

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
APPENDIX
List of characters used in cladistics analysis (matrix provided as electronic appendage)
The data are largely based on Yan et al. (2017). One character (prosternal grooves for protarsi) was removed as it is only informative within Cupedidae. Characters 18 and 19 were added. The taxon sampling was slightly modified. Two ademosynid genera were added and few terminals from the extant suborders were removed of replaced.
- Externally visible membranes: (0) present; (1) absent. Largely or completely absent in fossil beetles and also in extant groups with very few exceptions (e.g., Ponomarenko, 1969; Beutel et al., 2008).
- Tubercles: (0) absent or indistinct; (1) present. Distinct in Cupedidae and Ommatidae, and also in stem group Coleoptera (Beutel et al., 2008). Absent in †Ademosynidae, †Catiniidae, †Peltosynidae, †Schizophoridae (partim; Ponomarenko, 1969) and in extant groups.
- Scale-like setae: (0) absent; (1) present. Absent in all species of †Ademosynidae, in other extinct groups lacking cuticular tubercles, in extant Archostemata excl. Cupedidae and Ommatidae, and in extant non-archostematan beetles.
- Punctuation of body surface: (0) punctures absent or with lower density; (1) dense punctuation. Very dense punctuation present in †Ademosynidae, Archosyne and †Peltosynidae. Unusually coarse in Peltosyne (Yan et al., 2017).
- Ocelli: (0) three; (1) absent. Absent in †Ademosynidae and †Peltosynidae, and also missing in almost all other extant and extinct groups.
- Shape of head capsule: (0) not distinctly transverse; (1) distinctly transverse. Distinctly transverse head in †Peltosynidae and few extant groups.
- Constricted neck and postocular extensions: (0) absent or indistinct; (1) present. Absent in †Ademosynidae and most other extant and extinct groups. Usually present in Archostemata (e.g., Beutel et al., 2008)
- Supraantennal protuberance (P1): (0) absent; (1) present. Present as moderately distinct bulge above antennal base in Ommatidae and as distinct protuberance in extant Cupedidae (Beutel et al., 2008; Hörnschemeyer, 2009).
- Supraocular protuberance (P2): (0) absent; (1) present as moderately distinct bulge; (2) present as strongly pronounced protuberance. Distinct protuberance in most extant Cupedidae (Beutel et al., 2008), moderately distinct supraocular bulge in some other archostematan groups (e.g., Pace, 1975) and in Archosyne and Sylvacoleus (Ponomarenko, 1969).
- Posteromesal protuberance (P3): (0) absent; (1) present, moderately convex; (2) conspicuous, strongly convex. Present in Cupedidae and Tetraphalerus and Sikhotealinia (Beutel et al., 2008). Absent in fossil taxa under consideration (Ponomarenko, 1969).
- Posterolateral protuberance (P4): (0) absent; (1) present. Present on posterolateral head region in some genera of Cupedidae (Beutel et al., 2008).
- Cephalic antennal groove; (0) absent; (1) below compound eye; (2) above compound eye. Present below compound eyes in Tetraphalerus and Peltosyne (Yan et al., 2017), and above it in Crowsoniella (Pace, 1975) and Jurodidae (Kirejtshuk, 1999). Absent in Omma, Cupedidae and Micromalthus, and also missing in coleopteran stem group taxa with the possible exception of †Rhombocoleidae (Ponomarenko, 1969).
- Number of antennomeres: (0) 13 or more; (1) 11 or less. Thirteen in †Tshercardocoleidae, †Permocupedidae and †Rhombocoleidae), but eleven in other fossil taxa with preserved antennae (Ponomarenko, 1969; Beutel et al., 2008). Eleven or less in extant beetles.
- Location of antennal insertion on head capsule: (0) laterally; (1) dorsally. Laterally in †Ademosynidae like in most other most extant beetles and fossils beetles (Ponomarenko, 1969; Beutel et al., 2008). On dorsal side in Cupedidae excl. Priacma (Beutel et al., 2008) and in Sikhotealinia (Kirejtshuk, 1999).
- Mandibular mola: (0) absent; (1) present. Present in Myxophaga and many basal polyphagan groups (e.g., Lawrence et al., 2011). Also present in †Peltosynidae (Yan et al., 2017). Absent in Archostemata and Adephaga (Beutel et al., 2008). Not verifiable in most fossils including ademosynid genera.
- Cutting edge of mandible: (0) horizontal, (1) three vertically arranged teeth. Three vertically arranged teeth in Ommatidae and Micromalthus (e.g., Beutel et al., 2008). Cutting edge almost always horizontal in other groups (e.g., Dolichosyne, Peltosyne).
- Lateral mental lobes enclosing prementum: (0) absent; (1). Distinctly developed in Adephaga (e.g., Dressler et al., 2010). Absent in other extant and extinct beetles.
- Anterior pronotal margin: (0) not distinctly convex and not overlapping posterodorsal head region; (1) distinctly convex and overlapping posterodorsal head region. The convex margin overlapping the posterior head region is an unusual and characteristic feature of †Ademosynidae.
- Anterolateral pronotal angles: (0) present; (1) reduced. Almost always distinctly developed (e.g., Ponomarenko, 1969) but reduced in †Ademosynidae. A tendency towards reduction of the anterolateral angle can be also observed in Ommatidae and Cupedidae, and it is entirely missing in Omma rutherfordi Lawrence, 1999 (Lawrence, 1999) and largely obliterated in Tetraphalerus bruchi Heller, 1931 (Friedrich et al. 2008). However, the conditions found in these families differ distinctly from what we observed in ademosynids, and a more or less distinctly recognizable anterolaterally or laterally directed angle is usually present (e.g., Ponomarenko 1969; Baehr, 1979; Tan et al., 2012) (coded as 0 for general of Ommatidae and Cupedidae).
- Propleural suture (0) present; (1) absent. Present in Chauliodinae (Maki, 1936), †Tshecardocoleidae, †Permocupedidae, and †Triadocupedinae (Ponomarenko, 1969). Condition in †Rhombocoleidae unclear. Absent in other beetles.
- Exposure of propleura: (0) largely or fully exposed; (1) internalized. Internalized in Polyphaga and also in † Peltosyne, † Petrosyne and † Ranis (Yan et al., 2017).
- Broad prothoracic postcoxal bridge: (0) absent; (1) present. Present in †Tshecardocoleidae, †Permocupedidae and †Rhombocoleidae (Ponomarenko, 1969; Beutel et al., 2008). Also developed in some few groups not included here (e.g., Rhysodini).
- Mesocoxal cavities: (0) not bordered by metanepisterum; (1) bordered by metanepisternum. Metanepisternum forms part of lateral border of mesocoxal cavities in Cupedidae, Ommatidae, †Jurodidea, Myxophaga, and Derodontidae (Kirejtshuk, 1999; Lawrence et al., 2011).
- Separation of mesocoxae: (0) moderately widely separated or adjacent; (1) very widely separated. Very widely separated in Myxophaga (e.g., Beutel, 1999) and a species of Ranis (coded as 0&1 for the genus).
- Transverse suture of mesoventrite: (0) present; (1) absent. Present in Cupedidae, Ommatidae and Sikhotealinia, and also in some fossil taxa under consideration including † Peltosyne (Yan et al., 2017). Absent in †Catiniidae, †Ademosynidae (in contrast to Ponomarenko, 1969: figure 71c) and † Ranis.
- Exposure of metatrochantin: (0) externally visible; (1) internalized or absent. Externally visible in Cupedidae and Ommatidae, and also in the fossil taxa under consideration with the exception of †Catiniidae and †Schizophoridae (narrow element may be exposed in † Catinius; Ponomarenko, 1969).
- Shape of penultimate tarsomere: (0) not distinctly bilobed; (1) distinctly bilobed. Bilobed in Cupedidae, Decliniidae and Sialidae (e.g., Lawrence, 1999; Yan et al., 2017).
- Sclerotization of fore wings: (0) membranous; (1) transformed into sclerotized elytra. Elytra with epipleura present in beetles with the exception of very few specialized forms (e.g., Beutel and Haas, 2000).
- Venation of fore wings: (0) with distinctly curved veins; (1) without distinctly curved veins. Distinctly curved veins only preserved in †Protocoleoptera (e.g., †Tshecardocoleidae; Ponomarenko, 1969).
- Elytral sclerotisation pattern: (0) pattern of unsclerotized window punctures; (1) entirely sclerotized. Window puncture pattern present in Cupedidae and Ommatidae, and also in most stem group Coleoptera (e.g., Ponomarenko, 1969). Absent in †Ademosynidae and some other extinct groups, and generally missing in non-archostemataqn beetles (e.g., Ponomarenko, 1969, 2004; Beutel et al., 2008; Yan et al., 2017).
- Elytral striae: (0) absent; (0) present. Present in †Ademosynidae, †Peltosynidae, Ranis, Archosyne and some other extinct taxa (Yan et al., 2017). Also occuring in some groups of Adephaga (e.g., Carabidae, major part) or Polyphaga (e.g., Histeridae partim, Elateridae partim) (Lawrence et al., 2011).
- Elytral apex: (0) distinctly reaching beyond abdominal apex posteriorly; (1) slightly reaching beyond abdominal apex posteriorly; (2) reaching abdominal apex or shorter. Distinctly reaching beyond abdominal apex in †Tshecardocoleidae and slightly in †Permocupedidae (Ponomarenko, 1969; Beutel, 1997; Beutel et al., 2008). Closely adapted to shape of abdomen in extant beetles with few exceptions.
- Abdominal sternite I: exposed; (1) concealed under metacoxae, largely or completely reduced. Reduced in all extant and fossil beetles (e.g., Beutel and Haas, 2000).
- Connate abdominal ventrites: (0) none or at least less than 3; (1) 3. Three sternites connate in Adephaga (e.g., Lawrence et al., 2011).
- Median ridge on ventrite 1: (0) absent; (1) present. Present in Sikhotealinia, † Jurodes, Cupedidae and Ommatidae (e.g., Beutel et al., 2008).
- Arrangement of abdominal sterna: (0) abutting, not overlapping; (1) tegular or overlapping. Overlapping in Cupedidae (Lawrence et al., 2011; Beutel et al., 2008).
 Evgeny V. Yan. Institut für Spezielle Zoologie und Evolutionsbiologie, Friedrich Schiller University Jena, 07737, Germany and Paleontological Institute, Russian Academy of Sciences, Profsoyuznaya str. 123, Moscow, 117997 Russia. yan-e@mail.ru
Evgeny V. Yan. Institut für Spezielle Zoologie und Evolutionsbiologie, Friedrich Schiller University Jena, 07737, Germany and Paleontological Institute, Russian Academy of Sciences, Profsoyuznaya str. 123, Moscow, 117997 Russia. yan-e@mail.ru
Scientific interests: early evolution of Coleoptera, paleontological reocnstructions.
Evgeny Yan received his Ph.D. at Palaeontological Institute, Russian Academy of Sciences. Currently he works as the visiting scientist in Institute of Special Zoology and Evolutionary Biology (Jena) and have a postion of Senior research assistant in Palaeontological Institute (Arthrhopod laboratory, Moscow).

 Rolf G. Beutel. Institut für Spezielle Zoologie und Evolutionsbiologie, Friedrich Schiller University Jena, 07737, Germany. rolf.beutel@uni-jena.de
Rolf G. Beutel. Institut für Spezielle Zoologie und Evolutionsbiologie, Friedrich Schiller University Jena, 07737, Germany. rolf.beutel@uni-jena.de
Scientific interests: Coleoptera macrosystematics; detailed morphological studies of Arthropoda morphology.
Rolf Georg Beutel received his PhD at the University of Tübingen. Presently he is Professor of Zoology at the Friedrich-Schiller-Universität Jena and also responsible for the scientific collection of the Phyletisches Museum. He has published an entomological textbook (Insect Morphology and Phylogeny, Beutel, Friedrich, Ge & Yang 2014) and more than 200 original studies. He is editor of the Handbook of Zoology series (Insecta).
 Alexander G. Ponomarenko. Paleontological Institute, Russian Academy of Sciences, Profsoyuznaya str. 123, Moscow, 117997 Russia. aponom@paleo.ru
Alexander G. Ponomarenko. Paleontological Institute, Russian Academy of Sciences, Profsoyuznaya str. 123, Moscow, 117997 Russia. aponom@paleo.ru
Scientific interests:evolution of insects (with particular interest in Coleoptera); evolutionary theory.
He received his Ph.D. at Palaeontological Institute, USSR Academy of Sciences. Prof. Ponomarenko is Leading research assistant in Palaeontological Institute (Laboratory of ancient organisms, Moscow).
FIGURE 1. Dolichosyne Ponomarenko, 1969 habitus photos. 1, D. confragosa PIN 2785-2675 new material. 2, 3, D. confragosa paratype PIN 2070-1551 part and counterpart. 4, D. rostrata holotype PIN 2555/1735. 5, D. sulcata holotype PIN 2240-274 part. 6, D. sulcata paratype PIN 2240-234 part. 7, D. sulcata paratype PIN 2240-305 part. 8, D. sulcata PIN 2785-2659. Scale bars equal 1 mm.

FIGURE 2. Dolichosyne Ponomarenko, 1969 line drawings. 1, D. confragosa PIN 2785-2675. 2, 3, D. confragosa paratype PIN 2070-1551 part and counterpart. 4, D. rostrata holotype PIN 2555/1735. 5, D. sulcata holotype PIN 2240-274 part. 6, D. sulcata paratype PIN 2240-234 part. 7, D. sulcata paratype PIN 2240-305 part. 8, D. sulcata PIN 2785-2659. Scale bars equal 1 mm.
Label abbreviations: c.e - compound eye; cx1, 2 - pro- and mesocoxae; eps3 - metepisternum; gul - gular plate (gula); i.p1 - prosternal intercoxal process; l.d - pronotal longitudinal depression; l.i - lateral incisions of submentum; l.r - longitudinal ridge; msc - mesoscutum; mt - mentum; par - parietal; pl1 - propleurit; p.mt - prementum; p.t.p. - posterior tentorial pit; scl - scutellum; sct - mesoscutum; smt - submentum; s.scl - subocular suture; tr1 - protrochanter; tr3 - metatrochanter.
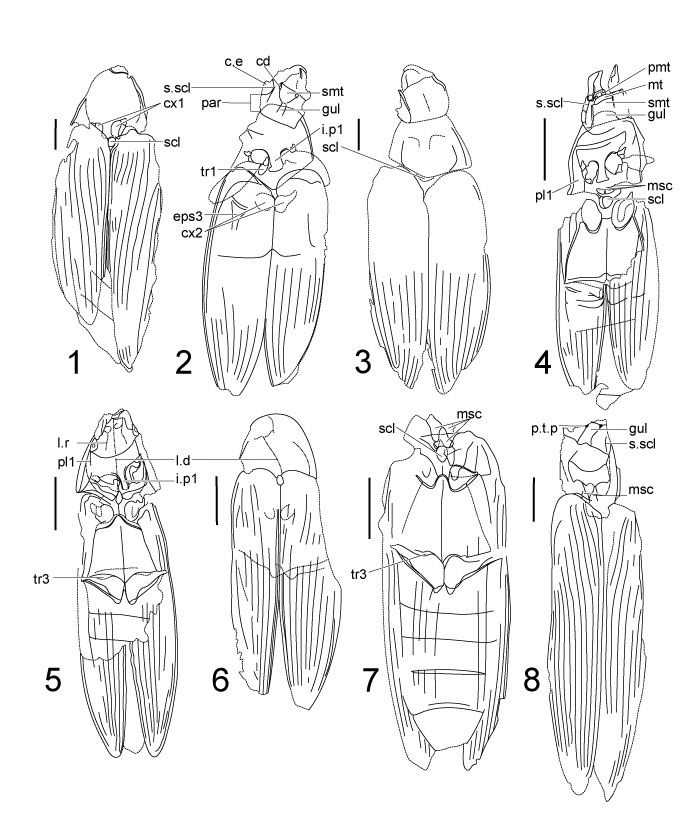
FIGURE 3. Genera of Ademosynidae family, habitual photos. 1, Gnathosyne akkolkensis Ponomarenko, 1969, holotype, PIN 2496/7 counterpart. 2, Ademosyne kirgizica Ponomarenko, 1969, holotype PIN 2096/1369. 3, 4, A. kirgizica Ponomarenko, 1969 paratype PIN 2555/1696 print and counterprint. 5, A. bacca Ponomarenko, 1969, holotype PIN 2240/310. 6, A. bacca Ponomarenko, 1969, paratype PIN 2240/294. 7, A. ellyptica Ponomarenko, 1969, holotype PIN 2240/224. 8, Cephalosyne capitata Ponomarenko, 1969 holotype PIN 2240/302. Scale bars equal 1 mm.

FIGURE 4. Genera of Ademosynidae family, line drawings. 1, Gnathosyne akkolkensis Ponomarenko, 1969, holotype, PIN 2496/7 counterpart. 2, Ademosyne kirgizica Ponomarenko, 1969, holotype PIN 2096/1369. 3 A. kirgizica Ponomarenko, 1969, paratype PIN 2555/1696 print and counterprint. 4, A. bacca Ponomarenko, 1969 holotype PIN 2240/310. 5, A. bacca Ponomarenko, 1969, paratype PIN 2240/294. 6, A. ellyptica Ponomarenko, 1969, holotype PIN 2240/224. 7, Cephalosyne capitata Ponomarenko, 1969, holotype PIN 2240/302. Scale bars: 1 mm.
Label abbreviations: c.e - compound eye; crd - cardo; cx.r - coxal rests; cx1, 2 - pro- and mesocoxae; ep - epipleura; epm2 - mesepimeron; fm1 - profemur; gul - gular plate (gula); i.p1 - prosternal intercoxal process; l.d - pronotal longitudinal depression; l.i - lateral incixions of submentum; l.r - longitudinal ridge; md - mandibles; msc - mesoscutum; mt - mentum; ms.p - mesosternal pit; o.f - occipital foramen; par - parietal; pd - pedicellus; pl1 - propleurite; p.mt - prementum; p.t.p. - posterior tentorial pit; sc - scapus; scl - scutellum; sct - mesoscutum; s.g - gular suture; s.l2 - longitudinal suture of mesoventrite; s.l3 - longitudinal suture of metaventrite; smt - submentum; s.pc - paracoxal suture.
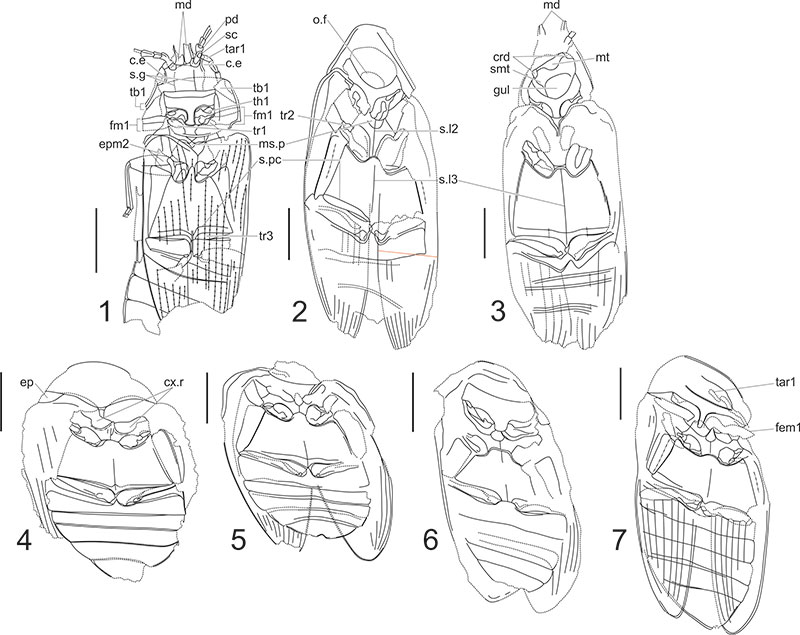
FIGURE 5. Petrosyne Ponomarenko, 1969 and ademosynid beetles re-assigned to Polyphaga suborder, photos and line drawings. 1, 2 Petrosyne liassica Ponomarenko, 1969, holotype PIN 166/39. 3, 4 Ranis collevus Ponomarenko, 1969, holotype PIN 371/1755 and line drawing. 5, 6, R. ovalis Ponomarenko, 1969, holotype PIN 2066/2642. Scale bars equal 1 mm.
Label abbreviations: cx.r - coxal rests; msc - mesoscutum; tb1 - protibia.

FIGURE 6. Comparative morphology of †Ademosynidae, polyphagan beetles and stem group Coleoptera. 1, 2, Stenocyphon sasaji Lawrence, 2001. 3, 4, Epiphanis cornutus (Eschscholtz, 1829). 5, 6, Dolichosyne sulcata Ponomarenko, 1969. 7, 8 Gnathosyne akkolkensis Ponomarenko, 1969. 9, 10, Archosyne permiana Ponomarenko et al., 2014. 11, 12, Tecticupes heckeri Rohdendorf, 1961 (†Taldycupedidae). 13, 14, Permocupes sojanensis Ponomarenko, 1963 (†Permocupedidae). Scale bars equal 1mm. Label abbreviations: mb.c - membranous connection.
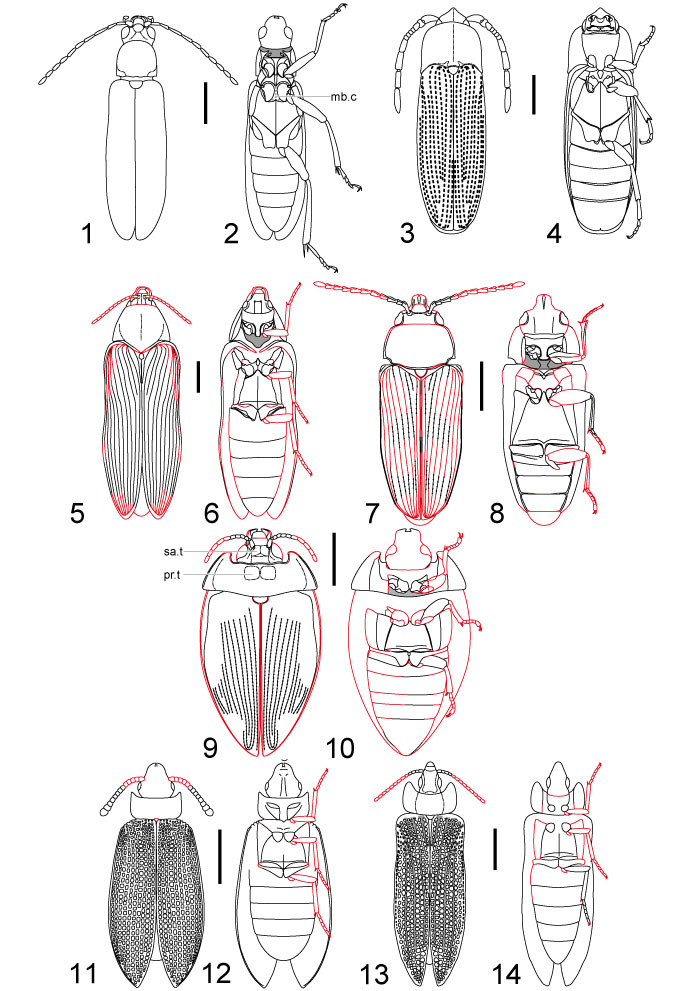
FIGURE 7. Strict consensus tree of 71 minimum length trees (NONA, ratchet, 1000 replicates, all characters unweighted and unordered), 91 steps, consistency index 0.43. Monopyhyly of †Ademosynidae supported by two unambiguous apomorphies: anterior pronotal margin convex and overlapping posterior part of head (18.1), anterolateral pronotal angles absent (19.1). Systematic position of †Ademosynidae and Archosyne ambiguous.
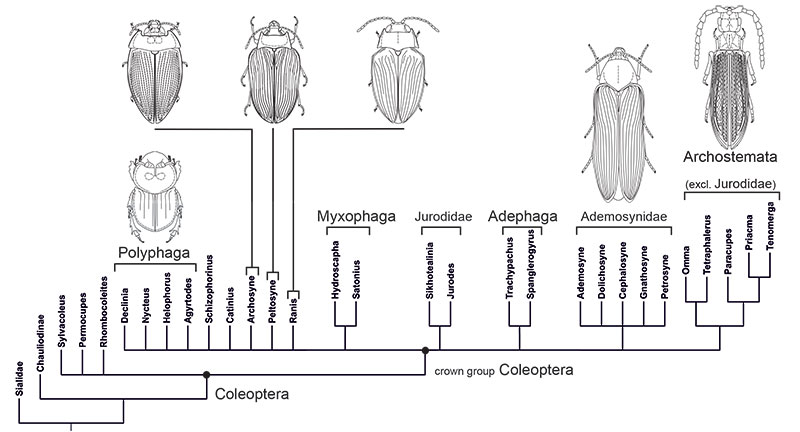
TABLE 1. Body measurements of †Ademosynidae. All dimensions given in millimetres.
| Species | Dolichosyne confragosa Ponomarenko, 1969 |
D. rostrata Ponomarenko, 1969 |
D. sulcata Ponomarenko, 1969 |
Gnathosyne akkolkensis Ponomarenko, 1969 |
Ademosyne kirgizica Ponomarenko, 1969 |
A. bacca Ponomarenko, 1969 |
A. ellyptica Ponomarenko, 1969 |
Cephalosyne capitata Ponomarenko, 1969 |
|
| Specimen number | PIN 2785-2675 | PIN 2070-1551 | PIN 2555/1735 | PIN 2240-274 | PIN 2496/7 | PIN 2096/1369 | PIN 2240/310 | PIN 2240/224 | PIN 2240/302 |
| Body | |||||||||
| length | 10 | 10,4 | 5,6 | 6,7 | 4,5 | 6,2 | 3,8 | 4,2 | 5,4 |
| width | 3,5 | 3,8 | 1,6 | 1,8 | 1,7 | 2,6 | 3 | 2,1 | 2,5 |
| Elytron | |||||||||
| length | 7,8 | 7,1 | 3,1 | 4,8 | 2,9 | 5,2 | ~ | ~ | 3,8 |
| width | 1,8 | 2,1 | 0,85 | 0,9 | ~ | 1,2 | ~ | ~ | 1 |
| Head | |||||||||
| length | ~ | 1,7 | 1,1 | 0,9 | 0,8 | ~ | ~ | ~ | ~ |
| width | ~ | 1,6 | 0,75 | 0,9 | 0,7 | ~ | ~ | ~ | ~ |
| Pronotum | |||||||||
| length | 2 | 2,7 | 1,1 | 1,1 | 0,9 | 1,3 | 0,7 | 0,8 | 0,7 |
| width | 2,4 | 2,6 | 1,4 | 1,6 | 1,2 | 1,7 | 2 | 2 | 1,8 |
| Abdomen | |||||||||
| length | ~ | ~ | 2 | 3,2 | 1,3 | 2,1 | 1,5 | 1,4 | 2,3 |
| width | ~ | ~ | ~ | 1,6 | 1,4 | 2,1 | 2,3 | 1,8 | 2,1 |
| Foreleg length | |||||||||
| femur | ~ | ~ | ~ | ~ | 0,7 | ~ | ~ | ~ | ~ |
| tibia | ~ | ~ | ~ | ~ | 0,6 | ~ | ~ | ~ | ~ |
| tarsus | ~ | ~ | ~ | ~ | 0,4 | ~ | ~ | ~ | ~ |
| Middle leg length | |||||||||
| femur | ~ | ~ | ~ | ~ | 0,7 | ~ | ~ | ~ | ~ |
| tibia | ~ | ~ | ~ | ~ | 0,7 | ~ | ~ | ~ | ~ |
| tarsus | ~ | ~ | ~ | 0,1 | ~ | ~ | ~ | ~ | |
| Hind leg length | |||||||||
| femur | ~ | ~ | ~ | ~ | 0,6 | ~ | ~ | ~ | 0,9 |
| tibia | ~ | ~ | ~ | ~ | ~ | ~ | ~ | ~ | 0,7 |
| tarsus | ~ | ~ | ~ | ~ | ~ | ~ | ~ | ~ | ~ |
TABLE 2. Genera of †Ademosynidae.
| 1. Prosternal intercoxal process parallel-sided, or widening apically, often with lateral tongue-like projections at its apex; elongate-oval beetles, body length exceeds maximum width 2,5-3 times (except some Ademosyne) | 2 | |
| - Intercoxal process acute, narrowing posteriorly; transverse, stout beetles, body at most two times as long as maximum width. Mesocoxae widely separated by anterior process of metaventrite; metanepisterna with thick median longitudinal ridge | Cephalosyne Ponomarenko, 1969 | |
| 2. Anterior prosternal margin almost straight, very slightly concave; metacoxae three times as wide as long, not narrowing laterally | Gnathosyne Ponomarenko, 1977 | |
| - Anterior prosternal margin distinctly curved; metacoxae narrowing laterally | 3 | |
| 3. Anterior margin of metanepisterna straight; anterior process of metaventrite elevated | Petrosyne Ponomarenko, 1969 | |
| - Outer angles of metanepisterna protruding forward; anterior process of metaventrite not elevated | 4 | |
| 4. Head hypognathous, less than half as wide as posterior pronotal margin. Body strongly convex, almost as long as wide | Sphaerosyne, Ponomarenko, 1969 | |
| - Head prognathous, less than twice as narrow as basal margin of pronotum; body elongated | 5 | |
| 5. Protrochantins more or less rectangular, length and width equal; apex of prosternal intercoxal process usually with lateral protrusions (if not, then mandibles with distinctly curved apices and pronotum with median longitudinal depression); metacoxal width two times exceeding length, coxae with maximum length medially | Dolichosyne Ponomarenko, 1969 | |
| - Protrochantins transverse, more than three times as wide as long; metacoxal width more than four times exceeding length, coxae weakly narrowing laterally | Ademosyne Ponomarenko, 1969 | |
Ademosynidae (Insecta: Coleoptera): A new concept for a coleopteran key taxon and its phylogenetic affinities to the extant suborders
Plain Language Abstract
Extinct beetles from the family †Ademosynidae existed from the Late Permian until the Early Jurassic. In this study they are re-described using modern paleontological techniques. Based on well-documented and phylogenetically evaluated characters we present a new concept of the family. Six genera share two unusual evolutionary novelties and are therefore a well-defined monophyletic unit. †Ademosynidae reflect an intermediate stage in a crucial period in beetle evolution, with some "primitive" features of the ancestral suborder Archostemata retained, but other derived characteristics shared with "modern" beetles, especially the megadiverse suborder Polyphaga. Our paleontological investigations yield detailed information on more than 200-million-year-old beetles and illustrate evolutionary transitions, also including parallel evolution in †Ademosynidae and the structurally advanced Polyphaga.
Resumen en Español
Ademosynidae (Insecta: Coleoptera) - un nuevo concepto para un taxón clave de coleópteros y sus afinidades filogenéticas con los subórdenes no extintos
Ademosynidae es una controvertida familia extinta de Coleoptera, descubierta por primera vez en los depósitos australianos del Triásico Superior. El último representante conocido es del Jurásico temprano de Kirguistán. Se redescribe y se interpreta filogenéticamente el material tipo de la colección del Instituto Paleontológico de Moscú. Los resultados muestran que la familia tal como se define actualmente no es monofilética, sino que posiblemente está dividida entre dos de cuatro subórdenes de coleópteros no extintos o existentes. Se establece un nuevo concepto más restringido de la familia, incluyendo los géneros Ademosyne Handlirsch, 1906, Dolichosyne Ponomarenko, 1969, Gnathosyne Ponomarenko, 1969, Cephalosyne Ponomarenko, 1969, Petrosyne Ponomarenko, 1969, y Sphaerosyne Ponomarenko, 1969. Esta unidad monofilética se caracteriza por dos sinapomorfías del protórax, la ausencia de ángulos antero-laterales del pronoto y un margen anterior del pronoto prominente y redondeado que cubre la parte posterior de la cabeza. Otros caracteres adicionales son las mandíbulas grandes que claramente sobresalen anteriormente, élitros estriados, un ángulo pronoto-elitral indistinto dando como resultado un cuerpo rayado-alineado, y una superficie cuticular con punteaduras gruesas. De acuerdo con la ausencia de las dos apomorfías protorácicas, el género Archosyne Ponomarenko et al., 2014 del Pérmico está excluido de la familia. Como su adscripción taxonómica sigue siendo muy incierta, debe tratarse como Coleoptera incertae sedis. El género Ranis Ponomarenko, 1968 también carece de las características del pronoto que definen a †Ademosynidae. De acuerdo con el propleurón internalizado se transfiere al megadiverso suborden Polyphaga. En su historia evolutiva durante el Mesozoico, †Ademosynidae refleja una tendencia general en Coleoptera, la cual es un refuerzo creciente de la configuración de los escleritos torácicos.
Palabras clave: Coleoptera; †Ademosynidae; reconstrucción paleontológica; evolución temprana
Traducción: Enrique Peñalver (Sociedad Española de Paleontología)
Résumé en Français
Ademosynidae (Insecta : Coleoptera) – un nouveau concept pour un taxon clé des coléoptères et ses affinités phylogénétiques avec les sous-ordres actuels
Ademosynidae est une famille controversée de Coleoptera éteints, découverte pour la première fois dans les dépôts du Trias supérieur d’Australie. Le dernier représentant est connu dans le Jurassique ancien du Kirghizistan. Le matériel-type de la collection du Moscow Paleontological Institute est redécrit ici et interprété phylogénétiquement. Les résultats montrent que la famille telle qu’elle est définie actuellement n’est pas monophylétique, mais qu’elle est potentiellement distribuée au sein de deux des quatre sous-ordres actuels de coléoptères. Nous formalisons un concept plus étroit de la famille en incluant les genres Ademosyne Handlirsch, 1906, Dolichosyne Ponomarenko, 1969, Gnathosyne Ponomarenko, 1969, Cephalosyne Ponomarenko, 1969, Petrosyne Ponomarenko, 1969, et Sphaerosyne Ponomarenko, 1969. Ce groupe monophylétique est caractérisé par deux synapomorphies du prothorax : l’absence d’angles pronotaux antérolatéraux et une marge pronotale antérieure arrondie et protubérante couvrant la partie postérieure de la tête. Les caractères supplémentaires sont de grandes mandibules qui sont fortement saillantes antérieurement, des élytres striés, un angle pronoto-élytral indistinct entraînant un corps fuselé, et une surface cuticulaire avec des ponctuations grossières. Le genre permien Archosyne Ponomarenko et al., 2014 est exclu de la famille car il ne présente pas les deux apomorphies prothoraciques. Comme son placement reste très incertain, il devrait être traité comme Coleoptera incertae sedis. Le genre Ranis Ponomarenko, 1968 ne présente pas les caractères pronotaux qui définissent †Ademosynidae. Il est transféré dans le sous-ordre extrêmement divers, Polyphaga, car il présente un propleuron interne. Pendant leur histoire évolutive au cours du Mésozoïque, les †Ademosynidae reflètent une tendance générale chez les Coleoptera : un renforcement de plus en plus fort de la configuration des sclérites thoraciques.
Mots-clés : Coleoptera ; †Ademosynidae ; reconstitution paléontologique ; évolution
Translator: Antoine Souron
Deutsche Zusammenfassung
Ademosynidae (Insecta: Coleoptera) – ein neues Konzept für ein Schlüsseltaxon der Coleoptera und die phylogenetischen Beziehungen zu den heute noch lebenden Taxa
Die †Ademosynidae sind eine kontroverse ausgestorbene Familie der Coleoptera, die als erstes in australischen obertriassischen Ablagerungen entdeckt wurden. Der letzte bekannte Vertreter ist aus dem frühen Jura von Kirgisistan bekannt. Es wird hier Typenmaterial aus der Sammlung des Instituts für Paläontologie in Moskau neu-beschrieben und phylogenetisch interpretiert. Die Ergebnisse zeigen, dass die Familie wie sie gegenwärtig definiert ist, nicht monophyletisch ist, sondern wahrscheinlich auf zwei oder vier der heute lebenden Subordnungen der Coleoptera aufgeteilt ist. Wir stellen ein neues engeres Konzept der Familie auf, das folgende Gattungen beinhaltet: Ademosyne Handlirsch, 1906, Dolichosyne Ponomarenko, 1969, Gnathosyne Ponomarenko, 1969, Cephalosyne Ponomarenko, 1969, Petrosyne Ponomarenko, 1969 und Sphaerosyne Ponomarenko, 1969. Diese monophyletische Einheit wird durch zwei Synapomorphien im Prothorax, der Abwesenheit von anterolateralen pronotalen Winkeln und durch einen abgerundeten vorstehenden anterioren pronotalen Rand, der den posterioren Teil des Kopfes bedeckt, charakterisiert. Zusätzliche Merkmale sind große Mandibeln, die anterior deutlich vorspringen, gefurchte Elytren, ein undeutlicher pronoto-elytraler Winkel, der in einem stromlinienförmigen Körper resultiert und eine groblöchrige Cuticula-Oberfläche. Da dem permischen Archosyne Ponomarenko et al., 2014 die zwei Apomorphien im Prothorax fehlen wird es aus der Familie herausgenommen. Da die Zuordnungen unklar bleiben, sollte das Taxon als Coleoptera incertae sedis behandelt werden. Bei Ranis Ponomarenko, 1968 fehlen ebenfalls die pronotalen Merkmale der †Ademosynidae. Wegen des internalisierten Propleurons wird es in die megadiverse Subordnung Polyphaga transferiert. Die †Ademosynidae spiegeln in ihrer Evolutionsgeschichte im Mesozoikum den generellen Trend der Coleoptera wieder, die Konfiguration der Thorax-Sklerite zunehmend zu verstärken.
Schlüsselwörter: Coleoptera; †Ademosynidae; paläontologische Rekonstruktion; frühe Evolution
Translator: Eva Gebauer
Arabic
in progress
Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024