

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
 Andrea Savorelli. Department of Earth Sciences, University of Florence, via La Pira 4, Florence 50121, Italy; andrea_savorelli@yahoo.it
Andrea Savorelli. Department of Earth Sciences, University of Florence, via La Pira 4, Florence 50121, Italy; andrea_savorelli@yahoo.it
Andrea Savorelli, of the Department of Earth Sciences at the University of Florence, is a postdoctoral researcher in vertebrate paleontology with expertise in the study of Neogene micromammals and insular evolution. His current research interests include the systematics, evolution and biogeography of insular mammals, with special focus on the ancestor-descendant relationships among insular taxa.

 Federico Masini. Department of Earth and Marine Sciences (DISTEM), University of Palermo, Via Archirafi 22, Palermo 90123, Italy; federico.masini@unipa.it
Federico Masini. Department of Earth and Marine Sciences (DISTEM), University of Palermo, Via Archirafi 22, Palermo 90123, Italy; federico.masini@unipa.it
Federico Masini works at the Department of Earth and Marine Science of the University of Palermo. He is a paleontologist specialized in the study of Neogene and Quaternary mammals. His interest is focused on taxonomy, paleoclimate, and biogeography. He has a long experience in the study of insular mammals and on evolutionary processes.
 Paul P. A. Mazza. Department of Earth Sciences, University of Florence, via La Pira 4, Florence 50121, Italy; paul.mazza@unifi.it
Paul P. A. Mazza. Department of Earth Sciences, University of Florence, via La Pira 4, Florence 50121, Italy; paul.mazza@unifi.it
Paul Mazza, of the Department of Earth Sciences at the University of Florence, is a paleontologist with expertise in vertebrate paleontology, vertebrate taphonomy, biogeography and paleoclimate. His current research focuses on the paleobiology and taphonomy of continental mammals. Another of his major research addresses are the systematics, evolution and biogeography of insular mammals, with special focus on the patterns and drivers of mammal colonization, diversity and body size on islands.
 Maria Adelaide Rossi. Archaeological Superintendency of Abruzzo, via degli Agostiniani 14, Chieti 66100, Italy; mariaadelaide.rossi@beniculturali.it
Maria Adelaide Rossi. Archaeological Superintendency of Abruzzo, via degli Agostiniani 14, Chieti 66100, Italy; mariaadelaide.rossi@beniculturali.it
Maria Adelaide Rossi works at the Superintendence for archeology, fine arts and landscape of Abruzzo as functionary paleontologist. She is head of the archeological and paleontological restoration laboratories. She is committed to the protection and enhancement of the geopaleontological sites and related museums.
 Silvano Agostini. Archaeological Superintendency of Abruzzo, via degli Agostiniani 14, Chieti 66100, Italy; silvano.agostini@beniculturali.it
Silvano Agostini. Archaeological Superintendency of Abruzzo, via degli Agostiniani 14, Chieti 66100, Italy; silvano.agostini@beniculturali.it
Silvano Agostini is a geologist. He works at the Superintendence for archaeology, fine arts and landscape of Abruzzo where he is coordinator for Education and Research. His research focuses on geology, geopaleontology, Quaternary geology, geoconservation, diagnostics, archeometry and GIS applications in the prehistoric ecology. His interests stretch to diagnostics and archeometry applied to restoration of archeological monuments, archeological sites and archeological finds in natural stone, as well as enviromental and hydrogeological restoration in the context of archeological parks.
FIGURE 1. Location map of Scontrone and Gargano. From Mazza (2013a), modified.
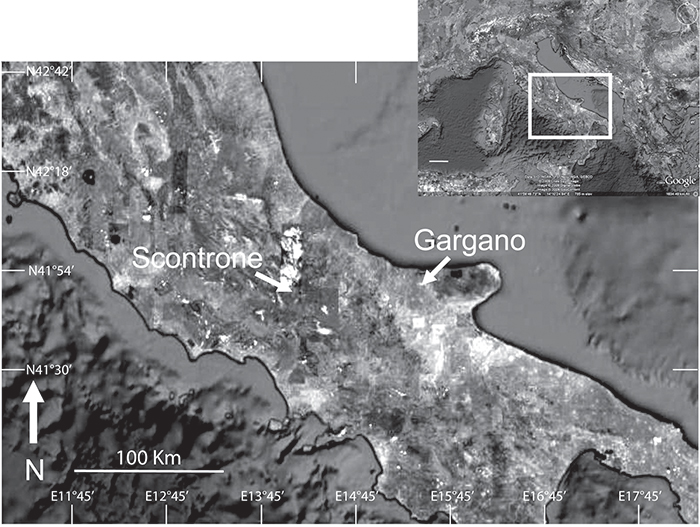
FIGURE 2. Schematic stratigraphy of the early Tortonian site of Scontrone (from Patacca et al., 2008a, modified). Succession showing the stratigraphic position of the bonebeds within the Scontrone Member of the Lithothamnium Limestone Formation. RC Rudist-bearing Calcarenite; SC Scontrone Member of the Lithothamnium Limestone Formation; SCa-SCd facies units of the Scontrone Member recording major shifts in the depositional setting and biotic associations (SCa = costal bar deposits; SCb = tidal creek deposits; SCc = marsh deposits; SCd = lagoon deposits). 1-Calcareous marls; 2-Bioclastic calcarenites; 3-Bioclastic calcarenites with oversized well-rounded lithoclast lags; 4-Lithoclastic calcirudites; 5-Major disconformity; 6-Low-angle cross-bedding; 7-Trough cross-bedding; 8-Root traces; 9-Oyster shell lags; 10-Hydrobiids; 11-Bonebeds; 12-Rudists. For further details see Patacca et al. (2008a).
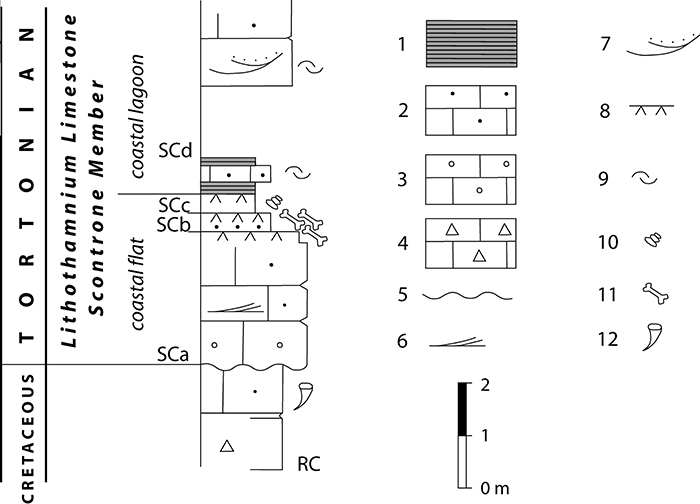
FIGURE 3. Guide to dental morphology terms used in this paper, drawn from Engesser (1980), Gould (1995), Lopatin (2006) by Masini and Fanfani (2013), and slightly modified here. 1, Upper molar. 2, Lower molar; (a) paralophid (paracristid) = paraconid crest + preprotocristid sensu Lopatin (2006); (b) postcristid (hypolophid) = postentocristid + posthypocristid sensu Lopatin (2006).
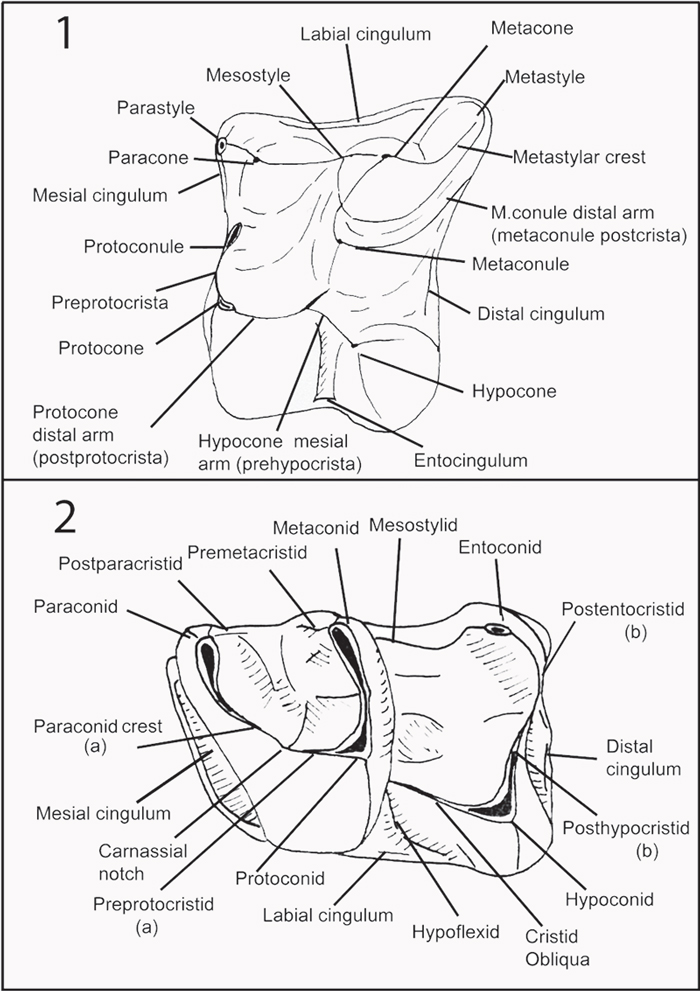
FIGURE 4. 1, Left M1 (F9-018), undetermined Deinogalerix sp. from fissure F9, oblique, occluso-distal view. 2, same as 1 with sketch showing contact of the distal arm of the metaconule with the uninterrupted distal cingulum. 3, left m1 (paratype SCT 347), Deinogalerix samniticus sp. nov., occlusal view. 4, same specimen as 3, lingual view. Arrows in 3 and 4 showing mesiolingual bulge. 5, left p4 (holotype SCT 246), Deinogalerix samniticus sp. nov., occlusal view. 6, same as 5, with sketch showing roundish, delimited lingually, trigonid valley. 7, right P3-P4 (paratype SCT 19), Deinogalerix samniticus sp. nov., occlusal view. 8, same as 7 with sketch showing the weak separation of protocone and hypocone. Hyc = Hypocone, Mecl = Metaconule, Mec = Metacone, Mecd = Metaconid, Pac = Paracone, Pacd = Paraconid, Prc = Protocone, Prcd = Protoconid. Figures not to scale.
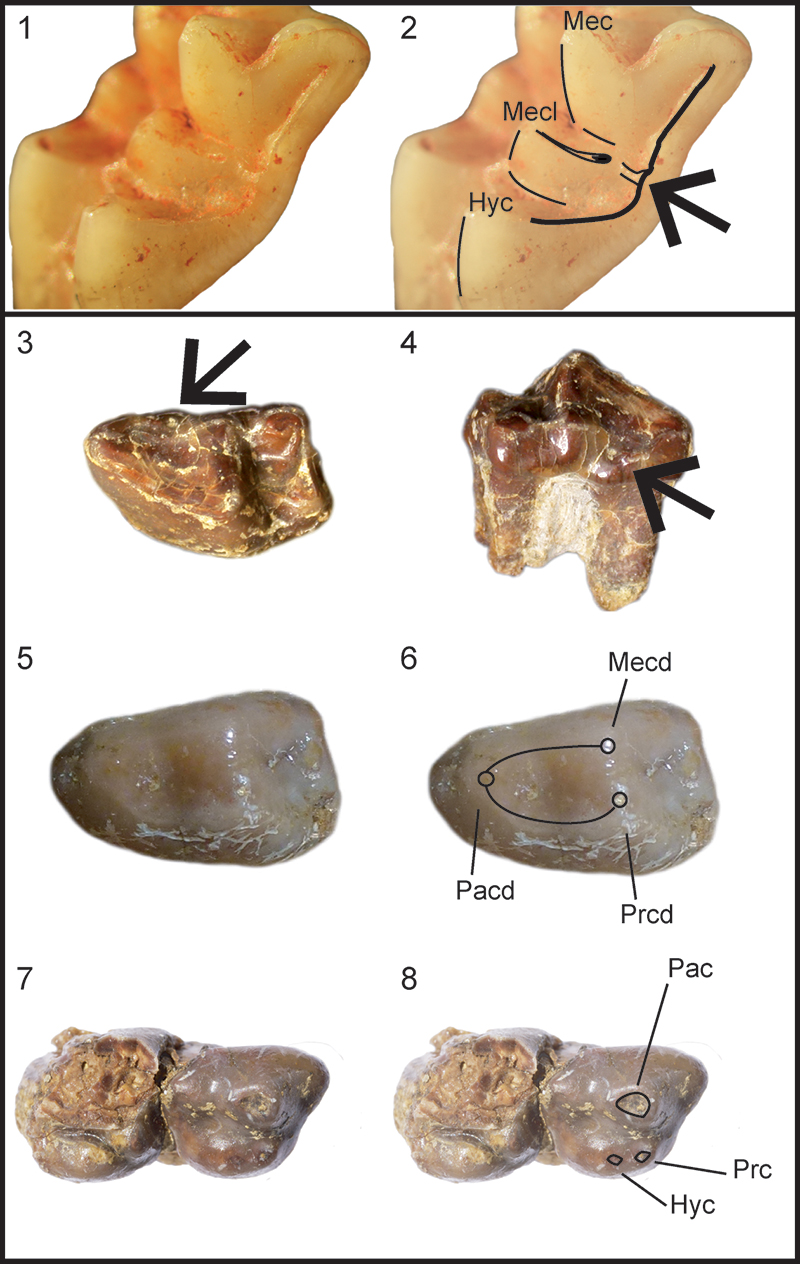
FIGURE 5. Deinogalerix samniticus sp. nov., from Scontrone. 1-2, Holotype, fragment of left hemimandible, with p3, p4, alveoli of p2, broken m1, m2, and m3 partially embedded in the rock (SCT 246). 1, labial view; 2, occlusal view. 3-5, Paratype, isolated left m1 (SCT 347). 3, occlusal view; 4, labial view; 5, lingual view. 6-7, Paratype, fragmental right maxillary with complete P3 and fragmented P4 (SCT 19). 6, occlusal view; 7, lingual view. 8, Paratype, fragmental right maxillary, with M3 and postero-lingual portion of M2 (SCT 232).

FIGURE 6.1-3, Holotype SCT 246 during its preparation from the rock matrix by vibrotool. The figure shows the premolars still partially encased in the rock matrix.

FIGURE 7. Comparisons of p3-m3 lengths in the different species of Deinogalerix, as well as in Parasorex, Galerix, Schizogalerix, and Apulogalerix (measurements and abbreviations in Table 3). 1, The graph shows that in Apulogalerix, Parasorex and other continental galericines molars grow progressively larger towards m1, which is the largest tooth of the row. The fourth lower premolar is larger than p3, which, in contrast, is the smallest of the toothrow (it is somewhat smaller than m3). Compared to the mainland counterparts, Deinogalerix bears a very large m1 (due to the increase of the trigonid length) and larger premolars, that grow progressively smaller moving rostrally, but less than in the continental genera (p3 intermediate in size between m1 and m2). The Scontrone specimen shows the size increase from m3 to m1 typical of Deinogalerix. In contrast, it bears a very large p4, similar in size to m1 and in the dimensional ranges of D. koenigswaldi, and a very reduced p3, more similar to those of the smaller and more primitive species of Deinogalerix from Gargano. 2, Ratio diagram comparing the p3-m3 lengths (see Table 4) in Deinogalerix samniticus sp. nov. with other species of Deinogalerix, using Parasorex socialis from La Grive as the standard (horizontal line). All the Deinogalerix specimens show a similar trend where the proportions of m2 and m3 are parallel to the standard, whereas, starting from m1, the teeth proportionally increase in size respect to the standard. It is noteworthy that this trend reaches its maximum expression in D. koenigswaldi. Compared to the other species of Deinogalerix, the curve of D. samniticus sp. nov.,shows a steeper rise toward p4, and it is the only one that drops toward p3.
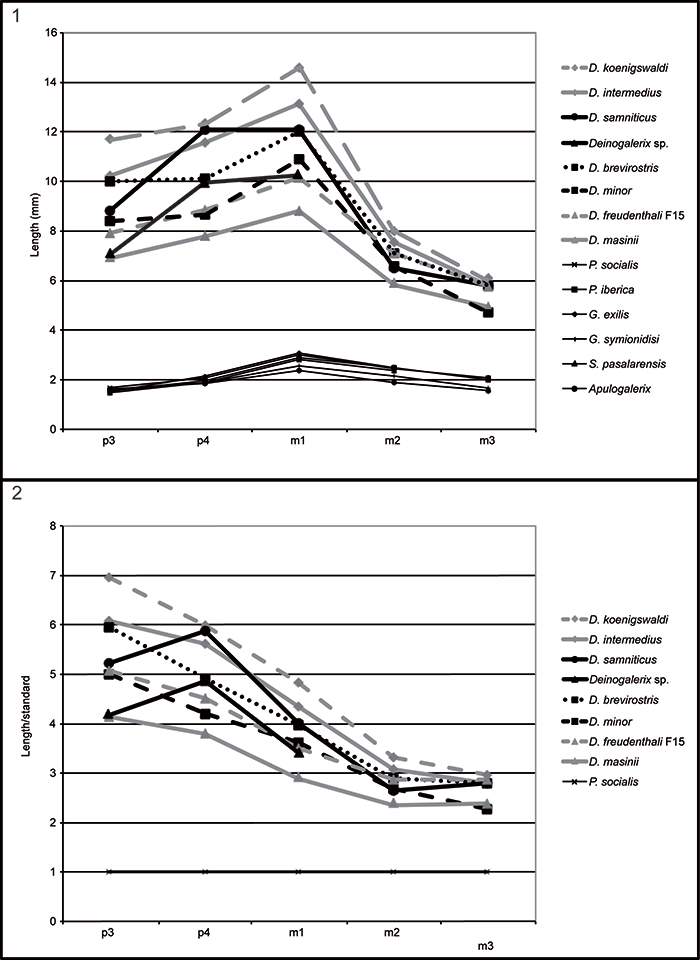
FIGURE 8. Scatter plot of Lp4 vs Lp3 (values in Table 5) for the species of Deinogalerix. The Deinogalerix specimens from Gargano align roughly in a linear trend. D. samniticus sp. nov. and Deinogalerix sp. in contrast, display an anomalously short p3. It is worth noting that also the smaller and more primitive species, e.g., D. freudenthali from F15 and D. masinii from M013, have fairly short p3s. Measurements in mm.
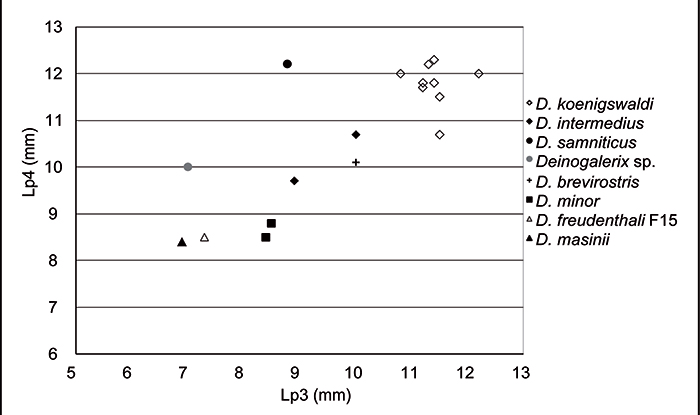
FIGURE 9. Left mandible (SCT 243), Deinogalerix sp. 1, lateral view; 2, dorsal view.

FIGURE 10. Histogram of the positions of the mental foramen in Deinogalerix and in other galericines. M = foramen located under the mesial root of p3 or p4, D = under the distal root of p3 or p4, C ("Central") = between the roots of p3 or p4, I ("Intermediate") = Foramen located between p3 and p4.
TABLE 1. Comparative material used for the present study; (h) = holotype; (p) = paratype.
| Species | Deposit | Inventory | Description |
| D. minor | Fina H | RGM 178184 (h) cast | fragmental right jaw with p2-m3 |
| Gervasio 1 | RGM 179174 (p) cast | fragmental left jaw with p3-m3 | |
| D. intermedius | Nazario 4 | RGM 179063 (h) cast | fragmental left jaw with p3-m1 |
| Chiro 20 C | RGM 177982 (p) cast | fragmental right maxillary with P3-M2 | |
| D. brevirostris | SG | RGM 179134 (h) cast | skull with right P1, both P2s, P3s, P4s, M1s, right M2-M3 |
| SG | RGM 179237 (p) cast | fragmental right premaxillary with I1, I3 | |
| SG | RGM 179232 (p) cast | fragmental left jaw with p3-m2 (broken m1) | |
| D. koenigswaldi | SG | RGM 177777 (h) cast | skull with both I1s, I3s, Cs, P1s, P3s, P4s, M1s, M2s, M3s and right P2 |
| SG | RGM 177778 (h) cast | fragmental left jaw with p1-m3 | |
| SG | RGM 177779 (h) cast | fragmental right jaw with p1-m3 | |
| SG | RGM 179194 cast | skull with right C, P1, P4 both P3s | |
| SG | RGM 179147 cast | fragmental right jaw with p3-m3 | |
| D. freudenthali | F15 | F15-015 | fragmental left jaw with p2-p4 |
| F15 | F15-016 | fragmental left jaw with m3 | |
| F15 | F15-031 | isolated left p4 | |
| F15 | F15-032 | isolated right M1 | |
| F15 | F15-033 | isolated right M2 | |
| F15 | F15-034 | isolated right m3 | |
| F15 | F15-036 | isolated left M3 | |
| F15 | F15-037 | isolated left M3 | |
| F15 | F15-038 | fragmental left maxillary with P3-M2 | |
| F15 | F15-039 | fragmental left jaw with m1, m3 | |
| F15 | F15-041 | isolated right P4 | |
| F15 | F15-042 | fragmental right jaw without teeth | |
| F15 | F15-043 | isolated right m1 | |
| F15 | F15-044 | isolated right m2 | |
| F15 | F15-045 | isolated right m2 | |
| F15 | F15-046 | isolated right p3 | |
| Deinogalerix sp. | F21c | F21-001 | isolated left M1 |
| F21c | F21-002 | isolated right p4 | |
| Deinogalerix sp. | P81D | P81D-001 | fragmental left jaw without teeth |
| P81D | P81D-002 | fragmental left jaw without teeth | |
| P81D | P81D-003 | fragmental right jaw without teeth | |
| P81D | P81D-004 | fragmental left jaw with m2-m3 | |
| P81D | P81D-009 | isolated left m1 | |
| P81D | P81D-023 | left jaw with p3 | |
| P81D | P81D-024 | left jaw with p3-p4 | |
| P81D | P81D-025 | fragmental right maxillary with P4 | |
| P81D | P81D-026 | isolated left P3 | |
| P81D | P81D-027 | isolated left p3 | |
| P81D | P81D-028 | isolated right p4 | |
| Deinogalerix sp. | NBS | NBS-001 | isolated left p4 |
| Deinogalerix sp. | F1 | F1-006 | isolated left P4 |
| F1 | F1-007 | isolated left P4 | |
| F1 | F1-009 | isolated left m2 | |
| Deinogalerix sp. | F8 | F8-040 | left jaw with c, p3-m3 (broken m1) |
| Deinogalerix sp. | F9 | F9-014 | fragmental skull with left C, both P1s, P3s, P4s, M2s and M3s |
| F9 | F9-017 | fragmental left maxillary with P1-P2, P4 | |
| F9 | F9-018 | isolated left M1 | |
| F9 | F9-022 | fragmental right jaw with p4 |
TABLE 2. Odontometrics of Deinogalerix samniticus sp. nov. and Deinogalerix sp. (SCT243). Measurement: L: length; W: width, Lm: minimum length (only M3), aL: alveolar length, H (m1, p2): height of mandible below m1 and p2, W m1: width of the mandible below m1. Italics: inferred value.
| Dental Measurements | ||||||||||
| L | W | L | W | L | W | L | W | L | W | |
| p3 | p4 | m1 | m2 | m3 | ||||||
| SCT246 | 8.8 | 5.0 | 12.1 | 7.6 | 12.25 | / | 6.5 | / | 5.8 | / |
| SCT243 | 7.1 | 3.5 | 10 | 6.7 | 10.3 | 6.5 | ||||
| SCT347 | 12.1 | 8 | ||||||||
| L | W | L | W | L | W | L | W | Lm | W | |
| P3 | P4 | M1 | M2 | M3 | ||||||
| SCT19 | 10.3 | 8.9 | / | 10.9 | ||||||
| SCT232 | 5.85 | 6.3 | ||||||||
| Mandible Measurements | ||||||||||
| aL | aL | aL | aL | H m1 | W m1 | H p2 | ||||
| m1-m3 | p2-p4 | p4-p3 | p4-p3 | |||||||
| SCT246 | 27.55 | 32.7 | 21.1 | 20.6 | 18.5 | 14.5 | >15 | |||
TABLE 3. Basic statistic parameters of lengths in mm of lower teeth (p3-m3) of the different Deinogalerix species, as well as in Parasorex, Galerix, Schizogalerix, and Apulogalerix . The mean values of Apulogalerix are actually grand means (i.e., mean of the means) of six different populations. The ranges have been approximated to the decimal digit to make them comparable to those reported by Butler (1980) and Doukas (1986). N = number of specimens; = mean values; σ = standard deviations. * Inferred from diagrams and figures. Raw data for Figure 7.1.
| Genus Species | Length | |||||||||
| Locality | Source | p3 | p4 | |||||||
| Deinogalerix | N | x̄ | σ | range | N | x̄ | σ | range | ||
| D. masinii | M013 | Villier, 2012; Villier et al., 2013 | 6 | 6.97 | 0.225 | 6.6-7.2 | 5 | 7.82 | 0.559 | 6.9-8.4 |
| D. freudenthali | F15 | This paper | 2 | 7.90 | 0.849 | 7.3-8.5 | 2 | 8.85 | 0.495 | 8.5-9.2 |
| D. minor | Various fissures | Butler, 1980 | 3 | 8.40 | 0.100 | 8.3-8.5 | 2 | 8.65 | 0.212 | 8.5-8.8 |
| D. brevirostris | SG Low | Butler, 1980 | 1 | 10.00 | / | / | 1 | 10.10 | / | / |
| D. samniticus | Scontrone | This paper | 1 | 8.80 | / | / | 1 | 12.10 | / | / |
| Deinogalerix sp. | Scontrone | This paper | 1 | 7.10 | / | / | 1 | 10.00 | / | / |
| D. intermedius | Various fissures | Butler, 1980 | 6 | 10.23 | 0.728 | 8.9-11 | 7 | 11.56 | 1.108 | 9.7-12.6 |
| D. koenigswaldi | SG, SG Low | Butler, 1980 | 6 | 11.70 | 0.360 | 11.3-12 | 7 | 12.34 | 0.560 | 11.5-13 |
| Galerix | ||||||||||
| G. symeonidisi | Aliveri | Doukas, 1986 | 3 | 1.56 | 1.5-1.6 | 14 | 1.90 | 1.7-2.1 | ||
| G. exilis | Goldberg | Ziegler, 1983 | 123 | 1.55 | 0.095 | 1.3-1.8 | 100 | 2.08 | 0.098 | 1.9-2.4 |
| Parasorex | ||||||||||
| P. socialis | La Grive | Masini & Fanfani, 2013 | 15 | 1.68 | 0.137 | 1.5-1.9 | 39 | 2.06 | 0.088 | 1.9-2.3 |
| P. ibericus | Otura 1 | Mein & Martin Suarez, 1993 | 2 | 1.56 | 1.5-1.6 | 3 | 1.96 | 1.9-2 | ||
| Schizogalerix | ||||||||||
| S. pasalarensis | Pasalar | Engesser, 1980 (*) | 1 | 1.48 | / | / | 7 | 1.88 | 0.040 | 1.8-1.9 |
| Apulogalerix | All fissures | this paper | 6 | 1.64 | 1.6-1.7 | 6 | 1.85 | 1.8-1.9 | ||
TABLE 3 (continued)..
| Length | |||||||||||
| m1 | m2 | m3 | |||||||||
| N | | σ | range | N | | σ | range | N | | σ | range |
| 9 | 8.77 | 0.308 | 8.3-9.2 | 10 | 5.88 | 0.467 | 5.3-6.5 | 5 | 4.86 | 0.363 | 4.1-5.5 |
| 2 | 10.14 | 0.325 | 9.9-10.4 | 2 | 7.10 | 0.566 | 6.7-7.5 | 3 | 5.77 | 0.221 | 5.5-5.9 |
| 2 | 10.90 | 0.141 | 10.8-11 | 3 | 6.57 | 0.208 | 6.4-6.8 | 1 | 4.70 | / | / |
| 1 | 12.00 | / | / | 2 | 7.05 | 0.071 | 7-7.1 | 1 | 5.80 | / | / |
| 2 | 12.18 | 0.106 | 12.1-12.25 | 1 | 6.50 | / | / | 1 | 5.80 | / | / |
| 1 | 10.30 | / | / | ||||||||
| 7 | 13.13 | 0.916 | 12-14.5 | 6 | 7.53 | 0.344 | 7.1-8 | 2 | 5.80 | 0.000 | 5.8-5.8 |
| 7 | 14.57 | 0.650 | 13.5-15.3 | 7 | 8.14 | 0.480 | 7.6-9 | 5 | 6.16 | 0.470 | 5.8-6.9 |
| 2 | 2.55 | 2.5-2.6 | 8 | 2.14 | 2-2.3 | 4 | 1.65 | 1.5-1.8 | |||
| 150 | 3.07 | 0.077 | 2.8-3.4 | 150 | 2.49 | 0.100 | 2.4-2.8 | 150 | 2.11 | 0.093 | 1.7-2.3 |
| 49 | 3.02 | 0.082 | 2.9-3.2 | 72 | 2.46 | 0.063 | 2.2-2.6 | 40 | 2.08 | 0.100 | 1.9-2.3 |
| 5 | 2.90 | 2.9-3 | 2 | 2.47 | 2.4-2.5 | 3 | 2.01 | 2-2.1 | |||
| 3 | 2.80 | 0.098 | 2.7-2.9 | 2 | 2.39 | 0.021 | 2.4-2.4 | ||||
| 6 | 2.39 | 2.3-2.5 | 6 | 1.91 | 1.8-2 | 6 | 1.57 | 1.5-1.7 | |||
TABLE 4. Ratios of lower teeth length (p3-m3) in Deinogalerix samniticus sp. nov. with other species of Deinogalerix respect to the standard (Parasorex socialis from La Grive). Raw data for Figure 7.2.
| Genus | Length ratio | |||||
| Species | Locality | p3 | p4 | m1 | m2 | m3 |
| Deinogalerix | ||||||
| D. masinii | Fissure M013 | 4.14 | 3.80 | 2.91 | 2.39 | 2.34 |
| D. freudenthali | Fissure F15 | 5.08 | 4.52 | 3.50 | 2.87 | 2.87 |
| D. minor | Various fissures | 4.99 | 4.20 | 3.61 | 2.68 | 2.26 |
| D. brevirostris | Fissure SG Low | 5.94 | 4.91 | 3.98 | 2.87 | 2.79 |
| D. samniticus sp. nov. SCT246 | Scontrone | 5.23 | 5.88 | 4.04 | 2.65 | 2.79 |
| Deinogalerix sp. | Scontrone | 4.22 | 4.86 | 3.41 | ||
| D. intermedius | Various fissures | 6.08 | 5.61 | 4.35 | 3.07 | 2.79 |
| D. koenigswaldi | Fissures SG and SG Low | 6.95 | 5.99 | 4.83 | 3.32 | 2.97 |
| Parasorex | ||||||
| P. socialis | La Grive | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
TABLE 5. Lengths of p3 and p4 of single specimens of Deinogalerix. Raw data for Figure 8.
| Species | Locality | Specimen | Source | Lp3 | Lp4 |
| D. masinii | Fissure M013 | PU128502 | Villier et al., 2013 | 6.9 | 8.4 |
| D. freudenthali | Fissure F15 | F15-015 | This paper | 7.3 | 8.5 |
| D. minor | Fissure Fina H | RGM178184 | Butler, 1980 | 8.5 | 8.8 |
| Fissure Gervasio | RGM179174 | Butler, 1980 | 8.4 | 8.5 | |
| D. intermedius | Fissure Nazario 4 | RGM179063 | Butler, 1980 | 10.0 | 10.7 |
| WG 354 294 | RGM178085/086 | Butler, 1980 | 8.9 | 9.7 | |
| D. brevirostris | Fissure SG Low | RGM179232 | Butler, 1980 | 10.0 | 10.1 |
| D. koenigswaldi | Fissure SG | RGM177778 | Villier, 2012 | 11.3 | 12.2 |
| Fissure SG | RGM177779 | Villier, 2012 | 11.2 | 11.8 | |
| Fissure SG | RGM179145 | Villier, 2012 | 10.8 | 12.0 | |
| Fissure SG | RGM179147 | Villier, 2012 | 11.4 | 12.3 | |
| Fissure SG | RGM179148 | Villier, 2012 | 12.2 | 12.0 | |
| Fissure SG | RGM179149 | Villier, 2012 | 11.4 | 11.8 | |
| Fissure SG | RGM179193 | Villier, 2012 | 11.5 | 10.7 | |
| Fissure SG Low | RGM179206 | Villier, 2012 | 11.2 | 11.7 | |
| Fissure SG Low | RGM179224 | Villier, 2012 | 11.5 | 11.5 | |
| D. samniticus sp. nov. | Scontrone | SCT 246 | This paper | 8.8 | 12.1 |
| Scontrone | SCT 243 | This paper | 7.1 | 10.0 |
New species of Deinogalerix (Mammalia, Eulipotyphla) from the late Miocene of Scontrone (Abruzzo, central Italy)
Plain Language Abstract
Deinogalerix is a giant hairy hedgehog that inhabited Scontrone (Abruzzo, central Italy) and Gargano (south-eastern Italy) during a time period approximately spanning 10 to 6 million years ago. At those times the two localities were part of a past island. The remains of Deinogalerix from Scontrone had never been described in detail before. Two new species are introduced herein. One of them is named Deinogalerix samniticus sp. nov., is larger sized and better documented, and is fully described in the paper. The other one is clearly distinct from D. samniticus sp. nov., but it is represented merely by a single mandible. It is therefore presented here as a different, but undetermined species, Deinogalerix sp. Comparisons show that the two new species have exclusive dental proportions that distinguish them from all the other species of Deinogalerix hitherto known and seems to exclude them from the direct ancestry of the Gargano representatives. Consistent with its early stratigraphic age, D. samniticus sp. nov. bears a suite of primitive characters.
The analysis of the giant hairy hedgehogs from Scontrone supports the assumption that Deinogalerix presumably descended from European relatives, similar to the representatives of the genus Parasorex, which date back to around 17–10 million years ago. On the other hand, the study seems to rule out that the giant Deinogalerix and Apulogalerix, i.e., a small hedgehog from Gargano, derived from a common ancestor. Given the age of the presumed ancestor of Deinogalerix, the study supports a hypothesis that the past island was not colonized in a short time lapse, but rather through different arrivals over a much longer time period.
Resumen en Español
Nuevas especies de Deinogalerix (Mammalia, Eulipotyphla) del Mioceno tardío de Scontrone (Abruzos, Italia central)
Deinogalerix es un componente típico de la fauna insular del Mioceno tardío de Gargano (sudeste de Italia). También se habían encontrado restos de este taxón durante los últimos 20 años en el yacimiento del Tortoniense temprano de Scontrone (Abruzos, Italia central), pero nunca se habían descrito en detalle. Se presentan dos nuevas formas. Una de ellas, D. samniticus sp. nov., es de mayor tamaño y está mejor documentada, y por ello es descrita formalmente. La otra forma es claramente distinta de D. samniticus sp. nov., pero está representada por una sola mandíbula. Por lo tanto, se presenta aquí como una especie diferente, pero no determinada, Deinogalerix sp. Los análisis comparativos muestran que las nuevas formas tienen proporciones dentales exclusivas que las distinguen de todas las otras especies de Deinogalerix y parecen excluirlas del ancestro directo del clado de Gargano. De acuerdo con su edad estratigráfica temprana, D. samniticus sp. nov. muestra una serie de caracteres primitivos del género. Además de estos, sin embargo, también muestra la proporción relativa entre p3 y p4 de naturaleza autapomórfica.
El análisis de los erinaceidos de Scontrone apoya la suposición de que Deinogalerix presumiblemente es descendiente de un hipotético galericino de tipo Parasorex del Mioceno temprano-medio. Por otro lado, parece descartar un ancestro común para el gigante Deinogalerix y el pequeño Apulogalerix de Gargano. Debido a que el presunto antecesor de Deinogalerix se remonta al Mioceno temprano-medio, el estudio corrobora la colonización asincrónica de la Plataforma de Apulia.
Palabras clave: Mioceno tardío; Abruzos (Italia central); endemismo; colonización insular; Deinogalerix; nueva especie
Traducción: Enrique Peñalver (Sociedad Española de Paleontología)
Résumé en Français
Une nouvelle espèce de Deinogalerix (Mammalia, Eulipotyphla) du Miocène récent de Scontrone (Abruzzes, Italie centrale)
Le genre Deinogalerix est un élément typique des faunes insulaires du Miocène récent de Gargano (sud-est de l'Italie). Ces 20 dernières années, des restes de ce taxon ont également été trouvés dans le site de Scontrone (Abruzzes, Italie centrale) datant du Tortonien ancien, mais ils n'avaient jamais été décrits en détail. Deux nouvelles espèces sont présentées. L'une d'entre elles, D. samniticus sp. nov., est de plus grande taille et mieux documentée, et reçoit une description officielle. L'autre est clairement distincte de D. samniticus sp. nov., mais elle n'est représentée que par une seule mandibule. Elle est donc ici présentée comme une espèce différente mais indéterminée, Deinogalerix sp. Des analyses comparatives montrent que les nouvelles espèces ont des proportions dentaires qui les distinguent de toutes les autres espèces de Deinogalerix et semblent les exclure du statut d'ancêtre direct du clade de Gargano. De manière cohérente avec son âge stratigraphique ancien, D. samniticus sp. nov. présente une suite de caractères plésiomorphes pour ce genre. Ce taxon montre cependant aussi une proportion relative des p3 et p4 autapomorphe. L'analyse des érinacéidés de Scontrone soutient l'hypothèse selon laquelle Deinogalerix descendrait d'un ancêtre hypothétique au Miocène ancien-moyen, un Galericinae semblable à Parasorex. D'un autre côté, l'analyse semble réfuter l'hypothèse selon laquelle le Deinogalerix géant et le petit gymnure Apulogalerix de Gargano partageraient un ancêtre commun proche. Comme l'ancêtre supposé de Deinogalerix daterait du Miocène ancien-moyen, cette étude corrobore une colonisation asynchrone de la Plateforme Apulienne.
Mots-clés : Miocène récent ; Abruzzes (Italie centrale) ; endémisme ; colonisation insulaire ; Deinogalerix ; nouvelle espèce
Translator: Antoine Souron
Deutsche Zusammenfassung
Eine neue Art von Deinogalerix (Mammalia, Eulipotyphla) aus dem späten Miozän von Scontrone (Abruzzen, Zentralitalien)
Deinogalerix ist eine typische Komponente der spätmiozänen Inselfauna von Gargano (südöstliches Italien). Es wurden zwar Überreste dieses Taxons über die letzten 20 Jahre auch in Sontrone (frühes Tortonium, Abruzzen, Zentralitalien) gefunden, jedoch wurden diese nie detailliert beschrieben. Es werden zwei neue Arten vorgestellt, von denen D. samniticus sp. nov., größer ist und besser dokumentiert ist und eine formale Beschreibung erhielt. Das andere Stück unterscheidet sich klar von D. samniticus sp. nov., besteht aber nur aus einer einzigen Mandibel. Es wird hier daher als die unterschiedliche aber unbestimmte Art Deinogalerix sp. vorgestellt. Vergleiche zeigen, dass die neuen Arten einzigartige Zahnproportionen aufweisen, welche sie von allen anderen Deinogalerix-Arten unterscheiden und die sie von der direkten Abstammungslinie der Gargano-Klade ausschließen. In Übereinstimmung mit seinem frühen stratigrafischen Alter zeigt D. samniticus sp. nov eine Reihe primitiver Merkmale dieser Gattung. Daneben weist sie die autapomorphen relativen Proportionen zwischen p3 und p4 auf.
Die Untersuchung der Erinaceiden aus Scontrone unterstützt die Annahme, dass Deinogalerix möglicherweise von einem früh-mittelpleistozänen 'Parasorex-artigen' Galericinen abstammt. Auf der anderen Seite scheint damit die Annahme eines gemeinsamen Vorfahren von Riesen- Deinogalerix und der kleinen Gymnure Apulogalerix aus Gargano ausgeschlossen zu werden. Da der angenommene Vorfahre von Deinogalerix bis ins Früh-Mittlelmiozän zurückreicht, unterstützt diese Untersuchung die asynchrone Besiedelung Apuliens.
Schlüsselwörter: spätes Miozän; Abruzzen (Zentralitalien); Endemismus; Inselbesiedelung; Deinogalerix; neue Art
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024