

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
APPENDIX 1.
Generalized baboon morphology illustrating measurements taken in this study.
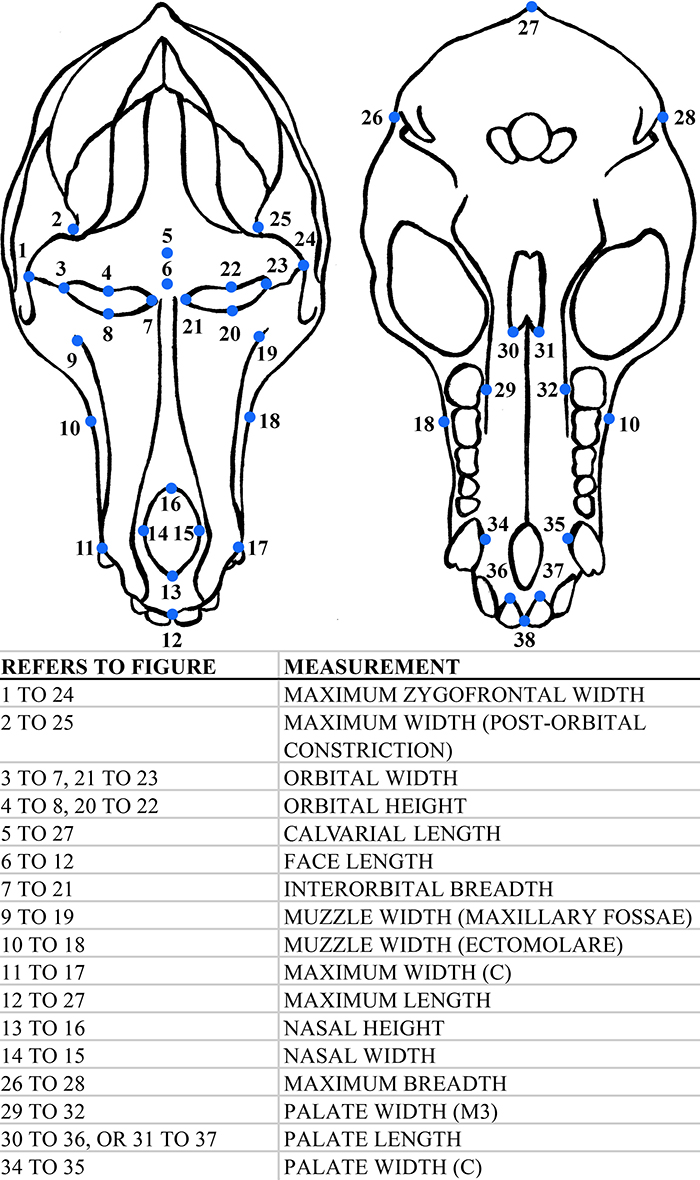
APPENDIX 2.
Raw data for fossils used in this study. A single set of parentheses indicates that the measurement was estimated with a high degree of confidence, and the values were used in the analyses. A double set of parentheses indicates that the measurement was estimated with a low degree of confidence, and the values were not used in the analyses. (Appendixes 2-8 available as spreadsheets in zipped file.)
APPENDIX 3.
Raw data for the extant papionins used in this study. (Appendixes 2-8 available as spreadsheets in zipped file.)
APPENDIX 4.
Descriptive statistics for the cranial measurements taken in this study, by fossil species. (Appendixes 2-8 available as spreadsheets in zipped file.)
APPENDIX 5.
Full ANOVA detailing genus comparisons for raw cranial measurements across fossil genera. Shaded cells are significant at p < 0.05. (Appendixes 2-8 available as spreadsheets in zipped file.)
APPENDIX 6.
ANOVA detailing extant genus comparisons for geometric mean scaled data for a subset of cranial measurements. Shaded cells are significant at p < 0.05, n = 81. (Appendixes 2-8 available as spreadsheets in zipped file.)
APPENDIX 7.
ANOVA detailing fossil genus comparisons for geometric mean scaled data for a subset of cranial measurements. Shaded cells are significant at p < 0.05, n = 25. (Appendixes 2-8 available as spreadsheets in zipped file.)
APPENDIX 8.
Full correlation matrix detailing fossil correlations and sample size for the 17 geometric mean scaled cranial measurements. Yellow highlighted cells are significant at p < 0.0004, and grey shaded cells give sample sizes for each measurement pair. (Appendixes 2-8 available as spreadsheets in zipped file.)
TABLE 1. Genera and species represented, repository information, sample sizes, and specimen numbers used in this study. U Wits: University of the Witwatersrand, Johannesburg, South Africa; UCMP: University of California Museum of Paleontology, Berkeley, California, USA; Ditsong: Ditsong National Museum of Natural History, Pretoria, South Africa. TM211 was originally described as Papio spelaeus (Broom, 1936). M3147 has also been published as P. h. robinsoni (Gilbert et al., 2015).
| Genus | Species | Repository | N | Specimen numbers | Site(s) | References |
| Cercopithecoides | C. molletti | Ditsong | 1 | SK551 | Swartkrans | McKee et al. (2011) |
| C. williamsi | Ditsong | 3 | KA195, KB5241C, MP36 | Kromdraii, Makapansgat | Szalay and Delson (1979), McKee et al. (2011), Geissler (2013) | |
| U Wits | 1 | STS394A | Sterkfontein | Geissler (2013) | ||
| Total | 5 | |||||
| Dinopithecus | D. ingens | Ditsong | 5 | SK553, SK554, SK599, SK600, SK604 | Swartkrans | Gilbert (2013) |
| Total | 5 | |||||
| Gorgopithecus | G. major | Ditsong | 3 | KA153, KA605, KA944 | Kromdraii | Gilbert (2013) |
| Total | 3 | |||||
| Papio | P. angusticeps | Ditsong | 5 | CO100, CO101, CO102, CO135A, KA194 | Cooper's Cave, Kromdraii | McKee and Keyser (1994), Williams et al. (2007), Gilbert (2013) |
| P. h. robinsoni | Ditsong | 5 | SK555, SK557, SK558, SK560, SK562 | Swartkrans | Carter (2006), Geissler (2013), Beaudet et al. (2016) | |
| UCMP | 2 | UCMP 56767, UCMP 56797 | Bolt's Farm | Freedman (1965) | ||
| U Wits | 1 | SWP31 | Sterkfontein | Eisenhart (1974) | ||
| P. h. ursinus | Ditsong | 1 | TM211 | Freedman (1957) | ||
| P. izodi | U Wits | 5 | T89-11-1, TP4, TP7, TP10, TP12 | Taung | Gilbert and Grine (2010), Gilbert (2013) | |
| Total | 19 | |||||
| Parapapio | P. broomi | Ditsong | 6 | STS254A, STS262, STS393, STS396A, STS397, STS564 | Sterkfontein | Carter (2006), Williams et al. (2007), Gilbert (2013) |
| UCMP | 1 | UCMP 56786 | Bolt's Farm | Freedman (1965) | ||
| U Wits | 7 | M3056, M3065, MP151, MP2, MP224, MP76, SWP12 | Makapansgat, Sterkfontein | Fourie (2006), Fourie et al. (2008), Gilbert (2013) | ||
| P. jonesi | Ditsong | 2 | STS260, STS565 | Sterkfontein | Pickering et al. (2004), Fourie et al. (2008), Gilbert (2013) | |
| U Wits | 5 | M3060, MP75, MP173, SWP389, SWP2947 | Makapansgat, Sterkfontein | Maier (1970) | ||
| P. whitei | UCMP | 1 | UCMP 56614 | Buxton Limeworks | Freedman (1965) | |
| U Wits | 10 | BF43, M3070, M3072, M3147, MP119, MP164, MP208, MP221, MP223, MP239 | Bolt's Farm, Makapansgat | Maier (1977), Williams et al. (2007), Fourie et al. (2008), Williams (2013) | ||
| unknown | U Wits | 4 | M3053, M3084, M3133, MP47 | Makapansgat | Fourie (2006), Fourie et al. (2008) | |
| Total | 36 | |||||
| Procercocebus | P. antiquus | UCMP | 3 | UCMP 56609, UCMP 56694, UCMP 125956 | Buxton Limeworks | Gilbert (2007) |
| U Wits | 5 | M3078, M3079, T89-154, TP8, TP9 | Makapansgat, Taung | Gilbert (2013) | ||
| Total | 8 | |||||
| Theropithecus | T. darti | U Wits | 4 | M3073, MP168, MP217, MP222 | Makapansgat | Maier (1972), Williams et al. (2007), Geissler (2013), Gilbert (2013) |
| T. oswaldi | Ditsong | 2 | SK561, SK563 | Swartkrans | Folinsbee and Reisz (2013) | |
| Total | 6 | |||||
| unknown | unknown | Ditsong | 6 | SB2, SK3, SK556, SK814, SKII25, STS537 | Swartkrans, Sterkfontein | N/A |
| UCMP | 1 | UCMP 218738 | Buxton Limeworks | Monson et al. (2015) | ||
| U Wits | 10 | unnumbered black skull, MP4A, SP1, SP24, SWP1728, SWP1731, SWP29, SWP2953, T8817, T9087 | Makapansgat, Sterkfontein, Taung | N/A | ||
| Total | 17 | |||||
| TOTAL | 99 |
TABLE 2. Extant papionin species represented and sample sizes used in this study. All specimens are from the National Museum of Natural History in Washington D.C., USA, with the exception of one Theropithecus specimen from the National Museum in Addis Ababa, Ethiopia.
| Species | Sample size (n) |
| Cercocebus torquatus | 20 |
| Lophocebus albigena | 15 |
| Mandrillus sphinx | 7 |
| Papio hamadryas | 34 |
| Theropithecus gelada | 5 |
| Total | 81 |
TABLE 3. Definitions of cranial measurements.
| Measurement | Definition |
| Maximum length | Maximum length of the cranium from prosthion to inion. |
| Maximum breadth | Maximum breadth of the cranium posterior to the external auditory meatus. |
| Orbital width | Maximum width of the orbit along a line perpendicular to the orbital height. |
| Orbital height | Maximum height of the orbit along a parasagittal plane. |
| Interorbital breadth | Breadth between the superomedial margins of the orbits. |
| Nasal width | Maximum width of the nasal aperture, from left to right alare. |
| Nasal height | Maximum height of the nasal aperture, from nasospinale to rhinion. |
| Muzzle width (ectomolare) | Maximum width of the muzzle, measured at the ectomolare. |
| Muzzle width (maxillary fossae) | Maximum width of the muzzle, measured at the superior border of the maxillary fossae, inferior to the orbits. |
| Maximum width (canine) | Maximum width of the muzzle, measured at the canine alveoli. |
| Maximum zygofrontal width | Maximum width of the upper face, measured from left to right frontomalare temporale. |
| Maximum calvarial width (post-orbital constriction) | Maximum calvarial width, measured at the post-orbital constriction. |
| Calvarial length | Maximum calvarial length, measured from ophryon to inion. |
| Palate width (M3) | Maximum width of the palate, measured from the lateral edges of the depth of the palate, along a line bisecting the third molars. |
| Palate width (canine) | Maximum width of the palate, measured from the lateral edges of the depth of the palate, along a line bisecting the most medial points of the canine alveoli. |
| Palate length | Maximum length of the palate, measured from orale to staphylion. |
| Face length | Projection of the face, measured from prosthion to glabella. |
TABLE 4. Sample size, mean, and standard deviation of each measurement for the fossil genera sampled in this study. All measurements are in millimeters.
| Genus |
Cercopithecoides |
Dinopithecus |
Gorgopithecus |
Papio |
Parapapio |
Procercocebus |
Theropithecus |
Unknown |
|||||||||
| Sample size | 5 | 5 | 3 | 19 | 36 | 8 | 6 | 17 | |||||||||
| Statistic | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Maximum length | 124.9 | NA | 173.9 | 21.7 | NA | NA | 148.2 | 9.8 | 152.4 | 19.9 | 147.9 | 16.5 | 171.5 | 8.1 | 163.9 | 30.6 | |
| Maximum breadth | 81.6 | 6.2 | 87.1 | 5.9 | NA | NA | 71.6 | 2.0 | 76.2 | 5.5 | 70.9 | 7.2 | 84.1 | 5.3 | 77.6 | 6.6 | |
| Orbital width | 29.4 | 2.2 | 30.1 | 6.0 | NA | NA | 27.5 | 1.9 | 26.9 | 2.2 | 26.4 | 1.6 | 28.7 | 2.4 | 27.0 | 2.3 | |
| Orbital height | 27.9 | 0.8 | 30.1 | 6.0 | NA | NA | 24.3 | 3.0 | 24.5 | 2.4 | 23.5 | 1.5 | 26.9 | 4.9 | 25.3 | 3.5 | |
| Interorbital | 15.0 | 2.9 | 11.9 | 5.9 | NA | NA | 11.5 | 2.7 | 11.4 | 2.0 | 8.9 | 1.2 | 12.1 | 1.5 | 11.8 | 2.4 | |
| Nasal width | 14.9 | 2.5 | 16.1 | 3.5 | 16.4 | NA | 19.1 | 4.0 | 17.6 | 2.4 | 17.7 | 1.8 | 18.9 | 1.7 | 18.0 | 2.0 | |
| Nasal height | 18.8 | NA | 27.6 | 1.8 | NA | NA | 29.8 | 9.1 | 26.5 | 4.2 | 25.8 | 2.7 | 29.3 | 2.4 | 29.3 | 8.0 | |
| Muzzle width (ectomolare) |
44.6 | 2.2 | 58.6 | 10.2 | 58.2 | NA | 51.4 | 8.0 | 51.2 | 4.2 | 46.8 | 2.4 | 51.9 | 3.1 | 48.1 | 6.6 | |
| Muzzle width (maxillary fossae) |
25.5 | 3.9 | 39.9 | 5.8 | NA | NA | 40.0 | 8.9 | 37.1 | 4.4 | 36.4 | 4.2 | 35.5 | 4.5 | 39.4 | 5.4 | |
| Maximum width (C) | 30.6 | 2.7 | 38.7 | 9.4 | 43.0 | 3.4 | 41.4 | 11.3 | 36.6 | 5.8 | 35.2 | 4.1 | 33.8 | 2.1 | 38.6 | 4.8 | |
| Maximum zygofrontal width | 81.0 | 3.2 | 88.1 | 21.3 | NA | NA | 72.4 | 6.5 | 72.4 | 6.9 | 75.8 | 4.9 | 77.1 | 1.1 | 80.5 | 5.4 | |
| Maximum cal. width | 54.9 | 3.8 | 54.1 | 0.9 | NA | NA | 53.4 | 5.8 | 53.3 | 3.0 | 56.4 | 3.6 | 48.2 | 4.7 | 53.4 | 3.5 | |
| Calvarial length | 94.4 | 10.7 | 92.3 | NA | NA | NA | 88.5 | 3.5 | 94.3 | 8.3 | 87.9 | 4.7 | 95.4 | 10.5 | 90.8 | 6.6 | |
| Palate width (M3) | 19.2 | 1.5 | 25.1 | 6.9 | 25.9 | 7.1 | 23.9 | 5.2 | 24.1 | 2.3 | 20.5 | 2.7 | 24.3 | 4.7 | 21.2 | 2.9 | |
| Palate width (C) | 18.8 | 1.8 | 27.8 | 8.1 | 23.6 | 0.0 | 24.2 | 6.2 | 21.5 | 3.0 | 17.8 | 2.8 | 20.4 | 1.1 | 22.0 | 3.0 | |
| Palate length | 40.4 | 2.1 | 68.9 | 10.5 | 65.5 | 4.4 | 67.4 | 16.9 | 51.5 | 8.2 | 51.2 | 6.0 | 62.9 | 3.8 | 57.6 | 11.7 | |
| Face length | 62.4 | 6.5 | 98.7 | 29.7 | NA | NA | 99.0 | 30.7 | 82.2 | 11.6 | 76.1 | 10.6 | 93.0 | 0.9 | 94.0 | 28.6 | |
TABLE 5. Results of the MANOVA for the extant and fossil papionins. Double asterisks denote significance at p < 0.001.
| Traits | Group | p |
| Max. length, Max. breadth, Orb. width, Orb. height, Interorb. breadth, Nas. width, Nas. height, Muz. width (ecto.), Muz. width (max. fos.), Max. cal. width | Extant | 0.000** |
| Max. length, Max. breadth, Orb. width, Orb. height, Interorb. breadth, Nas. width, Nas. height, Muz. width (ecto.), Muz. width (max. fos.), Max. cal. width | Fossil | 0.439 |
| Cal. length, Palate width (M3), Palate width (C), Palate length, Face length | Extant | 0.000** |
| Cal. length, Palate width (M3), Palate width (C), Palate length, Face length | Fossil | 0.382 |
TABLE 6. ANOVA results for the size-corrected values of the extant sample, n = 81. MANOVA is significant at p < 2.2e-16. Measurements are size-corrected using geometric mean (Mosimann, 1970). Shaded cells are significant at p < 0.05.
| Genus comparison | Muz. Width (Ecto.) |
Muz. Width (Canine) |
Palate Width (Canine) | Palate Length | Face Length |
| Lophocebus-Cercocebus | 0.087 | 0.017 | 0.017 | 0.980 | 0.036 |
| Mandrillus-Cercocebus | 0.000 | 0.712 | 0.712 | 0.000 | 0.002 |
| Papio-Cercocebus | 0.000 | 0.040 | 0.040 | 0.000 | 0.000 |
| Theropithecus-Cercocebus | 1.000 | 0.224 | 0.224 | 0.000 | 0.000 |
| Mandrillus-Lophocebus | 0.000 | 0.007 | 0.007 | 0.000 | 0.000 |
| Papio-Lophocebus | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| Theropithecus-Lophocebus | 0.522 | 0.001 | 0.001 | 0.000 | 0.000 |
| Papio-Mandrillus | 0.192 | 0.987 | 0.987 | 0.991 | 0.024 |
| Theropithecus-Mandrillus | 0.000 | 0.938 | 0.938 | 0.007 | 0.438 |
| Theropithecus-Papio | 0.000 | 0.986 | 0.986 | 0.000 | 0.957 |
TABLE 7. ANOVA results for a subset of the size-corrected values of the fossil sample, n = 38. MANOVA is significant at p < 0.0009. Measurements are size-corrected using geometric mean (Mosimann, 1970). Shaded cells are significant at p < 0.05.
| Genus Comparison | Muz. Width (Ecto.) |
Muz. Width (Canine) |
Palate Width (Canine) | Palate Length | Face Length |
| Dinopithecus-Cercopithecoides | 0.404 | 0.687 | 0.991 | 0.140 | 0.355 |
| Papio-Cercopithecoides | 0.035 | 0.966 | 0.930 | 0.004 | 0.300 |
| Parapapio-Cercopithecoides | 0.876 | 0.998 | 1.000 | 0.980 | 0.853 |
| Procercocebus-Cercopithecoides | 0.855 | 0.999 | 0.504 | 0.301 | 0.771 |
| Theropithecus-Cercopithecoides | 0.733 | 0.152 | 0.647 | 0.011 | 0.088 |
| Papio-Dinopithecus | 0.975 | 0.895 | 1.000 | 0.966 | 0.998 |
| Parapapio-Dinopithecus | 0.682 | 0.651 | 0.981 | 0.103 | 0.642 |
| Procercocebus-Dinopithecus | 0.855 | 0.318 | 0.892 | 0.938 | 0.882 |
| Theropithecus-Dinopithecus | 0.958 | 0.963 | 0.954 | 0.982 | 0.998 |
| Parapapio-Papio | 0.005 | 0.979 | 0.607 | 0.000 | 0.421 |
| Procercocebus-Papio | 0.099 | 0.605 | 0.800 | 0.167 | 0.912 |
| Theropithecus-Papio | 0.292 | 0.150 | 0.932 | 1.000 | 0.842 |
| Procercocebus-Parapapio | 1.000 | 0.837 | 0.090 | 0.169 | 0.997 |
| Theropithecus-Parapapio | 0.983 | 0.025 | 0.236 | 0.001 | 0.089 |
| Theropithecus-Procercocebus | 0.999 | 0.011 | 1.000 | 0.342 | 0.400 |
TABLE 8. Sample size and correlations for a subset of size-corrected measurement comparisons. A full correlation matrix comparing all size-corrected measurements is available in Appendix 8. All correlations are significant at p < 0.0004. “(C)” is the abbreviation for canine.
| Measurement pair | Sample size (n) |
Correlation (r) |
| Maximum length - face length | 31 | 0.898 |
| Maximum length - palate length | 35 | 0.880 |
| Maximum zygofrontal width - orbital width | 32 | 0.835 |
| Palate length - face length | 45 | 0.827 |
| Maximum length - maximum breadth | 29 | 0.817 |
| Maximum length - calvarial length | 31 | 0.814 |
| Maximum width (C) - palate width (C) | 54 | 0.798 |
| Calvarial length - face length | 29 | 0.784 |
| Calvarial length - maximum breadth | 39 | 0.770 |
| Muzzle width ectomolare - palate width (C) | 54 | 0.764 |
| Face length - nasal height | 35 | 0.760 |
| Maximum width (C) - face length | 43 | 0.758 |
FIGURE 1. Six of the fossil cercopithecid specimens used in this study. 1, UCMP 125854, Papio izodi. 2, UCMP 125856, Papio izodi. 3, MP221, Parapapio jonesi. 4, TP9, Procercocebus antiquus. 5, MP222, Theropithecus darti. 6, STS394A, Cercopithecoides williamsi.
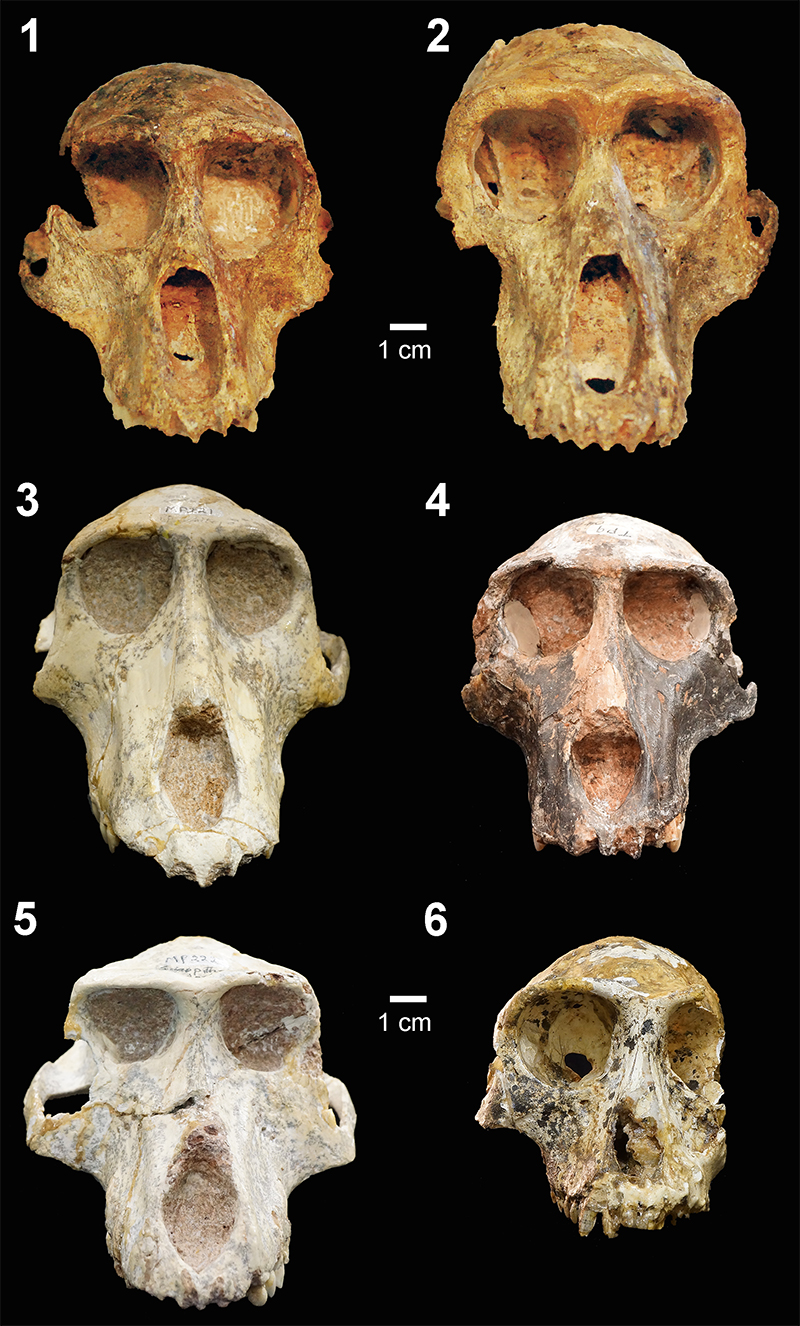
FIGURE 2. Bivariate plots of cranial measurements of extant papionins and the fossil sample. 1 and 2, Bivariate plot comparing cranial length and cranial breadth in extant papionins (1) and the fossil sample (2). 3 and 4, Bivariate plot comparing face length and muzzle width (ectomolare) in extant papionins (3) and the fossil sample (4). Note high correlations and the lack of a consistent taxonomic pattern across the distribution of fossil specimens. All bivariate correlations are significant at p < 0.0004.
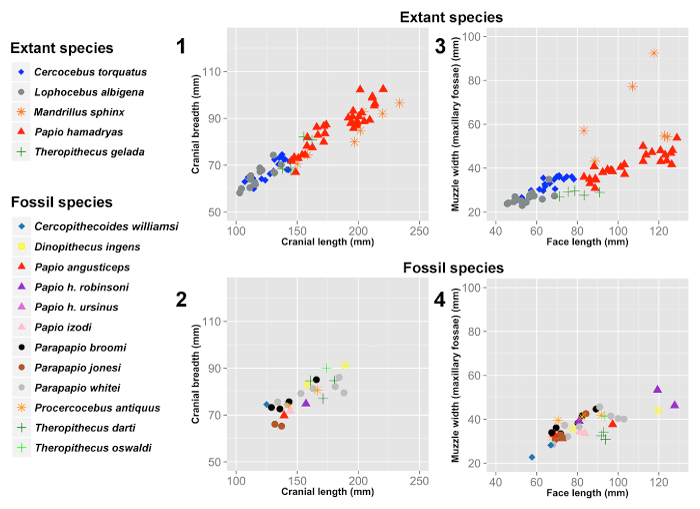
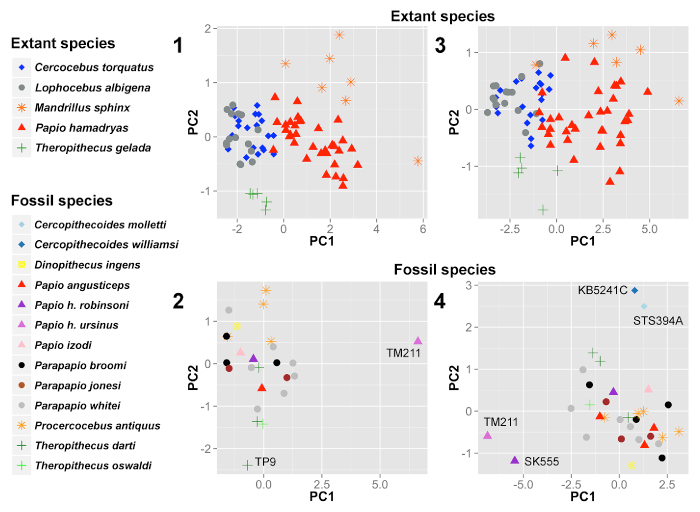
FIGURE 3. Principle components analyses (PCA) of cranial variation in extant papionins and the fossil sample. 1 and 2, PCA including maximum cranial length, orbital width, interorbital breadth, and palate length raw measurements for 81 extant papionins (1) and 25 fossil cercopithecids (2). PC1 and PC2 of the extant sample comprise 82.2% and 8.1% of the variation, respectively, and PC1 and PC2 of the fossil sample comprise 67.8% and 21.6% of the variation, respectively. Note how well the measurements discriminate extant species, and how Papio h. ursinus (TM211) and one specimen of Theropithecus darti (TP9) are distinct from the other fossils. 3 and 4, PCA including orbital width, interorbital breadth, muzzle width (ectomolare), maximum width (canine), palate width (M3), palate width (canine), and palate length in 81 extant papionins (3) and 33 fossil cercopithecids (4). PC1 and PC2 of the extant sample comprise 86.6% and 4.8% of the variation, respectively, and PC1 and PC2 of the fossil sample comprise 66.1% and 12.7% of the variation, respectively. Note how well the measurements discriminate extant species, and that Cercopithecoides and Papio hamadryas are distinct from the other fossils.
 Tesla A. Monson. Department of Integrative Biology, University of California, Berkeley, 3040 Valley Life Sciences Building #3140, Berkeley, CA 94720, USA. tesla.monson@berkeley.edu
Tesla A. Monson. Department of Integrative Biology, University of California, Berkeley, 3040 Valley Life Sciences Building #3140, Berkeley, CA 94720, USA. tesla.monson@berkeley.edu
Tesla Monson is a Ph.D. candidate in the Department of Integrative Biology at the University of California, Berkeley. Her research focuses on the evolution of mammalian craniodental variation, the evolution of body size and life history traits in mammals, and the interpretation of mammalian species variation in the fossil record. Tesla's dissertation addresses the relationship between phylogeny, life history, and dental eruption sequence in mammals. Tesla also hosts and produces The Graduates on KALX 90.7FM, a talk radio show dedicated to science education and outreach, and diversity in STEM.

 Marianne F. Brasil. Department of Integrative Biology, University of California, Berkeley; Human Evolution Research Center, 3040 Valley Life Sciences Building #3140, Berkeley, CA 94720, USA. mbrasil@berkeley.edu
Marianne F. Brasil. Department of Integrative Biology, University of California, Berkeley; Human Evolution Research Center, 3040 Valley Life Sciences Building #3140, Berkeley, CA 94720, USA. mbrasil@berkeley.edu
Marianne Brasil is a Ph.D. candidate in the Department of Integrative Biology at the University of California, Berkeley. Marianne's research interest is in mammalian dental and skeletal variation, particularly how variation is patterned and interpreted in the fossil record. Marianne's dissertation research focuses on early modern human skeletal remains from the Middle Awash area of Ethiopia, and she is an active member of the Middle Awash research project.
 Dominic J. Stratford. School of Geography, Archaeology and Environmental Studies, University of the Witwatersrand, 1 Jan Smuts Avenue, Braamfontein 2000, Johannesburg, South Africa. Dominic.Stratford@wits.ac.za
Dominic J. Stratford. School of Geography, Archaeology and Environmental Studies, University of the Witwatersrand, 1 Jan Smuts Avenue, Braamfontein 2000, Johannesburg, South Africa. Dominic.Stratford@wits.ac.za
Dominic Stratford is research director of the Sterkfontein Caves and senior lecturer of geoarchaeology at the University of the Witwatersrand, Johannesburg, South Africa. Dominic's research interests focus on the development and application of multidisciplinary analyses to help refine contextual control of palaeoanthropological and archaeological assemblages during and after their excavation. As a consequence of this focus, Dominic's interests cover a range of earth and biological sciences from vertebrate morphology to sediment micromorphology. Dominic is involved in field work in several areas of South Africa and Namibia spanning the late Pliocene to Holocene.
 Leslea J. Hlusko. Department of Integrative Biology, University of California, Berkeley; Human Evolution Research Center, 3040 Valley Life Sciences Building #3140, Berkeley, CA 94720, USA. hlusko@berkeley.edu
Leslea J. Hlusko. Department of Integrative Biology, University of California, Berkeley; Human Evolution Research Center, 3040 Valley Life Sciences Building #3140, Berkeley, CA 94720, USA. hlusko@berkeley.edu
Leslea Hlusko's research explores the genetic underpinnings of skeletal variation and how this has evolved through time as seen in the fossil record. The focus is on mammalian evolution, with an emphasis on primates. Lab methods span from quantitative genetic analyses to gene expression studies. Leslea has co-directed field paleontology projects in Kenya and Tanzania, and participates in field projects in Ethiopia and Turkey. Her paleontological research is focused on mammalian evolution in the Neogene of Africa and Eurasia. She is currently an Associate Professor of Integrative Biology at the University of California Berkeley.
Patterns of craniofacial variation and taxonomic diversity in the South African Cercopithecidae fossil record
Plain Language Abstract
South Africa has played a central role in our understanding of human evolution. However, the difficulty associated with dating fossils recovered from South African caves has placed a large burden on the accurate interpretation of species associated with hominids, which are used as indicators of the age of deposits, or 'biochronological markers'. Old World Monkeys (OWMs) are frequent biochronological markers at these sites, with dated fossils from eastern Africa being used to date South African sites where the same species are present. The interpretation of fossil OWM taxa relies heavily on morphology, particularly cranial and dental remains. Many of the analytical methods used to interpret the taxonomy of this group also rely on the assumption that the cranial and dental traits are developmentally, functionally, and genetically independent, an assumption that has been increasingly questioned. We statistically explored cranial variation in South African fossil OWMs (n = 99) to determine whether or not extinct taxa reflect the same morphological patterns of variation as has been reported for extant groups. Our results show that many of the cranial measurements are statistically significantly correlated and fail to distinguish between species or even genera of fossil OWMs despite the fact that these measurements do distinguish between extant species. Overall, our results suggest that these extinct OWM taxa cannot be discriminated using cranial measurements alone, or that the taxonomic designations, as they currently stand, are confounded.
Glossary:
OWM – Old World Monkey
Modules – interrelated suites of traits
Papionins – members of Tribe Papionini, Family Cercopithecidae
Cercopithecids – members of Family Cercopithecidae
Resumen en Español
Patrones de variación craneofacial y diversidad taxonómica en el registro fósil de Cercopithecidae de Sudáfrica
El rico registro paleontológico de Sudáfrica es capital en nuestra comprensión de la evolución de los mamíferos del Plio-Pleistoceno debido en gran parte al número de cráneos recuperados. Debido a la dificultad del control cronométrico, en muchos de los sistemas de cuevas de los cuales proceden estos fósiles, a menudo se emplean los Monos del Viejo Mundo extintos (OWMs) como marcadores biocronológicos, por lo que la identificación taxonómica de estos restos de primates es particularmente relevante para hacerse preguntas evolutivas más amplias relacionadas con la evolución humana y los cambios de diversidad faunística. Las relaciones taxonómicas de los fósiles OWM se reconstruyen a través de la sistemática filogenética que depende en gran medida de los caracteres craneodentales. Estos métodos analíticos suponen que estos caracteres son independientes respecto al desarrollo, la funcionalidad y la genética. Esta suposición es cada vez más cuestionada por los análisis de datos fenotípicos existentes y por la genética. Se estudió estadísticamente la variación craneal en los Papionini fósiles sudafricanos (n = 99) para determinar si los taxones extintos reflejan la misma estructura de covarianza fenotípica que se ha establecido para los OWM existentes. Nuestros resultados muestran que muchas de las mediciones craneales están correlacionadas estadísticamente y no distinguen entre especies o incluso géneros de Papionini fósiles a pesar de distinguir especies existentes. En general, nuestros resultados sugieren que estos taxones extintos OWM no pueden ser discriminados utilizando mediciones craneofaciales únicamente, o que las designaciones taxonómicas, tal como están establecidas actualmente, son confusas.
Palabras clave: Monos del Viejo Mundo; Pleistoceno; Taxonomía; Papionini; morfología; covariación fenotípica
Traducción: Enrique Peñalver (Sociedad Española de Paleontología)
Résumé en Français
Schémas de variation crânio-faciale et diversité taxinomique dans le registre fossile sud-africain des Cercopithecidae
Le riche registre paléontologique d'Afrique du Sud est central pour la compréhension de l'évolution des mammifères au Plio-Pléistocène en grande partie grâce au grand nombre de crânes trouvés. Afin de pallier les difficultés de contrôle chronologique des nombreux systèmes de grottes d'où proviennent ces fossiles, les Singes de l'Ancien Monde (« Old World Monkeys », OWM) sont fréquemment utilisés comme marqueurs biochronologiques, ce qui rend l'identification taxinomique de ces restes de primates particulièrement pertinente pour explorer des questions évolutives plus larges liées à l'évolution humaine et aux changements de diversité des faunes. Les relations taxinomiques des fossiles d'OWM sont reconstruites par la systématique phylogénétique qui s'appuie fortement sur les traits crânio-dentaires. Ces méthodes analytiques supposent que ces caractères sont indépendants en termes de développement, fonction, et génétique. Cette hypothèse est de plus en plus remise en cause par des analyses de jeux de données sur le phénotype et le génotype des espèces actuelles. Nous avons exploré statistiquement la variation crânienne des Papionini sud-africains fossiles (n = 99) pour déterminer si les taxons éteints présentaient la même structure de covariance phénotypique que celle des OWM actuels. Nos résultats montrent que de nombreuses mesures crâniennes sont significativement corrélées et ne permettent pas de distinguer les espèces et même les genres de Papionini fossiles alors que ces mêmes mesures distinguent les espèces actuelles. En général, nos résultats suggèrent que ces taxons éteints d'OWM ne peuvent pas être discriminés en n'utilisant que des mesures crânio-faciales, et que leurs appellations taxinomiques actuelles prêtent à confusion.
Mots-clés : Singes de l'Ancien Monde ; Pléistocène ; taxinomie ; Papionini ; morphologie ; covariation phénotypique
Translator: Antoine Souron
Deutsche Zusammenfassung
Kraniofaziale Variations-Modelle und taxonomische Diversität im südafrikanischen Fossilbericht der Cercopithecidae
Der reichhaltige paläontologische Nachweis von Südafrika ist ein wesentlicher Bestandteil unseres Verständnisses der plio-pleistozänenen Säugetierevolution, der zum großen Teil der Anzahl der Schädelfunde zu verdanken ist. Wegen der schwierigen chronometrischen Kontrolle vieler Höhlensysteme aus denen die Fossilien stammen, werden oft ausgestorbene Altweltaffen als biochronologische Marker benutzt, was die taxonomische Zuordnung dieser Primatenüberreste besonders wichtig bei der Frage nach der Evolution des Menschen und Diversitätsveränderungen macht.
Die taxonomischen Beziehungen der Altweltaffen wurden mit phylogenetischer Systematik rekonstruiert, die stark auf craniodentalen Merkmalen beruht. Diese analytischen Methoden setzen voraus, dass diese Merkmale entwicklungstechnisch, funktional und genetisch unabhängig sind. Diese Annahme wird jedoch zunehmend durch Analysen von heutigen phänotypischen Datensätzen und der Genetik in Frage gestellt. Wir untersuchten statistisch kraniale Variation bei fossilen südafrikanischen Papioninen (n = 99) um zu bestimmen, ob ausgestorbene Taxa dieselbe phänotypische Kovarianz-Struktur reflektieren wie es für Altweltaffen beschrieben wurde. Unsere Ergebnisse zeigen, dass viele kranialen Maße statistisch signifikant korreliert sind und daher keine Unterscheidungen zwischen Arten oder sogar Gattungen fossiler Papioninen getroffen werden können, trotz der Unterscheidung heutiger Arten. Allgemein legen unsere Ergebnisse nahe, dass diese ausgestorbenen Altweltaffen nicht alleine auf kraniofaziale Messwerte reduziert werden können oder, dass sie nach gegenwärtigem Stand, unstrukturiert sind.
Schlüsselwörter: Altweltaffen; Pleistozän; Taxonomie; Papionini; Morphologie; phänotypische Kovariation
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024