

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
 Simone Colombero. Dipartimento di Scienze della Terra, Università degli Studi di Torino, Via Valperga Caluso 35, 10125 Torino, Italy. simone.colombero@unito.it
Simone Colombero. Dipartimento di Scienze della Terra, Università degli Studi di Torino, Via Valperga Caluso 35, 10125 Torino, Italy. simone.colombero@unito.it
Simone Colombero is a vertebrate paleontologist and researcher at the Dipartimento di Scienze della Terra of the Università di Torino. He received his PhD in 2012 from the Università di Torino. His research is mainly focused on the Neogene mammals of Europe, with particular regards to the systematics, paleobiogeography, paleoecology and biochronology of small mammals (rodents and insectivores), mustelids and artiodactyls.

 David M. Alba. Institut Català de Paleontologia Miquel Crusafont, Universitat Autònoma de Barcelona, Edifici ICTA-ICP, Carrer de les Columnes s/n, Campus de la UAB, E-08193 Cerdanyola del Vallès, Barcelona, Spain. david.alba@icp.cat
David M. Alba. Institut Català de Paleontologia Miquel Crusafont, Universitat Autònoma de Barcelona, Edifici ICTA-ICP, Carrer de les Columnes s/n, Campus de la UAB, E-08193 Cerdanyola del Vallès, Barcelona, Spain. david.alba@icp.cat
David M. Alba (Barcelona, 1975) is a paleontologist working at the Institut Català de Paleontologia Miquel Crusafont, where he is the head of the Neogene and Quaternary Faunas Research Group. He received a PhD in Biology at the Universitat de Barcelona in 2005, with a dissertation focusing on fossil apes. His current research lines are devoted to the paleobiology, systematics and evolution of primates, as well as on the paleobiodiversity of Miocene vertebrate faunas during the Miocene.
 Carmine D’Amico. Dipartimento di Bioscienze e Territorio, Università degli Studi del Molise, Via C. da Fonte Lappone, I-86090 Pesche (Isernia), Italy. crmn.damico@gmail.com
Carmine D’Amico. Dipartimento di Bioscienze e Territorio, Università degli Studi del Molise, Via C. da Fonte Lappone, I-86090 Pesche (Isernia), Italy. crmn.damico@gmail.com
Carmine D'Amico obtained his Master degree in natural sciences at the Sapienza University of Rome and his PhD at the University of Molise. His research is focused on the taxonomy, systematics and palaeoecology of Neogene-Quaternary marine and non-marine mollusks.
 Massimo Delfino. Dipartimento di Scienze della Terra, Università degli Studi di Torino, Via Valperga Caluso 35, 10125 Torino, Italy
Massimo Delfino. Dipartimento di Scienze della Terra, Università degli Studi di Torino, Via Valperga Caluso 35, 10125 Torino, Italy
and Institut Català de Paleontologia Miquel Crusafont, Universitat Autònoma de Barcelona, Edifici ICTA-ICP, Carrer de les Columnes s/n, Campus de la UAB, E-08193 Cerdanyola del Vallès, Barcelona, Spain. massimo.delfino@unito.it
Massimo Delfino is a vertebrate paleontologist whose research activity is focused on the fossil record, morphology, taxonomy and biogeography of amphibians and reptiles. The main research lines are related to the contribution of the fossil record to the phlylogeny of crocodylians and the evolution of the European and Mediterranean herpetological assemblages. He has collaborated to the identification and interpretation of herpetofaunistic assemblages associated to hominoid remains found in Eritrea, Georgia, Indonesia, Kenya, Italy, Oman, and Sudan.
 Daniela Esu. Dipartimento di Scienze della Terra, Sapienza Università di Roma, Piazzale A. Moro 5, I-00185 Roma, Italy. daniela.esu@uniroma1.it
Daniela Esu. Dipartimento di Scienze della Terra, Sapienza Università di Roma, Piazzale A. Moro 5, I-00185 Roma, Italy. daniela.esu@uniroma1.it
Daniela Esu is Associate professor of Palaeontology at the University "Sapienza" of Rome. Her researches focus on systematics, palaeoecology and palaeobiogeography of Cenozoic non-marine molluscs, with special interest to Upper Messinian "Lago-mare" assemblages.
 Piero Giuntelli. Dipartimento di Scienze della Terra, Università degli Studi di Torino, Via Valperga Caluso 35, 10125 Torino, Italy. pierogiuntelli@virgilio.it
Piero Giuntelli. Dipartimento di Scienze della Terra, Università degli Studi di Torino, Via Valperga Caluso 35, 10125 Torino, Italy. pierogiuntelli@virgilio.it
Piero Giuntelli is a paleontologist and independent researcher that collaborates with the Dipartimento di Scienze della Terra of the Università di Torino. He is author of numerous scientific papers mainly focusing on marine and non-marine mollusk faunas from the Neogene of Italy.
 Mathias Harzhauser. Geological-Paleontological Department, Natural History Museum Vienna, Burgring 7, A-1010 Vienna, Austria. mathias.harzhauser@nhm-wien.ac.at
Mathias Harzhauser. Geological-Paleontological Department, Natural History Museum Vienna, Burgring 7, A-1010 Vienna, Austria. mathias.harzhauser@nhm-wien.ac.at
PD Dr. Mathias Harzhauser is head of the Geological-Paleontological Department of the Natural History Museum Vienna (Austria). He is expert in Cainozoic marine and freshwater molluscs. His main interests are the paleogeography and biostratigraphy of the Paratethys Realm and the Oligocene-Miocene biotic development in the eastern Mediterranean and the Indo-Pacific.
 Paul P.A. Mazza. Dipartimento di Scienze della Terra, Università degli Studi di Firenze, Via La Pira 4, 50121 Firenze, Italy. paul.mazza@unifi.it
Paul P.A. Mazza. Dipartimento di Scienze della Terra, Università degli Studi di Firenze, Via La Pira 4, 50121 Firenze, Italy. paul.mazza@unifi.it
Paul Mazza, of the Department of Earth Sciences at the University of Florence, is a paleontologist with expertise in vertebrate paleontology, vertebrate taphonomy, biogeography and paleoclimate. His current research focuses on the paleobiology and taphonomy of continental mammals. Another major research addresses the systematics, evolution and biogeography of insular mammals, with special focus on the patterns and drivers of mammal colonization, diversity and body size on islands.
 Michele Mosca. Dipartimento di Economia e Statistica "Cognetti de Martiis", Università degli Studi di Torino, Lungo Dora Siena 100, 10153 Torino, Italy. michele.mosca@unito.it
Michele Mosca. Dipartimento di Economia e Statistica "Cognetti de Martiis", Università degli Studi di Torino, Lungo Dora Siena 100, 10153 Torino, Italy. michele.mosca@unito.it
Michele Mosca is researcher at LABORatorio R. Revelli and temporary afferent at the Department of Economics and Statistics "Cognetti de Martiis" of University of Turin. He graduated in Statistical, Economic and Managerial Sciences at the University of Turin in 2010, attended the winter school of IRVAPP on "Fundamentals and Methods for Impact Evaluation of Public Policies". He has collaborated with the Labour Market Observatory of the Province of Turin for many years. His main research interests are labour market, pension systems, microsimulation and work and health dynamics. He collaborates with Roberto Leombruni to the development of Whip-Salute and other database derived from administrative data.
 Thomas A. Neubauer. Department of Animal Ecology & Systematics, Justus Liebig Universit, Heinrich-Buff-Ring 26-32 IFZ - 35392 Giessen, Germany. thomas.a.neubauer@allzool.bio.uni-giessen.de
Thomas A. Neubauer. Department of Animal Ecology & Systematics, Justus Liebig Universit, Heinrich-Buff-Ring 26-32 IFZ - 35392 Giessen, Germany. thomas.a.neubauer@allzool.bio.uni-giessen.de
Thomas A. Neubauer is a postdoctoral researcher at the Department of Animal Ecology & Systematics of the Justus Liebig University Giessen, Germany. His current research focus is the reconstruction of diversity and biogeography of freshwater gastropods during the Cenozoic. Above that, his research interests include gastropod taxonomy and nomenclature, as well as morphometric analyses and their applications in evolutionary studies.
 Giulio Pavia. Dipartimento di Scienze della Terra, Università degli Studi di Torino, Via Valperga Caluso 35, 10125 Torino, Italy giulio.pavia@unito.it
Giulio Pavia. Dipartimento di Scienze della Terra, Università degli Studi di Torino, Via Valperga Caluso 35, 10125 Torino, Italy giulio.pavia@unito.it
Giulio Pavia, born February, 1st 1942, is Emeritus Professor of Paleontology at the Dipartimento di Scienze della Terra of the Università di Torino. His researches deal mainly with palaeoecology, stratigraphy, systematics and taphonomy of Middle-to-Upper Jurassic ammonites of Western Tethys, marin molluk fossil assemblages from the Italian Neogene and continental vertebrate assemblages of the Mediterranean Neogene.
 Marco Pavia. Dipartimento di Scienze della Terra, Università degli Studi di Torino, Via Valperga Caluso 35, 10125 Torino, Italy. marco.pavia@unito.it
Marco Pavia. Dipartimento di Scienze della Terra, Università degli Studi di Torino, Via Valperga Caluso 35, 10125 Torino, Italy. marco.pavia@unito.it
Marco Pavia is the Collection Manager of the Department of Earth Science of the Università degli Studi di Torino. He received his PhD in 2000 from the Università of Genova. His research is mainly focused on the systematics, paleoecology and biogeography of Neogene and Pleistocene birds from Europe and Africa.
 Andrea Villa. Dipartimento di Scienze della Terra, Università degli Studi di Torino, Via Valperga Caluso 35, 10125 Torino, Italy. a.villa@unito.it
Andrea Villa. Dipartimento di Scienze della Terra, Università degli Studi di Torino, Via Valperga Caluso 35, 10125 Torino, Italy. a.villa@unito.it
Andrea Villa (Cuggiono, 1990) obtained a Master degree in Natural Sciences at the University of Milan, Italy in 2014. He is currently a PhD student in Earth Sciences at the University of Turin in Italy.
His main research topics are the fossil herpetofaunas of Europe and Africa and the comparative morphology of amphibians and reptiles. In particular, he is now working on the osteological characterization of extant European lizards and the palaeodiversity of these squamates in the continent.
 Giorgio Carnevale. Dipartimento di Scienze della Terra, Università degli Studi di Torino, Via Valperga Caluso 35, 10125 Torino, Italy. giorgio.carnevale@unito.it
Giorgio Carnevale. Dipartimento di Scienze della Terra, Università degli Studi di Torino, Via Valperga Caluso 35, 10125 Torino, Italy. giorgio.carnevale@unito.it
Giorgio Carnevale is Full Professor of Paleontology at the Università degli Studi di Torino. He received his PhD in 2004 from the Università di Pisa. His research primarily focuses on the systematics, paleoecology and biogeography of Mesozoic and Cenozoic teleost fishes.
APPENDIX 1.
Paleoecological requirements of fossil vertebrate taxa of Moncucco Torinese. See text for further discussion, details and references.
FISHES
| Family | Taxon | Ecology |
| Myctophidae | Benthosema aff. suborbitale | marine adventitious |
| Bolinichthys italicus | marine adventitious | |
| Diaphus befralai | marine adventitious | |
| Diaphus aff. pedemontanus | marine adventitious | |
| Diaphus aff. rubus | marine adventitious | |
| Diaphus splendidus | marine adventitious | |
| Diaphus taaningi | marine adventitious | |
| Hygophum aff. derthonensis | marine adventitious | |
| Myctophum coppa | marine adventitious | |
| Gadidae | Gadiculus labiatus | marine adventitious |
| Moridae | Physiculus sp. | marine adventitious |
| Trachichthyidae | Hoplostethus cf. mediterraneus | marine adventitious |
| Bythitidae | Grammonnus sp. | marine adventitious |
| Gobiidae | Lesueurigobius sp. | marine migrant |
| Sciaenidae | Argyrosomus sp. | marine migrant |
| Sciaenidarum sp. nov. | estuarine resident | |
| " Trewasciaena " sp. | estuarine resident |
AMPHIBIANS
APPENDIX 2.
Mollusk and vertebrate taxa identified in the uppermost Miocene of Moncucco Torinese.
APPENDIX 3.
Number of specimens (valves) of bivalves from Moncucco Torinese.
| Taxon | Fossil assemblages | ||||||
| MCC3 | MCC3/4 | MCC4 | MCC4/5 | MCC5 | MCC7 | unassigned | |
| Euxinicardium subodessae | - | - | 3 | 1 | - | - | 3 |
| Pachydacna (Parapachydacna) sp. | - | - | 1 | 1 | - | 2 | 6 |
| Pontalmyra bollenensis | - | 3 | 31 | 3 | - | 25 | 38 |
| Pontalmyra cf. bollenensis | - | 6 | 10 | - | - | - | 2 |
| Pontalmyra cf. incerta chiae | 6 | 11 | 22 | 5 | - | 4 | 9 |
| Pontalmyra partschi | - | - | - | - | - | - | 1 |
| Pontalmyra cf. partschi | 10 | 20 | 151 | 22 | 17 | 43 | 105 |
| Pontalmyra sp. | - | - | - | - | - | - | 2 |
| Prosodacnomya sabbae | - | - | 3 | 1 | - | 2 | 3 |
| Pseudocatillus nevesskayae | - | 2 | 2 | - | - | 5 | 1 |
| Pseudocatillus sp. | - | - | - | - | - | - | 5 |
| cf. Pseudocatillus sp. | - | - | 1 | - | - | - | - |
| Lymnocardiinae gen. et sp. indet. | - | 1 | - | - | - | - | - |
| Dreissena ex gr. rostriformis | 41 | 75 | 499 | 55 | 89 | 112 | 104 |
| Pisidium sp. | - | - | - | - | - | 1 | 4 |
APPENDIX 4.
Osteometrics and odontometrics (mm) of the measurable fossil specimens of perissodactyls, artiodactyls and carnivorans from Moncucco Torinese. Measures were taken following Von den Driesch (1976).
Dihoplus schleiermacheri
| L | B | H | Labs | Lanat | ||
| MGPT-PU 127005 | left M2 | 52 | 53.4? | - | - | - |
| MGPT-PU 127004 | left M3 | - | 55.3 | - | 61 | 57.8 |
| MGPT-PU 130524 | small cuneiform | - | - | 23 | - | - |
Tapirus arvernensis
| L | B | BP | ||
| MGPT-PU 127001 | right m1 | 20 | 15 | - |
| MGPT-PU 130549 | m2 | 20 | 20 | - |
| MGPT-PU 130517 | III Mt | - | - | 22.4? |
Euprox sp.
| L | B | H | BP | DP | DAP | DT | ||
| MGPT-PU 130664 | right C | - | - | - | - | - | 13? | 5 |
| MGPT-PU 130526 | left C | - | - | - | - | - | 6.6 | 3 |
| MGPT-PU 130675 | left d3 | 7.4 | 3.6 | - | - | - | - | - |
| MGPT-PU 130670 | right M1 | 11 | 13 | - | - | - | - | - |
| MGPT-PU 130548 | right M1 | 10 | 12 | - | - | - | - | - |
| MGPT-PU 130665 | right M1 | 11 | 13 | - | - | - | - | - |
| MGPT-PU 130666 | left M1 | 11 | 14 | - | - | - | - | - |
| MGPT-PU 130565 | left M1 | 9 | 10 | - | - | - | - | - |
| MGPT-PU 130668 | left M2 | 13 | 14 | - | - | - | - | - |
| MGPT-PU 130672 | right p2 | 5.1 | 3 | - | - | - | - | - |
| MGPT-PU 130671 | left p3 | 7.7 | 3.8 | - | - | - | - | - |
| MGPT-PU 130674 | left p4 | - | 5.5 | - | - | - | - | - |
| MGPT-PU 130521 | right radius | - | - | - | 23 | 14 | - | - |
| MGPT-PU 130530 | right radius | - | - | - | 19 | 10 | - | - |
| MGPT-PU 130667 | right naviculocuboid | - | - | 15 | - | - | 14 | 18 |
Pliocervus sp .
| Lped | PAD | PTW | DAD | DTW | ||
| MGPT-PU 130574 | left antler | 32.6 | 14.7 | 15.4 | 18.2 | 17 |
Pliocervus sp.
| L | B | |||
| MGPT-PU 130569 | left maxillary | D3 | 12.7 | 8.8 |
| D4 | 12.3 | - | ||
| MGPT-PU 130660 | right P2 | 9.7 | 8.6 | |
| MGPT-PU 130663 | right P2 | 10.9 | 8.2 | |
| MGPT-PU 130662 | right P3 | 10.6 | 10.2 | |
| MGPT-PU 130661 | left P4 | 8.0 | 10.7 | |
| MGPT-PU 130570 | left M1 | 12.3 | 13.7 | |
| MGPT-PU 130566 | right M2 | 12.8 | 15.0 | |
| MGPT-PU 130567 | left M2 | 13.3 | 14.5 | |
| MGPT-PU 130659 | right M3 | 12.6 | 14.9 | |
| MGPT-PU 127008 | right mandible | p2 | 7.8 | 4.4 |
| p3 | 11.1 | 6.7 | ||
| p4 | 10.8 | 7.4 | ||
| m1 | 13 | 8.9 | ||
| m2 | 14 | 9.7 | ||
| m3 | 21.9? | 9.4 | ||
| MGPT-PU 130563 | right mandible | m1 | 12.2 | 8.0 |
| m2 | 13.8 | 9.1 | ||
| MGPT-PU 127023 | left p3 | 10 | 16.9 |
Pliocervus sp.
| L | B | H | LC | DC | BT | Lprox | Ldist | Lsole | LL | ML | BP | Bpa | DP | DL | DM | BS | DS | BD | DD | DAP | DT | ||
| MGPT-PU 130657 | right humerus | - | - | - | - | - | 24.7 | - | - | - | - | - | - | - | - | - | - | - | - | 27.5 | 26.0 | - | - |
| MGPT-PU 130523 | right humerus | - | - | - | - | - | 27.9? | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| MGPT-PU 130656 | left femur | - | - | - | 19.3 | 13.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| MGPT-PU 130557 | right femur | - | - | - | 16 | 13.8 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| MGPT-PU 127011 | right astragalus | 32 | - | - | - | - | - | - | - | - | 32 | 28.7 | 19.4 | 17.3 | - | 16.9 | 17 | - | - | 19.6 | 15.4 | - | - |
| MGPT-PU 130553 | right astragalus | 30 | - | - | - | - | - | - | - | - | 30 | 27.9 | 18.2 | 16.4 | - | 16.5 | 15.6 | - | - | 17.9 | 13.7 | - | - |
| MGPT-PU 130513 | left calcaneum | 62.7 | 21.3 | - | - | - | - | 43.4 | 21.9 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| MGPT-PU 130525 | right calcaneum | - | 22 | 23.3 | - | - | - | - | 23 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| MGPT-PU 127006 | left naviculocuboid | - | - | 20.9 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 21.1 | 25.5 |
| MGPT-PU 127012 | left naviculocuboid | - | - | 21 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 21 | 25 |
| MGPT-PU 130556 | left I cuneiform | - | - | 6.6 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 12.5 | 9.5 |
| MGPT-PU 127010 | right Mt | 183? | - | - | - | - | - | - | - | - | - | - | 21.9 | - | 20.1 | - | - | 13.9 | 14.9 | 23.6 | 15.4 | - | - |
| MGPT-PU 127013 | III phalanx | - | - | - | - | - | - | - | - | 25 | - | - | - | - | - | - | - | - | - | - | - | - | - |
aff. Palaeomeryx
| L | B | ||
| MGPT-PU 130658 | left P4 | 18 | 21 |
| MGPT-PU 127009 | left tibia | - | - |
Gazella aff. pilgrimi
| L | B | |||
| MGPT-PU 130655 | left C | 13.7 | 11 | |
| MGPT-PU 127007 | left mandible | p3 | 10.3 | 7.1 |
| p4 | 10.5 | 7.5 | ||
| m1 | 12.8 | 8.7 | ||
| m2 | 15.2 | 11 | ||
| m3 | 23.5 | 11 | ||
| premolar length | 27.2? | - | ||
| toothrow length | 76.3? | - |
Viverridae gen. et sp. indet.
| L | B | H | BT | BP | DP | BS | DS | BD | BDa | DD | DAP | DT | |||
| MGPT-PU 130563 | left maxillary | P3 | 14 | 9.4 | - | - | - | - | - | - | - | - | - | - | - |
| P4 | 9.6 | 5.7 | - | - | - | - | - | - | - | - | - | - | - | ||
| MGPT-PU 130546 | skull | P4 | 14.6 | 9.5 | - | - | - | - | - | - | - | - | - | - | - |
| M1 | 8.7 | 11.7? | - | - | - | - | - | - | - | - | - | - | - | ||
| MGPT-PU 130652 | left I3 | - | - | - | - | - | - | - | - | - | - | - | 4.6 | 4.5 | |
| MGPT-PU 130550 | right C | - | - | - | - | - | - | - | - | - | - | - | 6.9 | 5.3 | |
| MGPT-PU 130551 | P | 9.1 | 5 | - | - | - | - | - | - | - | - | - | - | - | |
| MGPT-PU 130573 | left C | - | - | - | - | - | - | - | - | - | - | - | 7.1 | 5.1 | |
| MGPT-PU 130654 | right p4 | - | 5.6 | - | - | - | - | - | - | - | - | - | - | - | |
| MGPT-PU 130505 | left humerus | - | - | - | 16.1 | - | - | - | - | 19.1? | - | 13.3 | - | - | |
| MGPT-PU 130572 | left radius | - | - | - | - | - | - | - | - | 16.1 | - | 12.2 | - | - | |
| MGPT-PU 130512 | right III Mc | 39.1 | - | - | - | 7.8 | 7.9 | 4.6 | 3.8 | 8.0 | 6.7 | 6.4 | - | - | |
| MGPT-PU 130503 | left cuboid | - | - | 12.5 | - | - | - | - | - | - | - | - | 9.0 | 10.3 |
Pristifelis attica
| L | BP | DP | BS | DS | BD | BDa | DD | ||
| MGPT-PU 130541 | right IV Mc | 33.2 | 5 | 7 | 4 | 4 | 8 | 6.6 | 6 |
Pantherinae gen. et sp. indet.
| BP | DP | BS | DS | BD | DD | DAP | DT | |||
| MGPT-PU 130545 | left premaxilla | I1 | - | - | - | - | - | - | 4 | 3 |
| I3 | - | - | - | - | - | - | 6.3 | 6 | ||
| MGPT-PU 130543 | right V Mc | - | 12 | 17 | 9 | 9 | - | - | - | - |
| MGPT-PU 130518 | right tibia | - | - | - | - | - | 38 | 24.8 | - | - |
Baranogale cf. helbingi
| L | B | ||
| MGPT-PU 136483 | left M1 | 2.89 | 5.74 |
APPENDIX 5.
Number of identified specimens (NISP) of small mammals (Eulipotyphla, Rodentia and Lagomorpha) from Moncucco Torinese. Chiropterans excluded.
| MCC3 | MCC3/4 | MCC4 | MCC4/5 | MCC5 | MCC7 | TOTAL | |
| Parasorex aff. ibericus | 4 | 1 | 46 | 47 | 22 | 9 | 129 |
| Talpa sp. | 5 | 3 | 3 | 15 | 4 | 3 | 33 |
| Neomyiini indet. | 6 | 9 | 44 | 37 | 22 | 26 | 144 |
| Patenyia cf. hungarica | 0 | 1 | 1 | 0 | 0 | 2 | 4 |
| Castorinae indet | 1 | 0 | 0 | 0 | 0 | 0 | 1 |
| Hystrix depereti | 0 | 0 | 1 | 0 | 0 | 0 | 1 |
| Neocricetodon magnus | 3 | 3 | 9 | 12 | 2 | 6 | 35 |
| Apodemus atavus | 17 | 13 | 47 | 24 | 19 | 16 | 136 |
| Apodemus gudrunae | 53 | 44 | 243 | 118 | 83 | 79 | 620 |
| Centralomys benericettii | 21 | 17 | 103 | 61 | 67 | 87 | 356 |
| Micromys bendai | 6 | 3 | 18 | 15 | 16 | 29 | 87 |
| Occitanomys brailloni | 61 | 50 | 295 | 168 | 120 | 95 | 789 |
| Paraethomys meini | 19 | 21 | 143 | 85 | 73 | 72 | 413 |
| Stephanomys sp. | 1 | 0 | 4 | 0 | 0 | 0 | 5 |
| Muscardinus vireti | 11 | 18 | 74 | 47 | 46 | 35 | 231 |
| Glis minor | 7 | 6 | 15 | 14 | 14 | 5 | 61 |
| Glirulus lissiensis | 0 | 0 | 0 | 2 | 1 | 1 | 4 |
| Eliomys yevesi | 0 | 2 | 6 | 1 | 3 | 7 | 19 |
| Sciurus warthae | 0 | 1 | 0 | 2 | 0 | 2 | 5 |
| Pliopetaurista pliocaenica | 0 | 0 | 0 | 1 | 0 | 0 | 1 |
| Hylopetes hungaricus | 1 | 0 | 0 | 0 | 0 | 0 | 1 |
| Sciurinae indet. | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Prolagus sorbinii | 41 | 43 | 205 | 135 | 111 | 68 | 603 |
| Leporidae indet. | 0 | 0 | 5 | 0 | 1 | 0 | 6 |
| TOTAL | 257 | 235 | 1262 | 784 | 604 | 543 | 3685 |
APPENDIX 6.
Small mammals (Eulipotyphla, Rodentia and Lagomorpha) from Moncucco Torinese based on nNISP (normalized Number of Identified Specimens; see Material and methods for further details). Chiropterans excluded.
| MCC3 | MCC3/4 | MCC4 | MCC4/5 | MCC5 | MCC7 | |
| Parasorex aff. ibericus | 0.095 | 0.024 | 1.095 | 1.119 | 0.524 | 0.214 |
| Talpa sp. | 0.109 | 0.065 | 0.065 | 0.326 | 0.087 | 0.065 |
| Neomyiini indet. | 0.300 | 0.450 | 2.200 | 1.850 | 1.100 | 1.300 |
| Patenyia cf. hungarica | 0.000 | 0.050 | 0.050 | 0.000 | 0.000 | 0.100 |
| Castorinae indet. | 0.050 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| Hystrix depereti | 0.000 | 0.000 | 0.050 | 0.000 | 0.000 | 0.000 |
| Neocricetodon magnus | 0.250 | 0.250 | 0.750 | 1.000 | 0.167 | 0.500 |
| Apodemus atavus | 1.417 | 1.083 | 3.917 | 2.000 | 1.583 | 1.333 |
| Apodemus gudrunae | 4.417 | 3.667 | 20.250 | 9.833 | 6.917 | 6.583 |
| Centralomys benericettii | 1.750 | 1.417 | 8.583 | 5.083 | 5.583 | 7.250 |
| Micromys bendai | 0.500 | 0.250 | 1.500 | 1.250 | 1.333 | 2.417 |
| Occitanomys brailloni | 5.083 | 4.167 | 24.583 | 14.000 | 10.000 | 7.917 |
| Paraethomys meini | 1.583 | 1.750 | 11.917 | 7.083 | 6.083 | 6.000 |
| Stephanomys sp. | 0.083 | 0.000 | 0.333 | 0.000 | 0.000 | 0.000 |
| Muscardinus vireti | 0.688 | 1.125 | 4.625 | 2.938 | 2.875 | 2.188 |
| Glis minor | 0.438 | 0.375 | 0.938 | 0.875 | 0.875 | 0.313 |
| Glirulus lissiensis | 0.000 | 0.000 | 0.000 | 0.125 | 0.063 | 0.063 |
| Eliomys yevesi | 0.000 | 0.125 | 0.375 | 0.063 | 0.188 | 0.438 |
| Sciurus warthae | 0.000 | 0.063 | 0.000 | 0.125 | 0.000 | 0.125 |
| Pliopetaurista pliocaenica | 0.000 | 0.000 | 0.000 | 0.063 | 0.000 | 0.000 |
| Hylopetes hungaricus | 0.063 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| Sciurinae indet. | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.063 |
| Prolagus sorbinii | 1.708 | 1.792 | 8.542 | 5.625 | 4.625 | 2.833 |
| Leporidae indet. | 0.000 | 0.000 | 0.192 | 0.000 | 0.038 | 0.000 |
APPENDIX 7.
Relative abundances of small mammal taxa (Eulipotyphla, Rodentia and Lagomorpha) from Moncucco Torinese based on nNISP and distribution of their habitat requirements used to calculate variation of the habitat spectrum of Moncucco Torinese. Chiropterans excluded. Further details in Material and methods. See also Blain et al. (2008) and Piñero et al. (2015).
| Relative Abundances (based on nNISP) | Distribution of habitat requirements (see Piñero et al., 2015) | ||||||||||||
| MCC3 | MCC3/4 | MCC4 | MCC4/5 | MCC5 | MCC7 | Closed canopy forests | Woodlands / bushlands | Grass lands | Rocky outcrops | Sandy/soft substrate | Water edges | Unknown / eurytopic | |
| Parasorex aff. ibericus | 0.005 | 0.001 | 0.012 | 0.021 | 0.013 | 0.005 | - | - | - | - | - | - | 1 |
| Talpa sp. | 0.006 | 0.004 | 0.001 | 0.006 | 0.002 | 0.002 | - | - | - | - | 1 | - | - |
| Neomyiini indet. | 0.016 | 0.027 | 0.025 | 0.035 | 0.026 | 0.033 | - | - | - | - | - | 1 | - |
| Patenyia cf. hungarica | 0.000 | 0.003 | 0.001 | 0.000 | 0.000 | 0.003 | - | - | - | - | - | - | 1 |
| Castorinae indet. | 0.003 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | - | - | - | - | - | 1 | |
| Hystrix depereti | 0.000 | 0.000 | 0.001 | 0.000 | 0.000 | 0.000 | - | - | - | - | - | - | 1 |
| Neocricetodon magnus | 0.014 | 0.015 | 0.008 | 0.019 | 0.004 | 0.013 | - | - | - | - | - | - | 1 |
| Apodemus atavus | 0.077 | 0.066 | 0.044 | 0.038 | 0.038 | 0.034 | 0.5 | 0.5 | - | - | - | - | - |
| Apodemus gudrunae | 0.240 | 0.222 | 0.227 | 0.186 | 0.166 | 0.167 | - | 0.5 | - | 0.5 | - | - | - |
| Centralomys benericettii | 0.095 | 0.086 | 0.096 | 0.096 | 0.134 | 0.184 | - | 0.5 | 0.5 | - | - | - | - |
| Micromys bendai | 0.027 | 0.015 | 0.017 | 0.024 | 0.032 | 0.061 | - | - | 1 | - | - | - | - |
| Occitanomys brailloni | 0.276 | 0.252 | 0.275 | 0.265 | 0.240 | 0.201 | - | 1 | - | - | - | - | - |
| Paraethomys meini | 0.086 | 0.106 | 0.133 | 0.134 | 0.146 | 0.152 | - | 0.5 | 0.5 | - | - | - | - |
| Stephanomys sp. | 0.005 | 0.000 | 0.004 | 0.000 | 0.000 | 0.000 | - | - | - | - | - | - | 1 |
| Muscardinus vireti | 0.037 | 0.068 | 0.052 | 0.056 | 0.069 | 0.055 | 0.5 | 0.5 | - | - | - | - | - |
| Glis minor | 0.024 | 0.023 | 0.010 | 0.017 | 0.021 | 0.008 | 1 | - | - | - | - | - | - |
| Glirulus lissiensis | 0.000 | 0.000 | 0.000 | 0.002 | 0.001 | 0.002 | 1 | - | - | - | - | - | - |
| Eliomys yevesi | 0.000 | 0.008 | 0.004 | 0.001 | 0.004 | 0.011 | - | 0.5 | - | 0.5 | - | - | - |
| Sciurus warthae | 0.000 | 0.004 | 0.000 | 0.002 | 0.000 | 0.003 | 1 | - | - | - | - | - | - |
| Pliopetaurista pliocaenica | 0.000 | 0.000 | 0.000 | 0.001 | 0.000 | 0.000 | 1 | - | - | - | - | - | - |
| Hylopetes hungaricus | 0.003 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 1 | - | - | - | - | - | - |
| Sciurinae indet. | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.002 | 1 | - | - | - | - | - | - |
| Prolagus sorbinii | 0.086 | 0.100 | 0.088 | 0.098 | 0.102 | 0.066 | - | - | - | - | - | - | 1 |
| Leporidae indet. | 0.000 | 0.000 | 0.002 | 0.000 | 0.001 | 0.000 | - | - | - | - | - | - | 1 |
APPENDIX 8.
Values of beta coefficients figured in Figure 17 and their statistical significance assessed by computing Wald Chi-square tests (small characters below the value of the coefficient) of the binary logistic regression models estimating the log odds of finding a murid species at Moncucco Torinese with MCC3 as reference variable.
| Constant | MCC3_4 | MCC4 | MCC4_5 | MCC5 | MCC7 | |
| Apodemus atavus | -2.2481 | -0.0922 | -0.5938 | -0.6764 | -0.6907 | -0.8709 |
| <.0001 | 0.8115 | 0.0448 | 0.0404 | 0.0466 | 0.0158 | |
| Apodemus gudrunae | -0.858 | -0.002 | -0.062 | -0.238 | -0.410 | -0.473 |
| <.0001 | 0.993 | 0.730 | 0.224 | 0.046 | 0.022 | |
| Centralomys benericettii | -2.012 | -0.030 | 0.026 | 0.106 | 0.477 | 0.804 |
| <.0001 | 0.931 | 0.918 | 0.693 | 0.076 | 0.002 | |
| Micromys bendai | -3.356 | -0.522 | -0.481 | -0.059 | 0.237 | 0.868 |
| <.0001 | 0.466 | 0.315 | 0.905 | 0.627 | 0.058 | |
| Occitanomys brailloni | -0.651 | -0.022 | 0.014 | 0.062 | -0.114 | -0.440 |
| <.0001 | 0.927 | 0.936 | 0.739 | 0.554 | 0.026 | |
| Paraethomys meini | -2.125 | 0.325 | 0.522 | 0.611 | 0.695 | 0.678 |
| <.0001 | 0.337 | 0.044 | 0.024 | 0.012 | 0.014 | |
| Stephanomys sp. | -5.176 | -12.001 | -0.182 | -12.001 | -12.001 | -12.001 |
| <.0001 | 0.978 | 0.871 | 0.961 | 0.965 | 0.965 |
APPENDIX 9
Frequencies of different habitat systems in each fossil assemblage of Moncucco Torinese (see Figure 18). According to the Habitat Weightings Method (Blain et al., 2008), the values are based on the small mammal relative abundances within each fossil assemblage and on their habitat preference as indicated in Appendix 7. See also Material and methods.
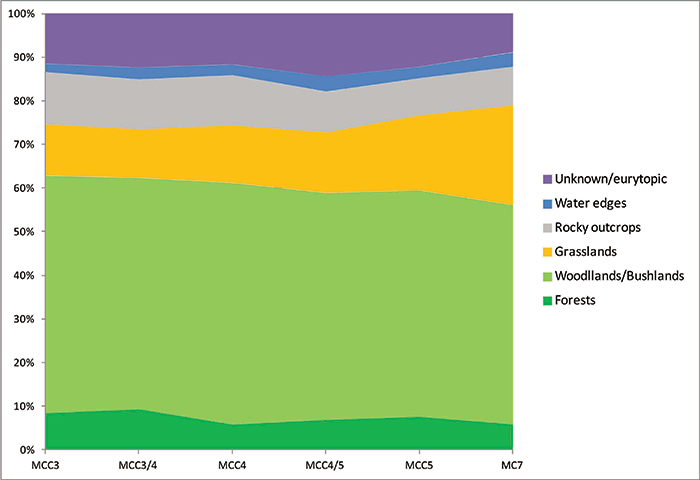
| Forests | Woodlands/ Bushlands | Grasslands | Rocky outcrops | Sandy/soft substrate | Water edges | Unknown/ eurytopic | |
| MCC3 | 0.084 | 0.544 | 0.118 | 0.120 | 0.006 | 0.019 | 0.109 |
| MCC3/4 | 0.093 | 0.530 | 0.111 | 0.115 | 0.004 | 0.027 | 0.120 |
| MCC4 | 0.058 | 0.553 | 0.132 | 0.115 | 0.001 | 0.025 | 0.116 |
| MCC4/5 | 0.069 | 0.520 | 0.139 | 0.093 | 0.006 | 0.035 | 0.138 |
| MCC5 | 0.076 | 0.519 | 0.172 | 0.085 | 0.002 | 0.026 | 0.120 |
| MC7 | 0.059 | 0.502 | 0.229 | 0.089 | 0.002 | 0.033 | 0.087 |
APPENDIX 10.
Values of the CRI (Climatic Restriction Index) of each rodent species in each climatic zones and bioclimatic characterization of Moncucco Torinese according to the methods of Hernández Fernández (2001), Hernández Fernández and Peláez-Campomanes (2003) and Hernández Fernández et al. (2007) with modifications (see material and methods). Following Hernández Fernández et al. (2007), the climate zone and the main vegetational types (in brackets) are defined as follows: I: Equatorial (Evergreen tropical rainforests); II: Tropical with summer rains (Tropical deciduous forests); II/III: Transition tropical semiarid (savannah-like); III: Subtropical arid (subtropical desert); IV: Subtropical with winter rains and summer drought/Mediterranean (Sclerophyllous woodland-shrubland); V: Warm-temperate (Temperate evergreen forests); VI: Typical temperate (Nemoral broadleaf-deciduous forest); VII: Arid-temperate (Steppe to cold desert); VIII: Cold Temperate/boreal (Boreal coniferous forest/taiga); IX: Polar (tundra).
| I | II | II/III | III | IV | V | VI | VII | VIII | IX | |
| Castorinae indet. | - | - | - | - | 0.200 | 0.200 | 0.200 | 0.200 | 0.200 | - |
| Hystrix depereti | 0.143 | 0.143 | 0.143 | 0.143 | 0.143 | 0.143 | - | 0.143 | - | - |
| Neocricetodon magnus | - | - | - | - | 0.333 | - | 0.333 | 0.333 | - | - |
| Apodemus atavus | - | - | - | - | 0.250 | - | 0.250 | 0.250 | 0.250 | - |
| Apodemus gudrunae | - | - | - | - | 1.000 | - | - | - | - | - |
| Centralomys benericettii | - | 0.333 | 0.333 | - | 0.333 | - | - | - | - | - |
| Micromys bendai | - | - | - | - | - | - | 0.500 | - | 0.500 | - |
| Occitanomys brailloni | - | 1.000 | - | - | - | - | - | - | - | - |
| Paraethomys meini | - | 0.250 | 0.250 | 0.250 | 0.250 | - | - | - | - | - |
| Stephanomys sp. | - | 0.333 | 0.333 | 0.333 | - | - | - | - | - | |
| Muscardinus vireti | - | - | - | - | 0.500 | - | 0.500 | - | - | - |
| Glis minor | - | - | - | - | 0.500 | - | 0.500 | - | - | - |
| Glirulus lissiensis | - | - | - | - | - | 1.000 | - | - | - | - |
| Eliomys yevesi | - | - | - | 0.500 | - | 0.500 | - | - | - | |
| Sciurus warthae | - | - | - | - | 0.333 | - | 0.333 | - | 0.333 | - |
| Pliopetaurista pliocaenica | 0.200 | 0.200 | - | - | - | 0.200 | 0.200 | - | 0.200 | - |
| Hylopetes hungaricus | 1.000 | - | - | - | - | - | - | - | - | - |
| Sciurinae indet. | 0.167 | 0.167 | - | - | 0.167 | 0.167 | 0.167 | - | 0.167 | - |
| Bioclimatic spectrum | 8.386 | 13.479 | 5.886 | 2.183 | 26.905 | 9.497 | 19.352 | 5.146 | 9.167 | 0.000 |
TABLE 1. Chi-square tests evaluating differences between the samples of murids of the different fossil assemblages from MCC. In light gray the moderately significant differences (0.05<p<0.1), in dark grey the highly significant differences (p<0.05); df: degrees of freedom; 5df when Stephanomys sp. is absent in both layers.
| MCC3 | MCC3/4 | MCC4 | MCC4/5 | MCC5 | MCC7 | |||||||
| MCC3 | - | Χ2 =2.2377 | df:6 | Χ2 =8.4798 | df:6 | Χ2 =12.401 | df:6 | Χ2 =17.58 | df:6 | Χ2 =32.087 | df:6 | |
| p=0.897 | p=0.205 | p=0.053 | p=0.007 | p<0.001 | ||||||||
| MCC3/4 | Χ2 =2.2377 | df:6 | - | Χ2 =3.5915 | df:5 | Χ2 =5.3069 | df:5 | Χ2 =10.852 | df:5 | Χ2 =24.571 | df:5 | |
| p=0.90 | p=0.732 | p=0.380 | p=0.055 | p<0.001 | ||||||||
| MCC4 | Χ2 =8.4798 | df:6 | Χ2 =3.5915 | df:5 | - | Χ2 =5.604 | df:6 | Χ2 =18.06 | df:6 | Χ2 =58.075 | df:6 | |
| p=0.21 | p=0.732 | p=0.469 | p=0.006 | p<0.001 | ||||||||
| MCC4/5 | Χ2 =12.401 | df:6 | Χ2 =5.3069 | df:5 | Χ2 =5.604 | df:6 | - | Χ2 =5.789 | df:5 | Χ2 =29.853 | df:5 | |
| p=0.053 | p=0.380 | p=0.469 | p=0.328 | p<0.001 | ||||||||
| MCC5 | Χ2 =17.58 | df:6 | Χ2 =10.852 | df:5 | Χ2 =18.06 | df:6 | Χ2 =5.789 | df:5 | - | Χ2 =9.6227 | df:5 | |
| p=0.007 | p=0.055 | p=0.006 | p=0.328 | p=0.087 | ||||||||
| MCC7 | Χ2 =32.087 | df:6 | Χ2 =24.571 | df:5 | Χ2 =58.075 | df:6 | Χ2 =29.853 | df:5 | Χ2 =9.6227 | df:5 | - | |
| p<0.001 | p<0.001 | p<0.001 | p<0.001 | p=0.087 | ||||||||
TABLE 2. Number of specimens and taxa of small mammals found at Moncucco Torinese with value of the diversity (Shannon) index for each studied sample. Chiropterans are excluded.
| MCC3 | MCC3/4 | MCC4 | MCC4/5 | MCC5 | MCC7 | Total | |
| Specimens (N) | 257 | 235 | 1262 | 784 | 604 | 543 | 3685 |
| Taxa (N) | 16 | 16 | 18 | 17 | 16 | 18 | 24 |
| Shannon (H’) | 2.094 | 2.141 | 2.063 | 2.177 | 2.151 | 2.193 | 2.157 |
TABLE 3. Estimated paleoclimatic values of MCC according to three different methodologies based on small mammals (see Material and methods for further details). MAP: Mean Annual Precipitation; MINP: Precipitation in the driest month; MAT: Mean Annual Temperature; MTW: Mean Temperature of the Warmest month; MTC: Mean Temperature of the Coldest month; SE: Standard Error.
| Methodology | Hernández Fernández (2001) | Van Dam (2006) | Montuire et al. (2006) | |||
| Proxy | Rodents | Small mammals | Murids | |||
| Results | SE | Results | SE | Results | SE | |
| MAP | 967.9 mm | ±470.6 mm | 849.8 mm | ±350-400 mm | - | - |
| MINP | - | - | 41.5 mm | ±17-19 mm | - | - |
| MAT | 16.2° C | ±3.6° C | - | - | 17.2° C | ±4.8°C |
| MTW | 24.4° C | ±4.7° C | - | - | - | - |
| MTC | 8.2° C | ±5.1° C | - | - | - | - |
FIGURE 1. Geographic and stratigraphic setting of the Tertiary Piedmont Basin (TPB). The star indicates the geographic positions of the fossiliferous site of Moncucco Torinese (MCC). Modified from Lozar et al. (2015).
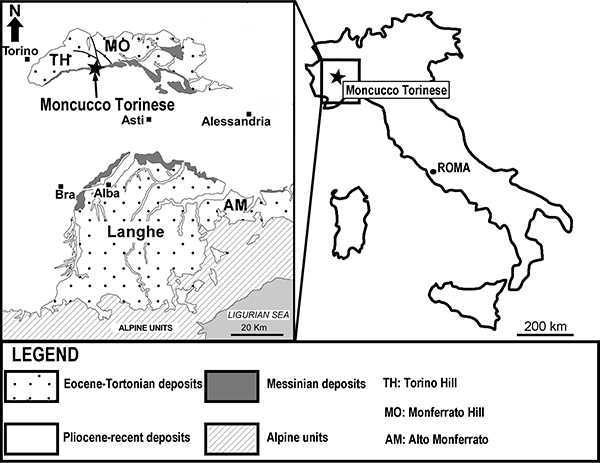
FIGURE 2. Stratigraphic scheme of the Messinian succession of the Tertiary Piedmont Basin with the stratigraphic log of the studied section of Moncucco Torinese section. Fossiliferous assemblages are indicated with the abbreviation MCC. The arrows indicate the main stratigraphic event recorded in the section. For further details see Dela Pierre et al. (2007) and Angelone et al. (2011).
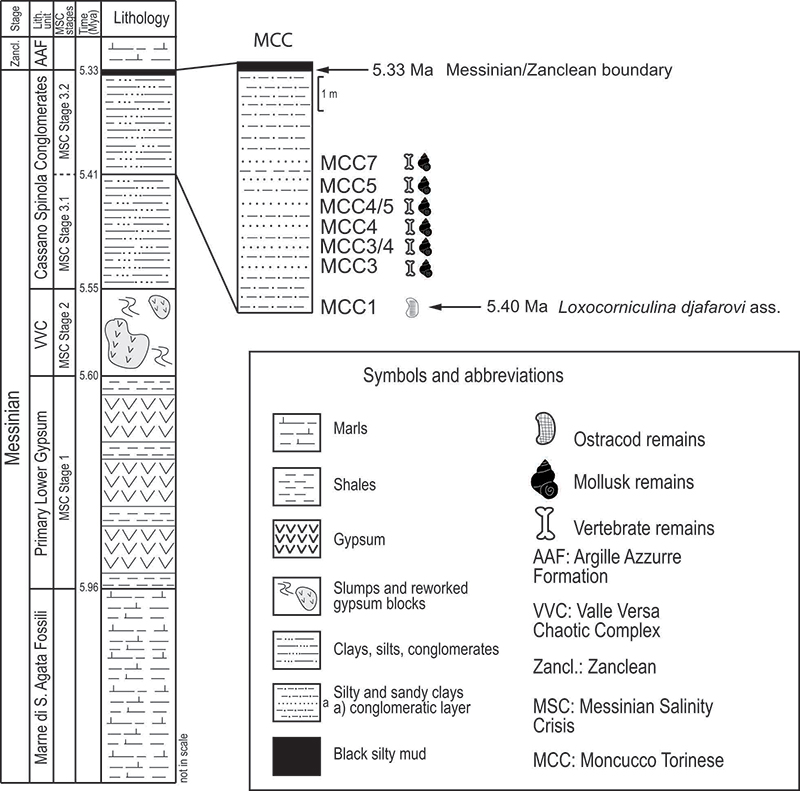
FIGURE 3. Gastropoda from Moncucco Torinese. 1-2. Cochlostomaesuanum: 1, lateral view of apex and protoconch (MGPT-PU 110978); 2: apertural view (MGPT-PU 110979). 3. ? Limax fossilis: dorsal view (MGPT-PU 110928). 4. Parmacella sp.: dorsal view (MGPT-PU 110931). 5. Lucilla miocaenica: apical view of paratype (MGPT-PU 108849). 6.Truciella ballesioi : fragment with aperture in apertural view (MGPT-PU 108851). 7. Nordsieckia pontica: apertural view (MGPT-PU 108848). 8. Acicula giuntellii: apertural view of holotype (MGPT-PU 108896). 9. Theodoxus doderleini : dorsal view (MGPT-PU 110959). 10.Theodoxus mutinensis: dorsal view (MGPT-PU 110962). 11. Melanopsis narzolina: apertural view (MGPT-PU 110967). 12. Melanoides curvicosta: apertural view (MGPT-PU 110965). 13. Saccoia cf. congermana: apertural view (MGPT-PU 108873). 14. Saccoia globosa: apertural view of holotype (MGPT-PU 108858). Scale bars equal 0.5 mm (1-2, 5-7) and 1 mm (3-4, 8-14).

FIGURE 4. Bivalvia from Moncucco Torinese. 1-4. Euxinicardium subodessae: external (1) and internal (2) views of LV (MGPT-PU 130575), external (3) and internal (4) views of RV (MGPT-PU 130576). 5. Pachydacna (Parapachydacna) sp.: umbonal view of RV (MGPT-PU 130577). 6-8. Pontalmyra bollenensis: 6, external view of RV (MGPT-PU 130578). 5, external (7) and internal (8) views of LV (MGPT-PU 130579). 9-10. Pontalmyra cf. incerta chiae: external (9) and internal (10) views of RV (MGPT-PU 130580). 11-12. Pontalmyra cf. partschi: external (11) and internal (12) views of RV (MGPT-PU 130581). 13-14. Prosodacnomya sabbae: external (13) and internal (14) views of RV (MGPT-PU 130582). 15-16. Pseudocatillus nevesskayae: external (15) and internal (16) views of RV (MGPT-PU 130583). 17-18. Pseudocatillus sp.: external (17) and internal (18) views of RV (MGPT-PU 130584). 19-20. Dreissena ex gr. rostriformis: external (19) and internal (20) views of RV, (MGPT-PU 130585). Scale bars equal 1 mm (1-5, 9-12, 15-18) and 5 mm (6-8, 13-14, 19-20). LV: left valve, RV: right valve.

FIGURE 5. Fish otoliths from Moncucco Torinese. 1.Benthosema aff. suborbitale (MGPT-PU 130304). 2. Diaphus befralai (MGPT-PU 130320). 3. Diaphus aff. rubus (MGPT-PU 130293). 4. Diaphus taaningi (MGPT-PU 130253). 5. Diaphus aff. pedemontanus (MGPT-PU 130288). 6. Diaphus splendidus (MGPT-PU 130308). 7. Hygophum aff. derthonensis (MGPT-PU 130275). 8. Myctophum coppa (MGPT-PU 130280). 9. Gadiculus labiatus (MGPT-PU 130324). 10. Physiculus sp. (MGPT-PU 130265). 11. Grammonus sp. (MGPT-PU 130273). 12. Hoplostethus cf. mediterraneus (MGPT-PU 130232). 13. Lesueurigobius sp. (MGPT-PU 130330). 14. “Trewasciaena” sp. (MGPT-PU 130246). 15. Sciaenidarum sp. nov. (MGPT-PU 130241). 16. Sciaenidarum sp. nov. (MGPT-PU 130263). 17. Argyrosomus sp. (MGPT-PU 130264). Scale bars: 1 mm.
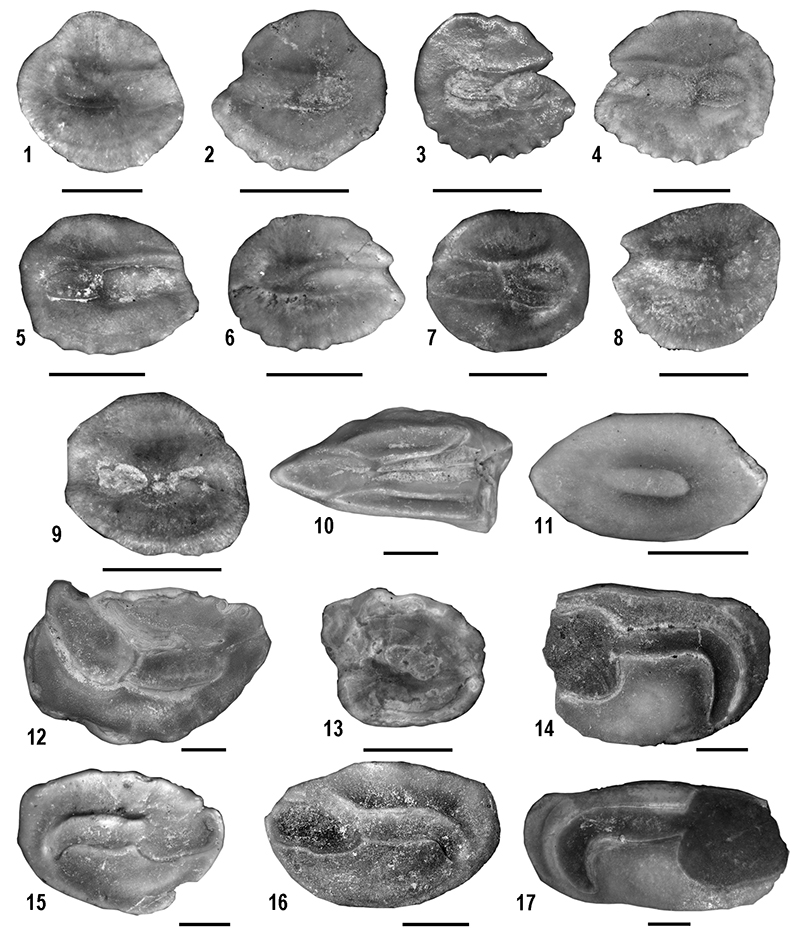
FIGURE 6. Amphibians from Moncucco Torinese. 1-2. Albanerpeton sp.: right dentary (MGPT-PU 132003) in medial (1) and dorsal (2) views. 3-5. Chelotriton sp.: trunk vertebra (MGPT-PU 132302) in anterior (3), dorsal (4) and left lateral (5) views. 6-7. Lissotriton sp.: trunk vertebra (MGPT-PU 132306) in dorsal (6) and posterior (7) views. 8. Bufo gr. B. viridis: left ilium (MGPT-PU 132177) in lateral view. 9. Pelophylax sp.: right ilium (MGPT-PU 132317) in lateral view. 10. Hyla gr. H. arborea: left ilium (MGPT-PU 132201) in lateral view. 11-12. Latonia sp.: left angular (MGPT-PU 132314) in ventral (11) and dorsal (12) views. 13-14. Pelobates sp.: left ilium (MGPT-PU 132308) in medial (13) and lateral (14) views. Scale bars equal 1 mm.

FIGURE 7. Reptiles from Moncucco Torinese. 1-2. Crocodylia indet.: isolated tooth (MGPT-PU 132316) in labial (1) and mesial (2) views. 3-4. Testudo sp: eight peripheral (MGPT-PU 132048) in external view (3) and pygal (MGPT-PU 132050) connected to the eleventh peripherals (MGPT-PU 132051-52) in external view (4). 5-8. Mauremys sp.: entoplastron (MGPT-PU 132059) in ventral view (5), right epiplastron (MGPT-PU 132320) in ventral view (6), pygal (MGPT-PU 132057) in external view (7) and tenth peripheral (MGPT-PU 132342) in external view (8). 9. cf. Euleptes sp.: left maxilla (MGPT-PU 132432) in lateral view. 10. Agamidae indet.: tooth bearing bone (MGPT-PU 132438) in lateral view. 11-12. non- Anguis Anguinae indet.: vertebra (MGPT-PU 132612) in dorsal (11) and ventral (12) views. 13-16. cf. Lacerta sp.: right dentary (MGPT-PU 132501) in lateral (13) and medial (14) views and fused frontals (MGPT-PU 132532) in dorsal (15) and ventral (16) views. 17-19. Amphisbaenia indet.: trunk vertebra (MGPT-PU 132593) in dorsal (17), ventral (18), and left lateral (19) views. 20. Scolecophidia indet.: trunk vertebra (MPUT-PU 132027) in left lateral view. 21-22. Eryx sp.: caudal vertebra (MGPT-PU 132626) in anterior (21) and left lateral (22) views; 23-25. Colubrines A: trunk vertebra (MPUT-PU 132662) in dorsal (23), ventral (24), and posterior (25) views. 26-28. Colubrines B: trunk vertebra (MPUT-PU 132663) in dorsal (26), ventral (27), and anterior (28) views. 29-30. Vipera sp. (Oriental group): trunk vertebra (MPUT-PU 1320661) in left lateral (29) and posterior (30) views. Scale bars equal 1 mm (1-2, 9-30) and 10 mm (3-6)

FIGURE 8. Aves from Moncucco Torinese. 1. Palaeortyx cf. gallica: right carpometacarpus (MGPT-PU 127032), ventral view. 2. Coturnix sp.: left coracoid (MGPT-PU 130574), dorsal view. 3-4. Coliidae gen. et sp. indet.: right tarsometatarsus (MGPT-PU 127020), dorsal (3) and distal (4) views. 5,7,9. Strigidae gen. et sp. indet.: 5, right tibiotarsus (MGPT-PU 127018), cranial view; right tarsometatarsus (MGPT-PU 127019) in proximal (7) and dorsal (9) views. 6. Columbidae gen. et sp. indet.: right ulna (MGPT-PU 127017), ventral view. 8. Apodidae gen. et sp. indet.: left scapula (MGPT-PU 127014), lateral view. Scale bars equal 5 mm.
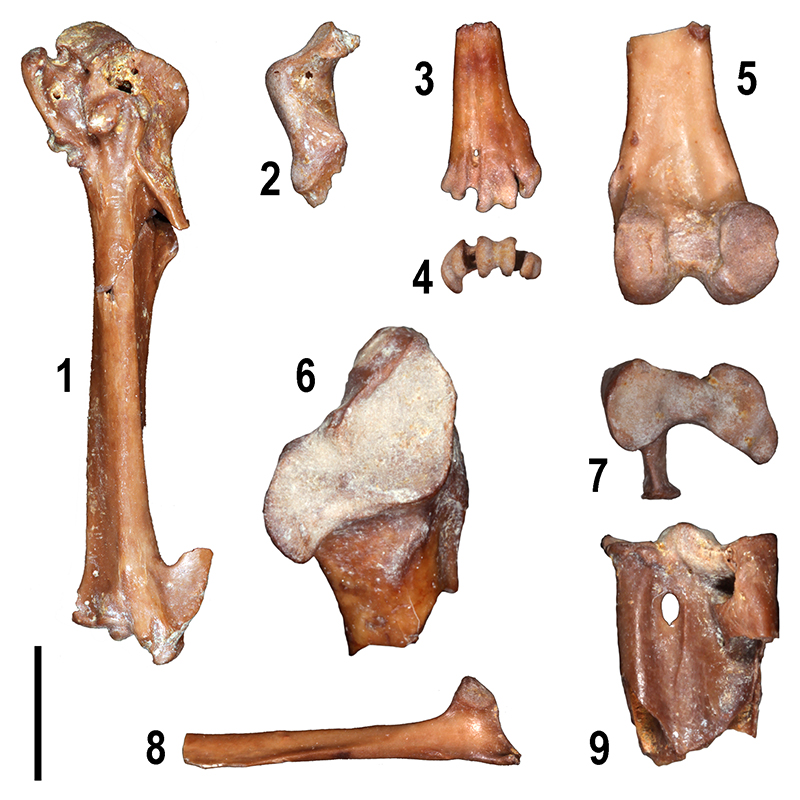
FIGURE 9. Large mammal remains from Moncucco Torinese. 1-2. Dihoplus schleiermacheri: 1, left upper second molar (MGPT-PU 127005), occlusal view; 2, left upper third molar (MGPT-PU 127004), occlusal view. 3. Dicoryphochoerini gen. et sp. indet.: right lower third premolar (MGPT-PU 130571), buccal view. 4-5. Tapirus arvernensis : 4, right lower molar (MGPT PU 130549), lingual (4) and occlusal (5) views. 6-7, 20-21. Gazella aff. G. pilgrimi: left hemimandible (MGPT-PU 125007), buccal (6) and occlusal (7) views; left maxillary fragment (MGPT-PU 130655), lingual (20) and buccal (21) views. 8-9, 12-13, 17. Pliocervus sp.: right hemimandible (MGPT PU 125008), buccal (8) and occlusal (9) views; right upper third premolar (MGPT-PU 130659), occlusial (12) and mesial (13) views; right metatarsal bone (MGPT-PU 127010), plantar view (17). 10-11, 18. aff. Palaeomeryx: left upper fourth premolar (MGPT-PU 130658), occlusal (10) and mesial (11) views; 18, left tibia (MGPT-PU 127009) dorsal view. 14-16, 19. Euprox sp.: left upper second molar (MGPT-PU 130668), occlusal 14) and mesial (15) views; 16, left lower third premolar (MGPT-PU 130671), occlusial view; 19, right upper canine (MGPT-PU 130664), buccal view. Scale bars equal 10 mm.
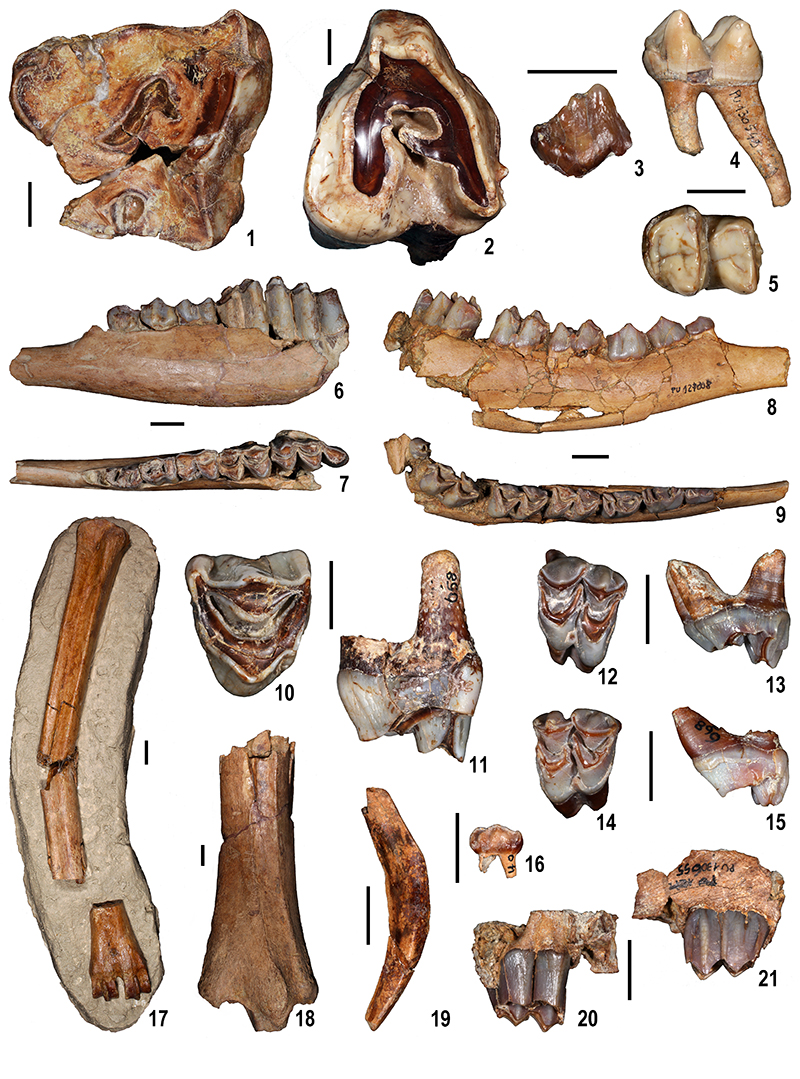
FIGURE 10. Large mammal remains from Moncucco Torinese. 1-2. Pliocervus sp.: fragmentary skull (MGPT-PU 130574), medial (1) and lateral (2) views. Scale bar equals 10 mm.

FIGURE 11. Large mammal remains from Moncucco Torinese. 1, 6-7, 10-13. Viverridae gen. et sp., indet.: fragmentary skull (MGPT-PU 130546), palatal view (1) and detail of P4 and M1 in occlusal view (6); 7, left upper canine (MGPT-PU 130573), buccal view; right lower fourth premolar (MGPT-PU 130654), occlusal (10) and buccal (11) views; 12, left radius (MGPT-PU 130572), dorsal view; 13, right metacarpal III (MGPT-PU 130512), dorsal view. 2-3. Euarctos pyrenaicus: left upper carnassial (MGPT-PU 130529), lingual (2) and occlusal (3) views. 4-5, 8-9. Pantherinae gen. et sp. indet.: left premaxilla (MGPT-PU 130545), lateral (4) and ventral (5) views; right tibia (MGPT-PU 130518), dorsal (8) and distal (9) views. 14. cf. Mesopithecus pentelicus pentelicus: right lower canine (MGPT-PU 130502). 15. cf. Macaca sp.: left lower third molar (MGPT-PU 130506). The figures 14 and 15 are from Alba et al., (2014). Scale bars equal 10 mm.
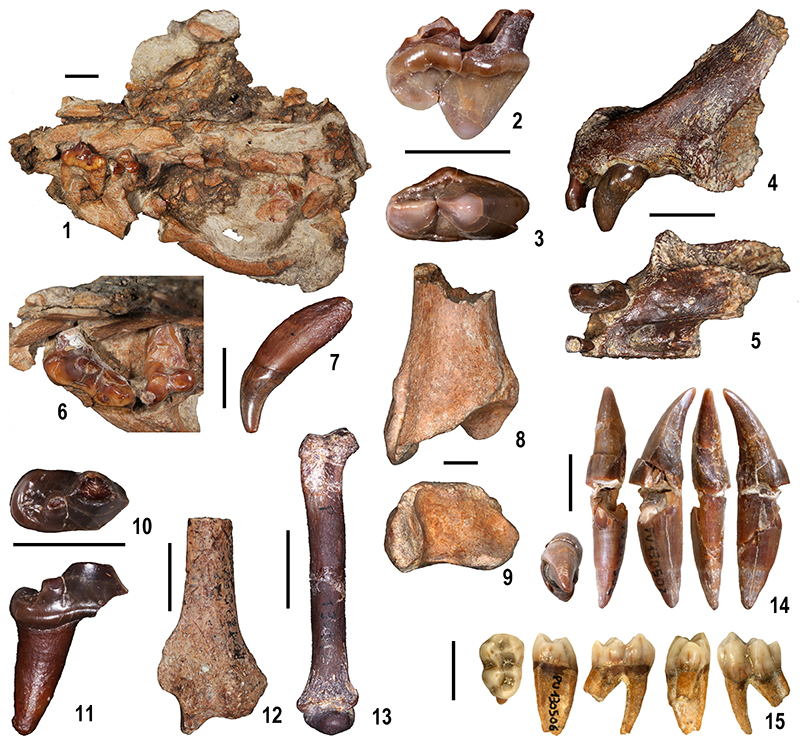
FIGURE 12. 1-3. Baranogale cf. helbingi from Moncucco Torinese. Left M1 (MGPT-PU136483). Mesial (reversed) (1), occlusal (2) and distal (3) views. Scale bar equals 1 mm.
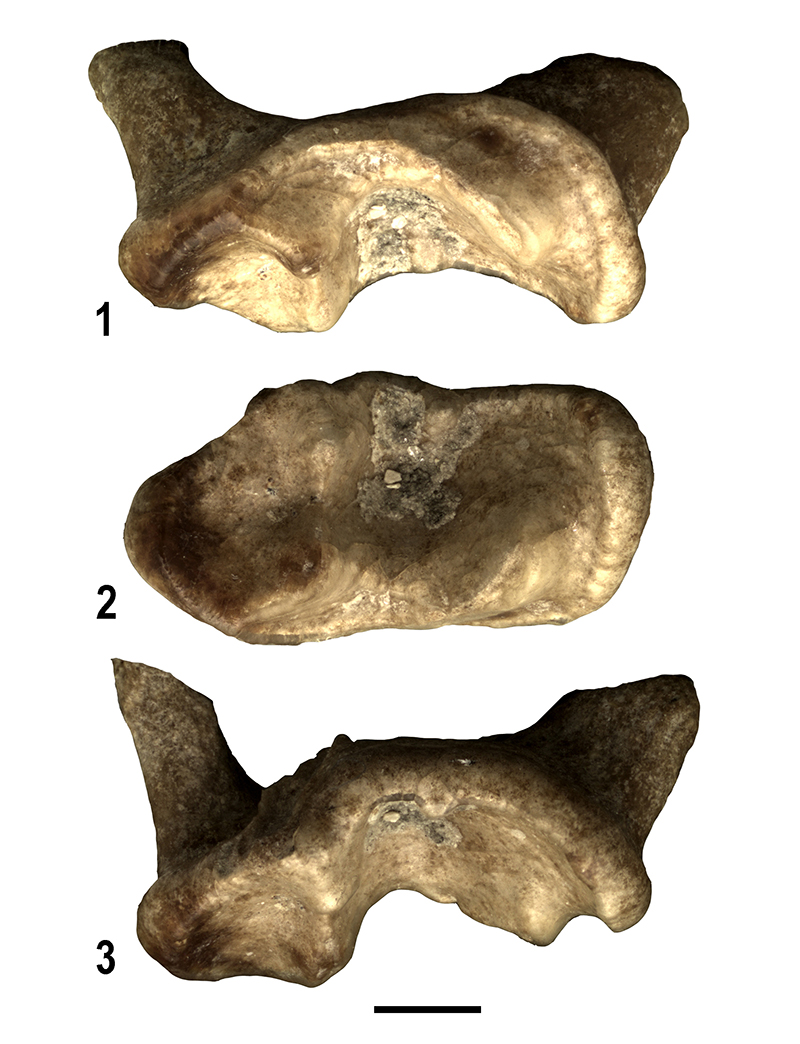
FIGURE 13. Small mammals from Moncucco Torinese. 1. Stephanomy s sp.: left M1 (MGPT-PU 136460). 2. Occitanomys brailloni, left M1 (MGPT-PU 136607). 3. Paraethomys meini: right M1(MGPT-PU 136672). 4. Apodemus gudrunae: right M1 (MGPT-PU 136375). 5. Apodemus atavus: leftM1 (MGPT136898). 6. Micromys bendai: left M1 (MGPT-PU 136932) 7. Centralomys benericettii: left M1 (MGPT-PU 128090). 8. Neocricetodon magnus: left M2 (MGPT-PU136481). 9. Glirulus lissiensis : right m3 (MGPT-PU 136523). 10. Eliomys yevesi : left M2 (MGPT-PU 136582). 11. Muscardinus vireti : right m2 (MGPT-PU136504). 12. Glis minor: left M1 (MGPT-PU136555). 13. Pliopetaurista pliocaenica: left m1 (MGPT-PU 128218). 14. Hylopetes hungaricus: left M1 (MGPT-PU 136589). 15. Sciurinae indet.: left m1-2 (MGPT-PU 136590). 16. Sciurus warthae: right m1 (MGPT-PU 128221). 17. Talpa sp.: right P4 (MGPT-PU 131249). 18. Petenyia cf. hungarica: left m2 (MGPT-PU 131388). 19. Neomyini indet.: left emimandible (MGPT-PU 131123). 20. Parasorex aff. ibericus: right M2 (MGPT-PU 131025). 21. Leporidae indet.: right upper molariform (MGPT-PU 136477). 22. Prolagus sorbinii: right P4 (MGPT-PU 136475). 23. Castorinae indet.: left upper incisor in distal view (MGPT-PU 136482). 24. Hystrix depereti: right radius in ventral view (MGPT-PU 128349). Scale bars equal 1 mm (1-22) and 10 mm (23-24). Figures 13-14 and 16 from Colombero and Carnevale (2016).
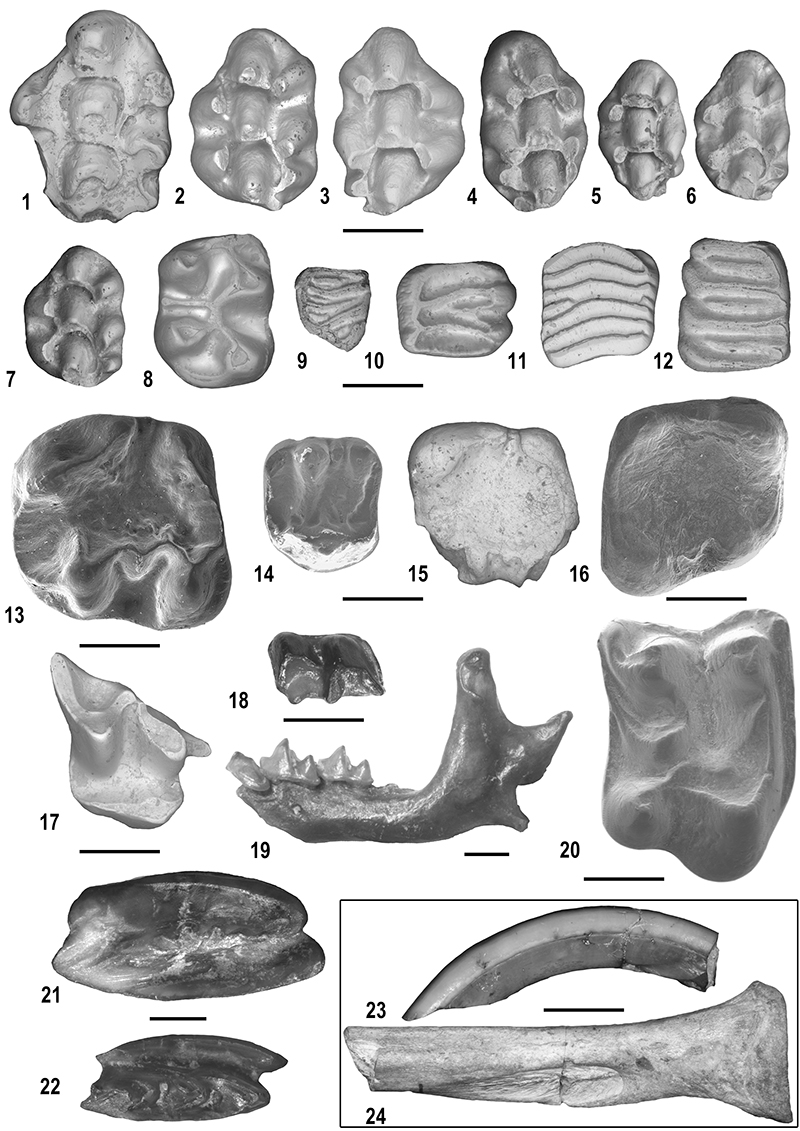
FIGURE 14. Relative abundances of the families of small mammals (chiropeterans excluded) from Moncucco Torinese. Proportions are based on nNISP (normalized number of identified specimens, see Material and methods for further details). Castoridae and Hystricidae (not visible in the graph) are present only in samples MCC3 and MCC4 respectively with only a single specimen each and are not shown herein.
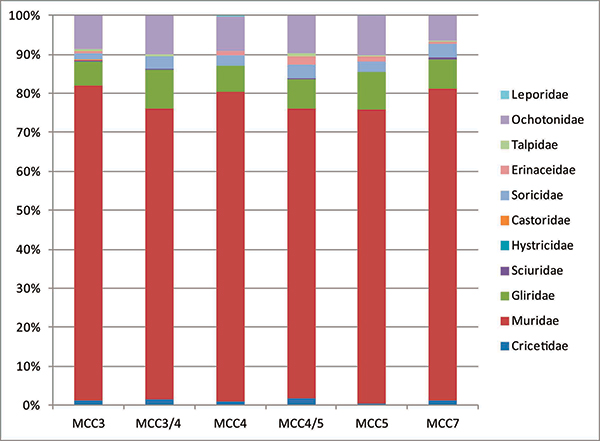
FIGURE 15. Relative abundances of the glirid species within each glirid assemblage from Moncucco Torinese.
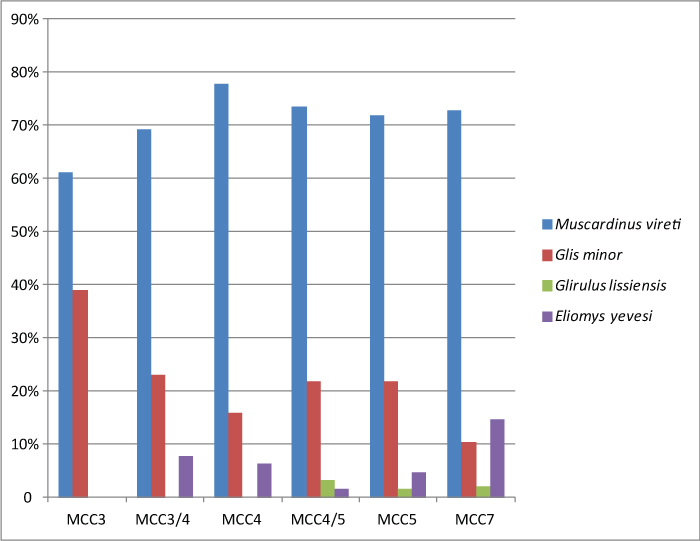
FIGURE 16. Relative abundances of the murid taxa within each murid assemblage of Moncucco Torinese.
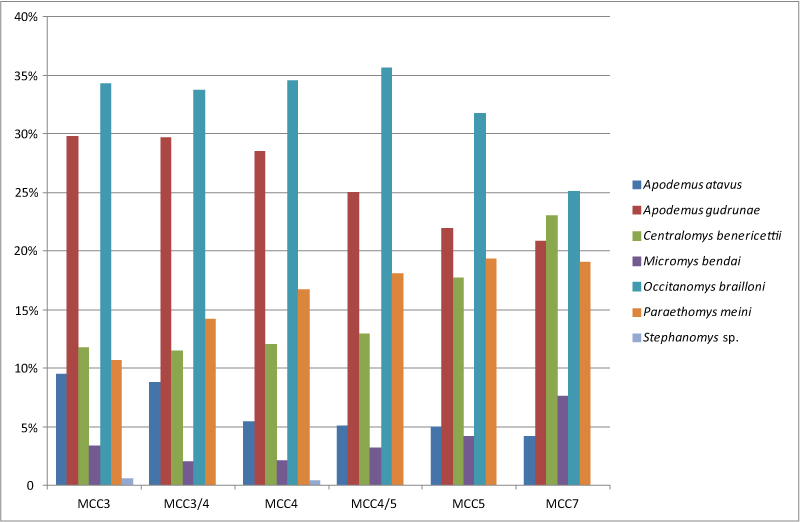
FIGURE 17. Variation of coefficients of the predictor variables of the Binary Logistic Regressions that model the probability of finding a specific murid taxon at MCC assuming the different layers as the only predictor variables. Blue dots indicate the values of the coefficients, bars indicate the 95% CI. MCC3 is chosen as reference category. For each of the identified taxa, the coefficients that are lower (or higher) than 0 indicate how much, respect to MCC3, that variable influences the probability (in terms of log odds) of finding that species at MCC. In other words, lower (or higher) coefficients of each variable (i.e., the given fossil assemblage) indicate that for that assemblage there are lower (or higher) probabilities of finding that species respect to MCC3. Significant statistical differences are obtained when the CI does not include the value 0. On the contrary, values approaching zero indicate that in that assemblage there are the same probabilities of finding that species that we have for MCC3. Stephanomys sp. is not included due to its rarity and not signifacant values. Additional information available in Appendix 8.
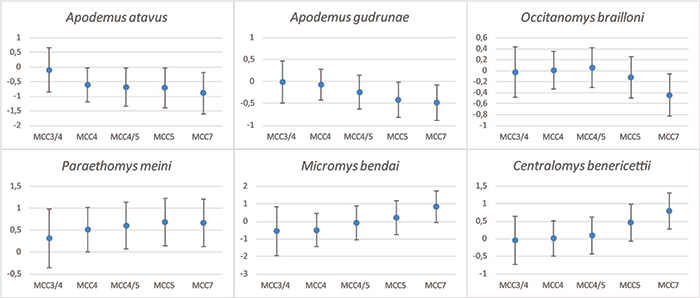
FIGURE 18. Distribution of habitats as indicated by the ecological preferences of small mammals (chiropterans excluded) from each assemblage of Moncucco Torinese.
Late Messinian mollusks and vertebrates from Moncucco Torinese, north-western Italy. Paleoecological and paleoclimatological implications
Simone Colombero, David M. Alba, Carmine D'Amico, Massimo Delfino, Daniela Esu, Piero Giuntelli, Mathias Harzhauser, Paul P.A. Mazza, Michele Mosca, Thomas A. Neubauer, Giulio Pavia, Marco Pavia, Andrea Villa, and Giorgio Carnevale
Plain Language Abstract
A new paleontological site has been recently discovered in the nearby town of Moncucco Torinese, NW Italy. The age of the fossiliferous deposits of this site is constrained between 5.40 and 5.33 Ma (late Miocene, late Messinian). We collected more than 20,000 fossils belonging to 90 vertebrates and 65 mollusk taxa. We study this highly diverse fossil assemblage to get information about the environmental, ecological and climatic conditions of the studied area at the end of the Miocene. The high number of taxa identified at Moncucco Torinese suggests the presence of a variety of continental habitats. Our analyses indicate that the deposition occurred in lagoonal or littoral areas. At that time, the terrestrial landscape was dominated by open woodlands and/or bushlands with scarce closed areas mainly developed around limited and/or ephemeral freshawater basins. Open habitats were also present. The estimated average precipitations and temperatures are very similar to those previously estimated based on paleobotanical data reported for the late Miocene of NW Italy. Overall, these point to moderate moist conditions in a subtropical climate similar to that currently observed in several coastal areas of the northern and eastern Mediterranean.
Resumen en Español
Moluscos y vertebrados del Messiniense tardío de Moncucco Torinese, noroeste de Italia. Implicaciones paleoecológicas y paleoclimáticas
El análisis sistemático de más de 20.000 fósiles (Vertebrata y Mollusca), recuperados de la sucesión post-evaporítica del Messiniense (5,41-5,33 millones de años) de Moncucco Torinese (noroeste de Italia), resultó en la identificación de 90 taxones de vertebrados y 65 taxones de moluscos que proporcionan información sobre el contexto paleoecológico y los paleoambientes del noroeste de Italia un poco antes del límite Mio-Plioceno. Nuestros análisis indican un paisaje dominado por bosques abiertos dentro de un ambiente de mosaico que también incluía bosques de dosel cerrados, pastizales, áreas rocosas y limitados bordes de agua. El amplio espectro de hábitats puede haber tenido un papel prominente en la alta paleobiodiversidad que ha sido constatada en la paleocomunidad de Moncucco Torinese. Las ligeras variaciones en la abundancia de las especies de roedores más comunes a lo largo de la sucesión investigada probablemente están relacionadas con los cambios locales en el paleopaisaje. Desde un punto de vista paleoclimático, la información general proporcionada por la fauna indica condiciones mésicas en un clima subtropical, lo que también es coherente con la interpretación derivada de los análisis paleobotánicos y sedimentológicos del Messiniense tardío del norte de Italia.
Palabras clave: Paleoecología; Mioceno; Messiniense; Turoliense; Italia; Vertebrata; Molusco
Traducción: Enrique Peñalver (Sociedad Española de Paleontología)
Résumé en Français
Implications paléoécologiques et paléoclimatologiques des mollusques et vertébrés du Messinien récent de Moncucco Torinese, nord-ouest de l'Italie
L'analyse taxinomique de plus de 20000 fossiles (Vertebrata et Mollusca), trouvés dans la succession post-évaporitique messinienne (5,41 Ma–5,33 Ma) de Moncucco Torinese (nord-ouest de l'Italie), a donné lieu à l'identification de 90 taxons de vertébrés et 65 taxons de mollusques qui fournissent des informations nouvelles sur le contexte paléoécologique et paléoenvironnemental du nord-ouest de l'Italie un peu avant la limite Miocène-Pliocène. Nos analyses indiquent un paysage dominé par des forêts ouvertes au sein d'un environnement mosaïque incluant aussi des forêts à canopée fermée, des prairies herbeuses, des affleurements rocheux, et des accès limités à des corps d'eau. Le large éventail d'habitats a pu avoir un rôle déterminant sur la paléobiodiversité élevée observée dans la paléocommunauté de Moncucco Torinese. De légères variations d'abondances des espèces de rongeurs les plus communes au sein de la succession étudiée sont probablement liées à des changements locaux dans le paléopaysage. D'un point de vie paléoclimatique, les informations globales fournies par la faune indiquent des conditions mésiques sous un climat subtropical, ce qui est également cohérent avec l'interprétation des analyses paléobotaniques et sédimentologiques du Messinien terminal du nord de l'Italie.
Mots-clés : paléoécologie ; Miocène ; Messinien ; Turolien ; Italie ; Vertebrata ; Mollusca
Translator: Antoine Souron
Deutsche Zusammenfassung
Mollusken und Vertebraten aus dem späten Messinium aus Moncucco Torinese, Nordwest-Italien: paläoökologische und paläoklimatische Rückschlüsse
Die systematische Analyse von mehr als 20.000 Fossilen (Vertebrata und Mollusca) aus den post-evaporitischen Abfolgen des Messiniums (5.41–5.33 Ma) von Moncucco Torinese (NW Italien) ergab 90 Vertebraten-Taxa und 65 Mollusken-Taxa. Sie liefern zusätzliche Information über den paläoökologischen Kontext und die Paläoumgebung von Nordwest-Italien kurz vor der Mio-Pliozän Grenze. Unsere Analyse weist auf eine Landschaft hin, die durch offene Wälder dominiert wird, innerhalb einer Mosaik-Umgebung mit dichten Wäldern, Wiesen, Felsen und limitierten Wasserzugängen. Das große Spektrum an Habitaten mag eine große Rolle für die große Paläodiversität der Paläogemeinschaft von Moncucco Torinese gespielt haben. Leichte Häufigkeitsvariationen bei der am weitesten verbreiteten Nagetier-Art in der untersuchten Abfolge hängen vermutlich mit lokalen Änderungen der Paläolandschaft zusammen. Paläoklimatisch gesehen weist die allgemeine, durch die Fauna gewonnene Information, auf mesische Konditionen in einem subtropischen Klima hin. Dies steht auch im Einklang mit Interpretationen aus paläobotanischen und sedimentologischen Analysen aus dem späten Messinium von Norditalien.
Schlüsselwörter: Paläoökologie; Miozän; Messinium; Turolium; Italien; Vertebrata; Mollusca
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024