF) del Eoceno/Oligoceno de las Montes Madonie (Sicilia, Italia Meridional)
El análisis cuantitativo y cualitativo de las asociaciones de foraminíferos aglutinados de aguas profundas (DWAF) de Portella Colla (Montes Madonie) revela una variación del continuo trófico en el Eoceno tardío y el Oligoceno temprano. Se procesaron 29 muestras obteniendo 138 especies de foraminíferos aglutinados, 59 de las cuales se presentan en nomenclatura abierta, pertenecientes a 46 géneros atribuidos a cuatro morfogrupos funcionales según su posición de vida y estrategias de alimentación. La densidad de la fauna aumenta hacia la parte alta de la sección, mientras que la diversidad específica fluctúa y alcanza su mínimo dentro de la transición E/O. En el Eoceno más superior, predominan los especímenes de Cyclamminidae Haplophragmoides y Repmanina charoides, que son formas epifaunales e infaunales inferiores que prefieren el agua de fondo bien oxigenada y un suministro normal de alimento. Dentro de la transición E/O la abundancia de DWAF disminuye ampliamente y las asociaciones están dominadas por taxones oportunistas como Repmanina charoides. En el Rupeliense inferior, los suspensívoros y las asociaciones ricas en Paratrochamminoides están presentes, mientras que los taxones hialinos desaparecen. Al final del Rupeliense inferior, Nothia, los ammodiscidos y los foraminíferos hialinos reaparecen, y el morfogrupo infaunal profundo se convierte en dominante marcando las condiciones de agua del fondo con poco oxígeno disuelto. En el Rupeliense superior, las comunidades infaunales profundas (reofácidos y Caudammina) dominan las asociaciones, y los foraminíferos epifaunales de superficie disminuyen. El LO de Caudammina gutta coincide con un aumento en la deposición de arenas (transición de la Formación Caltavuturo a "Portella Colla Clays"), una disminución en los suspensívoros y los habitantes de la superficie, y el predominio de asociaciones oligotípicas de Reticulophragmium rotundidorsatum. La oxigenación de las aguas de fondo vuelve a niveles similares a los del Eoceno superior, con un suministro de nutrientes medio a alto, como se deduce de la abundancia de Cyclamminidae.
Palabras clave: foraminífero aglutinado; Eoceno; Oligoceno; paleobatimetría; ecología; cambio climático
Traducción: Enrique Peñalver (Sociedad Española de Paleontología)
Résumé en Français
Assemblages éocènes/oligocènes de foraminifères à test agglutiné d'eau profonde des Madonies (Sicile, sud de l'Italie)
Une analyse quantitative et qualitative des assemblages de foraminifères à test agglutiné d'eau profonde (FAEP) de Portella Colla (Madonies) révèle une variation du continuum trophique à l'Éocène récent et l'Oligocène ancien. Vingt-neuf échantillons ont été traités et ont permis d'obtenir 138 foraminifères à test agglutiné (dont 59 sont identifiés en nomenclature ouverte) appartenant à 46 genres attribués à quatre morphogroupes fonctionnels d'après leur position de vie et leur stratégie alimentaire. La densité de faune augmente dans la partie supérieure de la section alors que la diversité spécifique fluctue et atteint son minimum pendant la transition Éocène/Oligocène. Pendant l'Éocène terminal, les cyclamminidés, Haplophragmoides, et Repmanina charoides, des formes épifauniques et endofauniques inférieures qui préfèrent les eaux de fond bien oxygénées et une quantité de nourriture normale, prédominent. Pendant la transition Éocène/Oligocène, l'abondance des FAEP diminue dans l'ensemble et les assemblages sont dominés par des taxons opportunistes tels que Repmanina charoides. Dans le Rupélien inférieur, les suspensivores et les assemblages riches en Paratrochamminoides sont présents, et les taxons à test hyalin disparaissent. À la fin du Rupélien inférieur, Nothia, des ammodiscidés, et des foramminifères à test hyalin réapparaissent, et les morphogroupes endofauniques profonds deviennent dominants, reflétant des eaux de fond pauvres en oxygène. Dans le Rupélien supérieur, les communautés endofauniques profondes (des rhéophacidés et Caudammina) dominent les assemblages, et les foraminifères épifauniques vivant à la surface deviennent moins abondants. La dernière occurrence de C. gutta coïncide avec une augmentation du dépôt de sable (transition de la formation de Caltavuturo vers les argiles de Portella Colla), une diminution des suspensivores et des taxons de surface, et des assemblages oligotypiques dominés par Reticulophragmium rotundidorsatum. L'oxygénation des eaux de fond retourne à des niveaux similaires à ceux de l'Éocène supérieur, avec un approvisionnement en nutriments moyen à élevé déduit de l'abondance des cyclamminidés.
Mots-clés : foraminifère à test agglutiné ; Éocène ; Oligocène ; paléobathymétrie ; écologie ; changement climatique
Translator: Antoine Souron
Deutsche Zusammenfassung
Eozän/oligozäne agglutinierte Tiefwasser-Foraminiferen-Assemblagen aus dem Madonie-Gebirge
(Sizilien, Süditalien)
Quantitative und qualitative Analysen von agglutinierten Tiefwasser-Foraminiferen-Assemblagen aus Portella Colla (Madonie Gebirge) zeigen eine Variation des tropischen Kontinuums im späten Eozän und frühen Oligozän. Es wurden 29 Proben aufgearbeitet die 138 agglutinierte Arten anthielten, von denen 59 in offener Nomenklatur sind und zu 46 Gattungen gehören, die nach ihren Standorten und ihren Fressgewohnheiten vier funktionellen Morphogruppen zugeordnet sind. Die Faunendichte nimmt innerhalb der Sektion nach oben hin zu, wohingegen die spezifische Diversität fluktuiert und das Maximum innerhalb der Eozän/Oligozän Grenze erreicht wird. Im obersten Eozän herrschen Cyclamminiden, Haplophragmoides und Repmanina charoides vor, epifaunale und untere infaunale Formen, die sauerstoffhaltige Tiefenwässer und ein normales Nahrungsangebot bevorzugen. Innerhalb der Eozän/Oligozän Grenze nimmt die Häufigkeit der agglutinierten Tiefwasser-Foraminiferen-Assemblagen stark ab und die Assemblagen werden durch opportunistische Taxa wie Repmanina charoides dominiert. Im unteren Rupelium kommen Suspensionfresser und Assemblagen reich an Paratrochamminoides vor und hyaline Taxa verschwinden. Zum Ende des unteren Rupeliums tauchen Nothia, Ammodisciden und hyaline Foraminiferen wieder auf und tief-infaunale Morphogruppen dominieren, was auf sauerstoffarme Bodenwasser-Konditionen hinweist. Im oberen Rupelium dominieren tief-infaunale Gemeinschaften (Rheophaciden und Caudammina) die Assemblagen und epifaunale an der Oberfläche lebende Foraminiferen nehmen zu. Das LO von Caudammina gutta fällt mit einer Sandzunahme zusammen (Übergang von der Caltavuturo Fm. zu den Portella Colla Tonen), der Abnahme von Suspensionsfressern und Oberflächen-Bewohnern, und der Dominanz von oligotypischen Assemblagen mit Reticulophragmium rotundidorsatum. Die Sauerstoffanreicherung der Tiefenwässer geht zurück auf das Maß wie im oberen Eozän mit einer mittleren bis hohen Nährstoffzufuhr, wie aus dem Reichtum an Cyclamminiden geschlossen werden kann.
Schlüsselwörter: agglutinierte Foraminiferen; Eozän; Oligozän; Paläobathymetrie; Ökologie; Klimawandel
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024