

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
 Alexandra A.E. van der Geer. Naturalis Biodiversity Center, P.O. Box 9517, 2300 RA Leiden, the Netherlands. alexandra.vandergeer@naturalis.nl
Alexandra A.E. van der Geer. Naturalis Biodiversity Center, P.O. Box 9517, 2300 RA Leiden, the Netherlands. alexandra.vandergeer@naturalis.nl
Alexandra van der Geer is a palaeontologist, specialised in aspects of evolution and biogeography of extinct mammals on islands. She is also an indologist, specialised in the interface between human material culture and native faunas in South Asia. She has worked at the National and Kapodistrian University of Athens (Greece) since 2004 and currently works at Naturalis Biodiversity Center, Leiden, the Netherlands. Through her research on island mammals she collaborates with geologists, biogeographers, anatomists and archaeologists in attempting to reconstruct the evolution, adaptation and extinction of insular endemics globally and to investigate the role of humans. She is currently expanding her research focus to include extant insular mammals with emphasis on South East Asia and the Pacific.
FIGURE 1. Schematic map of the Mediterranean with the island of Crete indicated with an oblong and showing the fossil sites with antler remains of the endemic deer dwarf Candiacervus mentioned in this study. 1 = Melidoni, 2 = Liko, 3 = Peristeri 2, 4 = Kaló Chorafi, 5 = Sifanos, 6 = Kharoum(b)es, 7 = Gerani 2, 8 = Gerani 4, 9 = Gerani 5, 10 = Sourida, 11 = Bate Cave, 12 = Mavro Muri 3 and 4, 13 = Simonelli Cave, 14 = Gumbes, 15 = Grida Avlaki, 16 = Panajia 2. Not indicated here are Rethymnon fissure (just west of Rethymnon) and Gerani 1 and 6 (near Gerani 5).
FIGURE 2. Composite mount of an adult stag of a dwarf form (withers height c. 50 cm) with of Candiacervus with typical ropalophorus-type of antler. Scale bar equals 30 cm.

FIGURE 3. Three antlers of Candiacervus ropalophorus. 1, Cast of the type antler (RGM 438460). 2, Curved variety, right shed antler (AMPG[V] 560). 3, Curved variety with clear bludgeon-shaped distal end (AMPG[V] 2133).

FIGURE 4. 3D scan of the type antler (RGM 438460) of C. ropalophorus.

FIGURE 5. 3D scan of the curved antler variety (AMPG[V] 560) of C. ropalophorus.

FIGURE 6. Candiaverus devosi sp. nov. 1, Holotype AMPG(V) 1735, skull with preserved proximal left antler, latero-frontal view. 2, Paratype AMPG(V) 1733, right shed antler. 3, Artificial combination of the holotype and paratype to visually reconstruct the original configuration.

FIGURE 7. 3D scan of the paratype of C. devosi sp. nov.

FIGURE 8. Candiacervus listeri sp. nov. 1, Holotype AMPG(V) 1734, skull with preserved proximal antlers, lateral view. 2, Paratype AMPG(V) 1726, partial skull with attached left and right antlers missing the distal tips, dorso-frontal view. 3, Referred right shed antler AMPG(V) 2106 (Kalo Chorafi?), showing the posterior rudimentary palmation. 4, Referred left shed antler AMPG(V) 2115 (Rethymnon area) with rudimentary back tine.

FIGURE 9. 3D scan of the right antler of the paratype of C. listeri sp. nov. (AMPG[V] 1726).
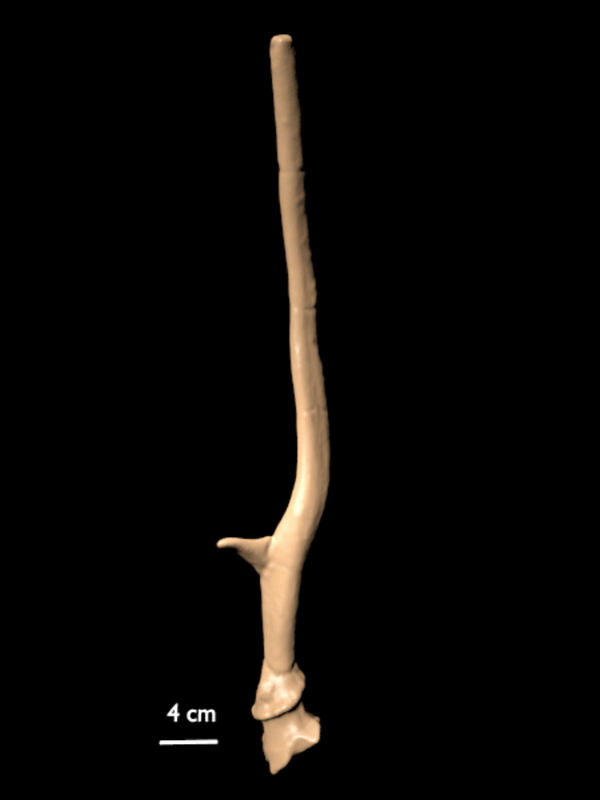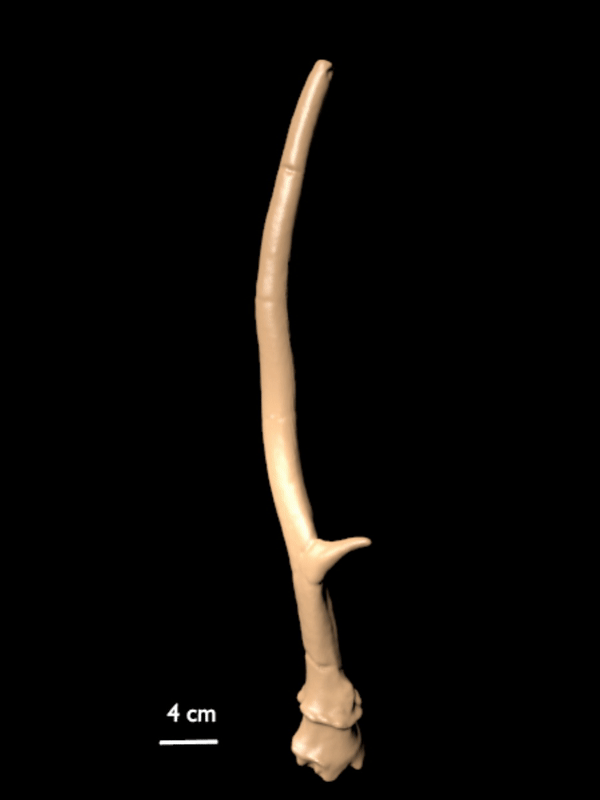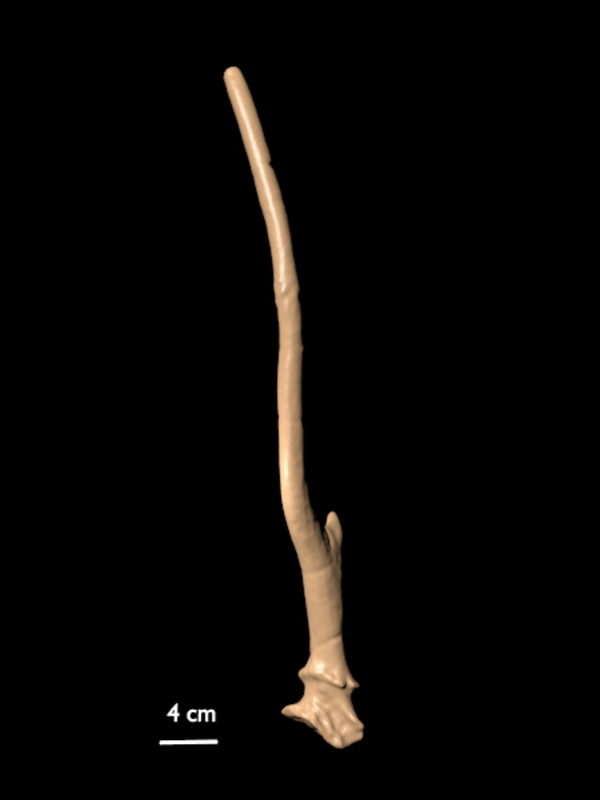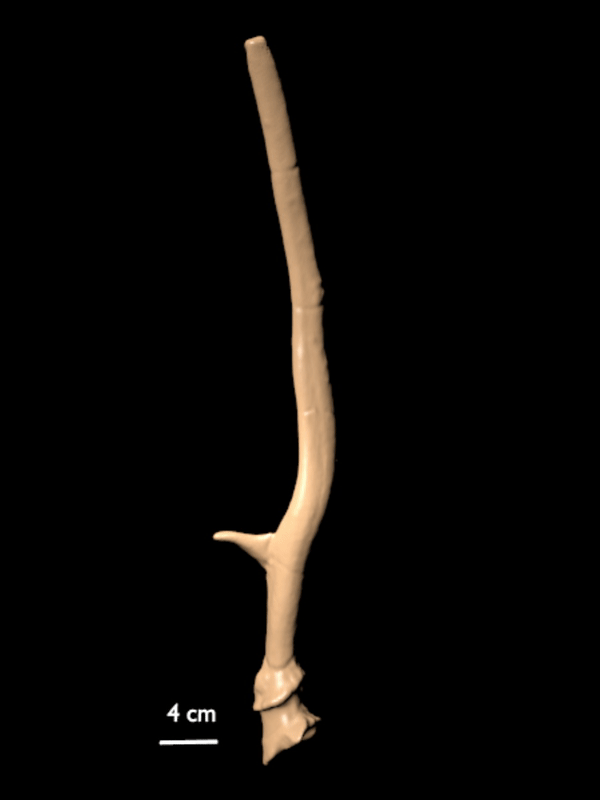
FIGURE 10. 3D scan of a right shed antler of C. listeri sp. nov. (AMPG[V] Li-2593), type antler of de Vos' (1984) type 2.
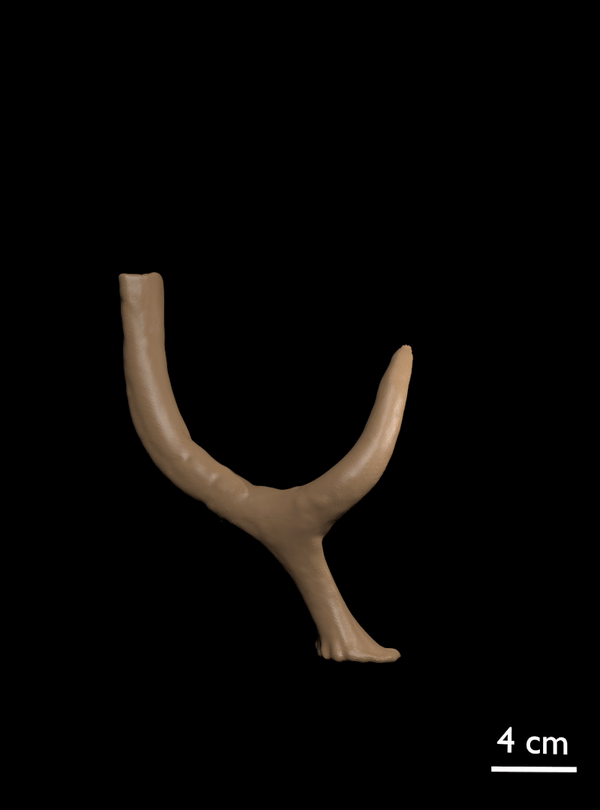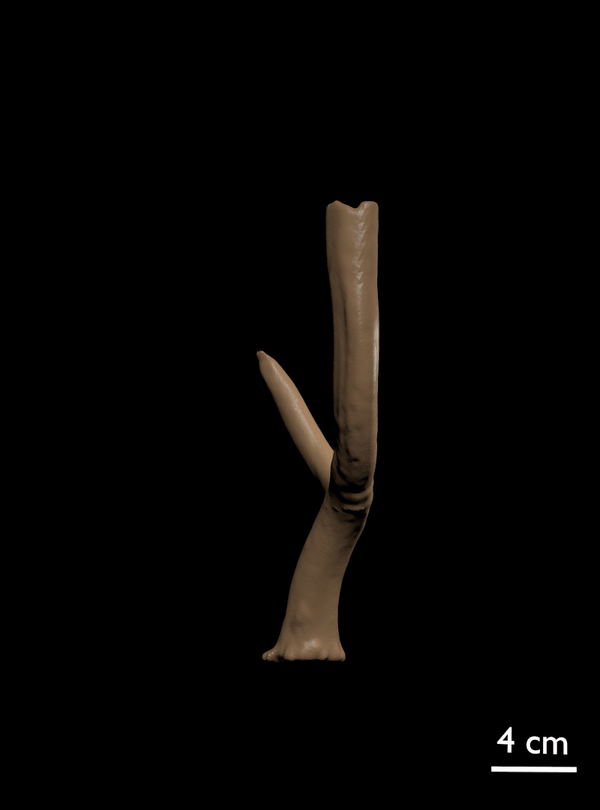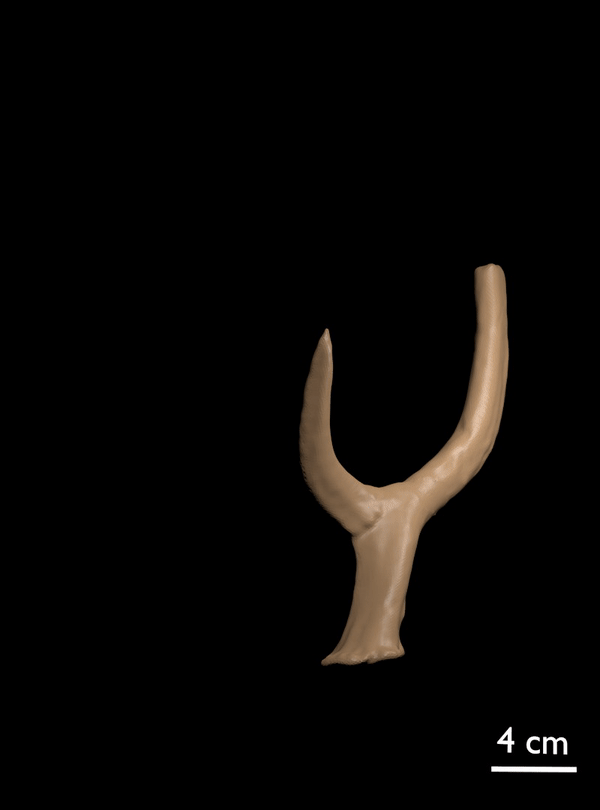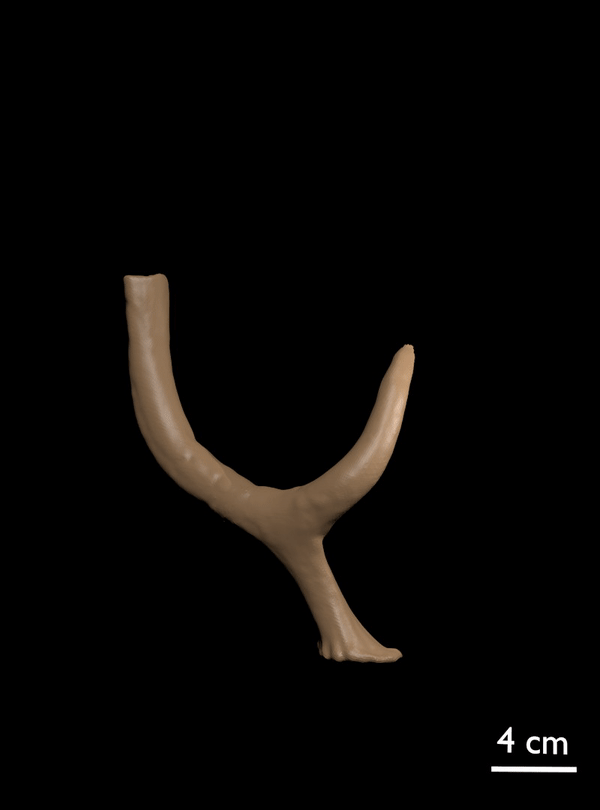
FIGURE 11. 3D scan of referred antler of C. listeri with rudimentary posterior palmation (AMPG[V] 2106).
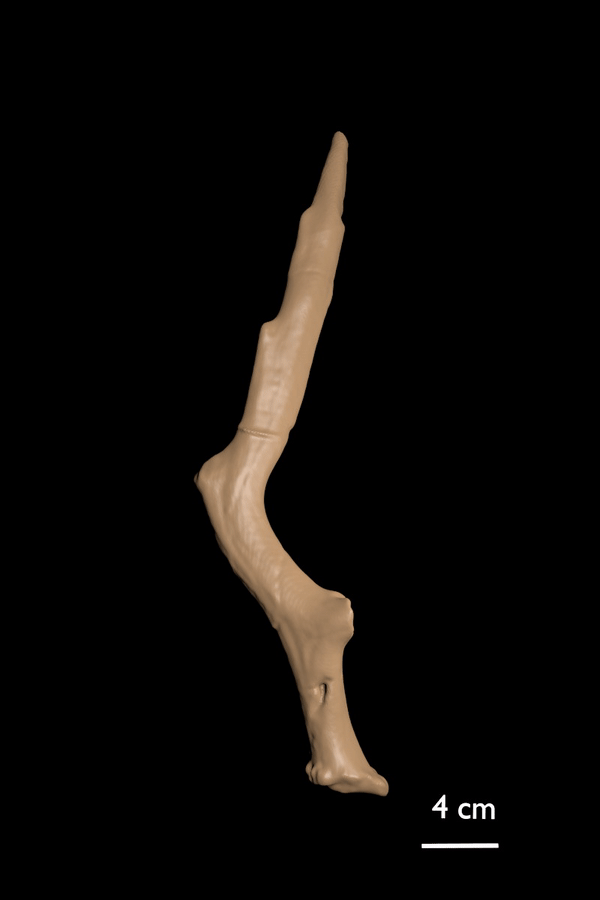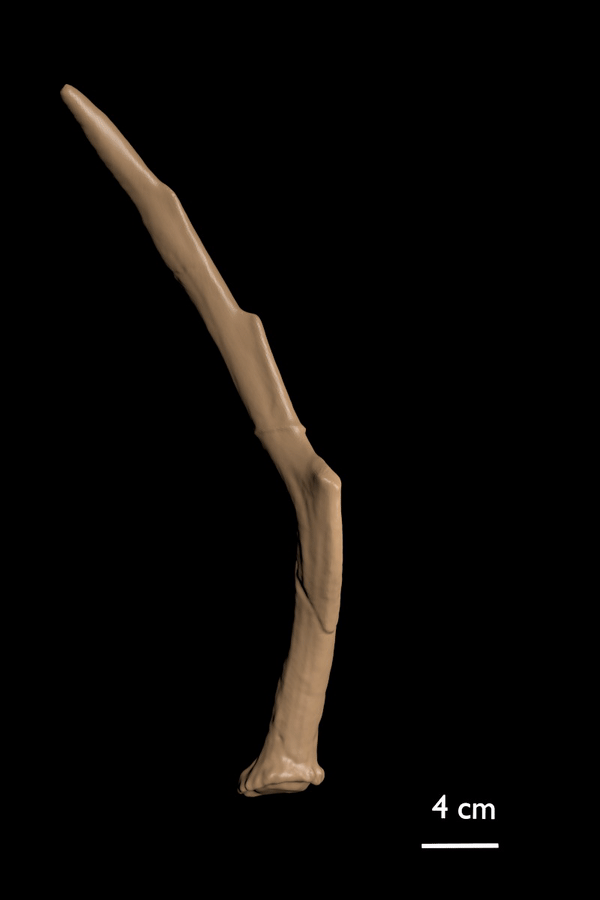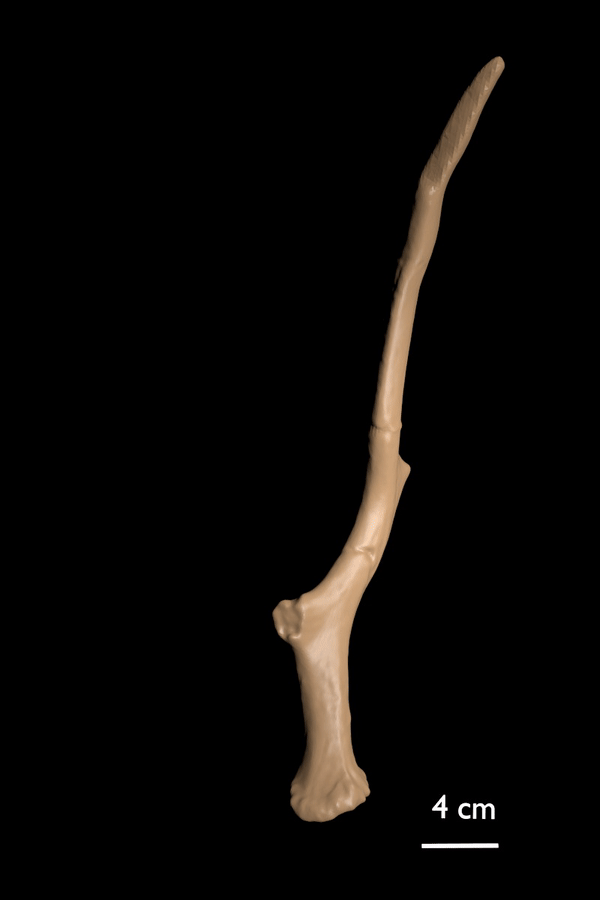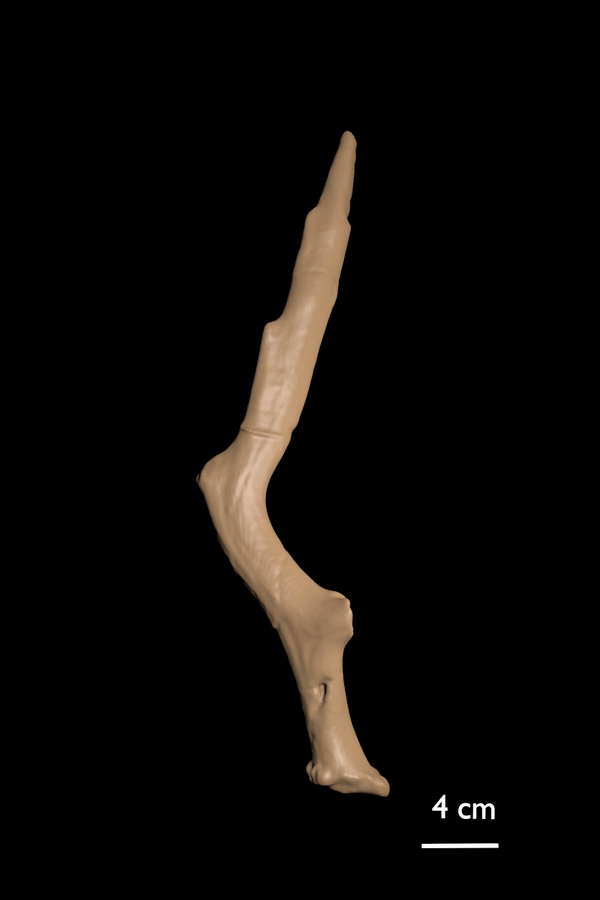
FIGURE 12. Candiacervus reumeri sp. nov. 1, Holotype AMPG(V) 1736, skull with attached proximal antlers, lateral view. 2, Paratype RGM 442702, Left shed antler. 3, Referred right shed antler AMPG(V) 562 (Liko OD), showing a rudimentary second tine. 4, Referred right shed antler AMPG(V) 2105 (Peristeri 2).
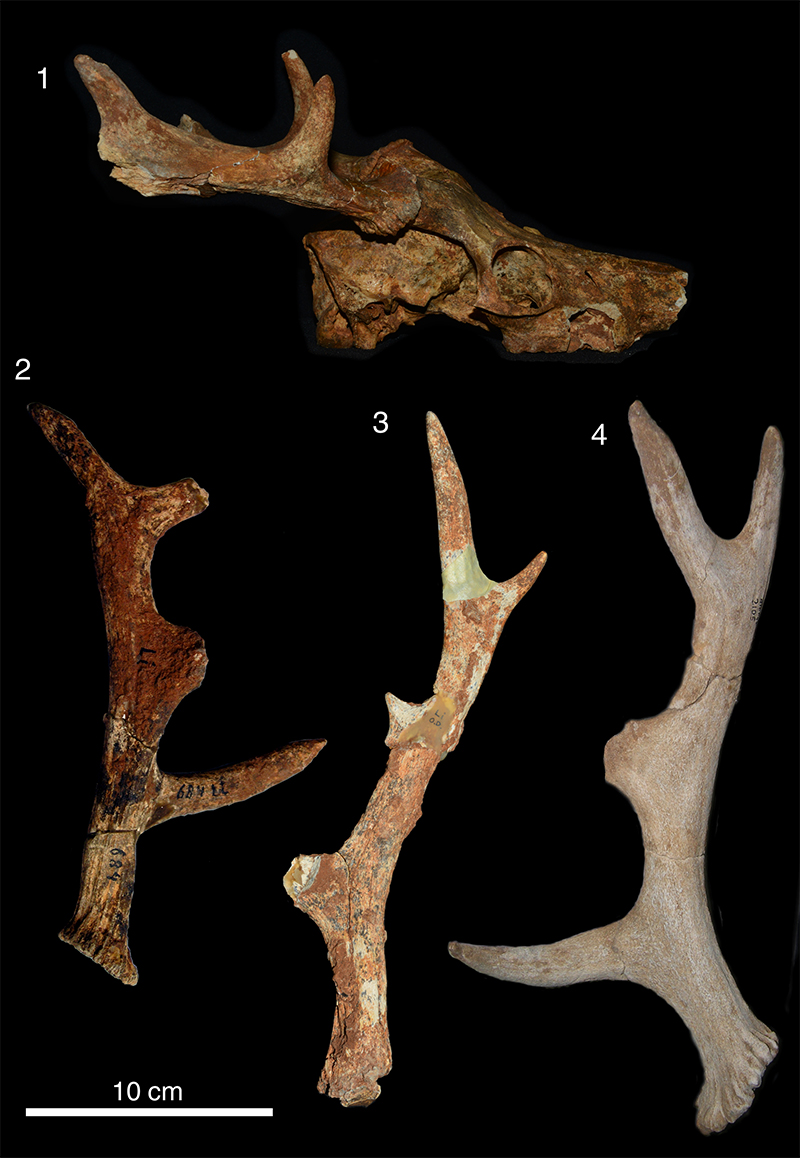
FIGURE 13. 3D scan of referred right shed antler of C. reumeri sp. nov. (AMPG[V] 562).
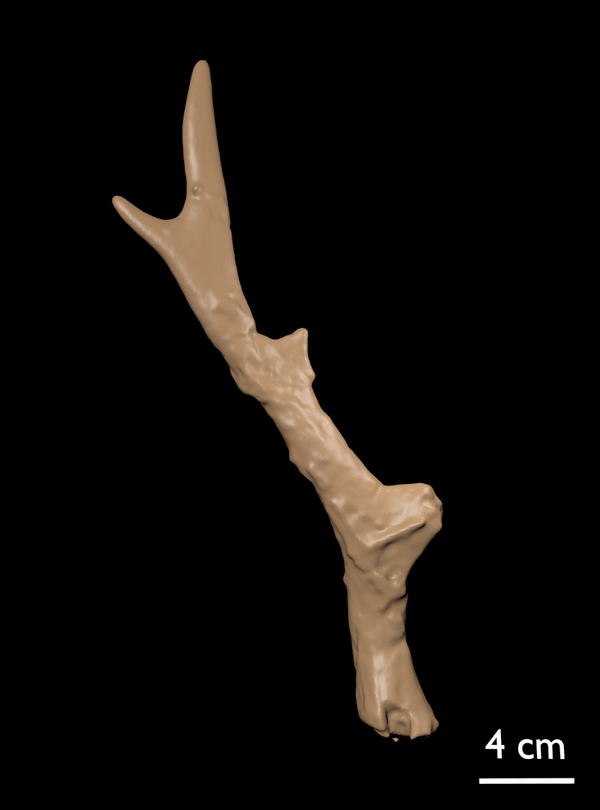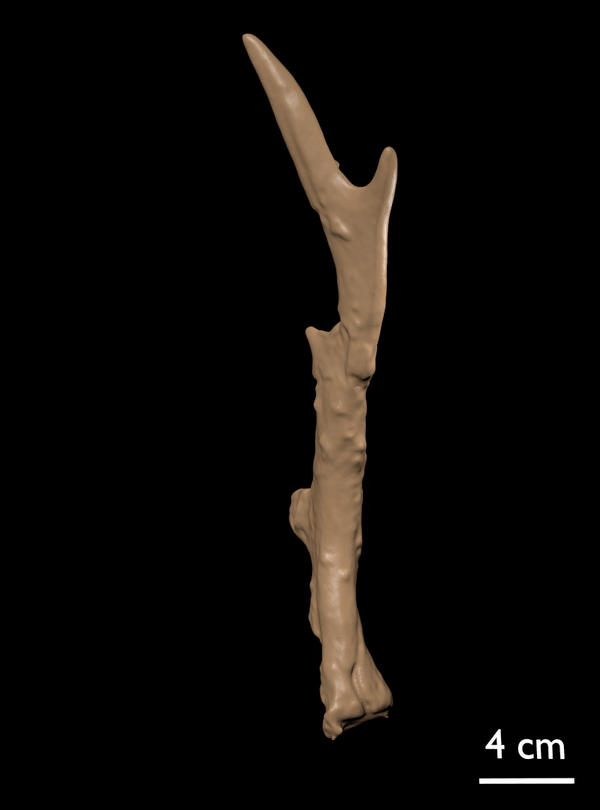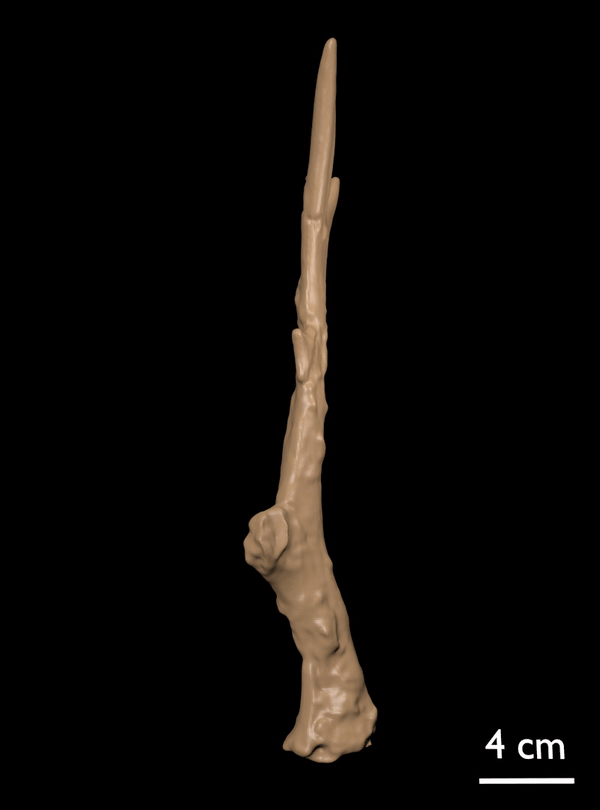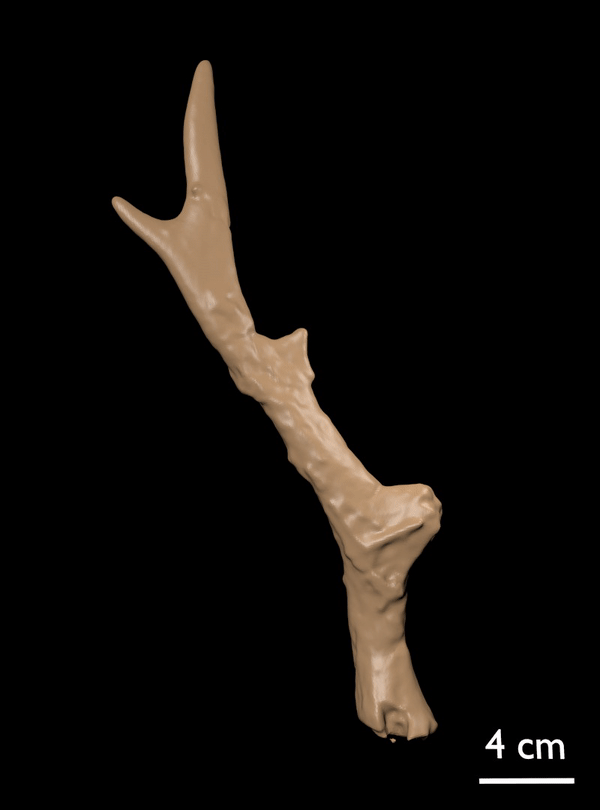
FIGURE 14. Candiacervus cf. reumeri. 1, Referred left shed antler AMPG(V) 561 (Liko OD), showing the rudimentary sub-basal tine. 2, Referred right shed antler AMPG(V) 2138 (Kalo Chorafi).

FIGURE 15. Schematic representation of the evolution of two main types of antlers of Candiacervus. Several antler types can be distinguished, but these can be interpreted as evolutionary derivations (stages) of two basic bauplans. These derivations likely represent contemporaneous, geographically separated species or ecomorphs. The connecting lines only show the order of changes needed to evolve another antler morphology. The evolution as presented here is merely an illustration of how the various antler shapes of Candiacervus may have evolved, simply by adding and reduction of tines and lengthening of the last segment. 1, C. devosi stage; 2, C. listeri stage; 3, C. ropalophorus stage; 4, C. reumeri stage.

APPENDIX 1.
Antler burr diameters and minimal length (measured in a straight line) for Candiacervus material included in this study. DAP burr = maximal anterior-posterior width of the burr, DT burr = maximal transverse width of the burr, L antler = total antler length in a straight line (L only measured in complete specimens). All measurements are in mm.
| Liko B variety sp. IIa (de Vos 1984) |
Collection | Number | DAP burr | DT burr | L antler |
| Liko B | AMPG(V) | 1721 | 45 | 42.4 | |
| Liko B | AMPG(V) | 1734 | 37 | 42 | |
| Liko B | AMPG(V) | Li-635 | 45.8 | 45.2 | |
| Liko B | AMPG(V) | Li-636 | 47.2 | 48 | |
| Liko B | AMPG(V) | Li-637 | 41.6 | 39.4 | |
| Liko B | AMPG(V) | Li-638 | 52.6 | 50.4 | |
| Liko B | AMPG(V) | Li-639 | 37.6 | 35.6 | |
| Liko B | AMPG(V) | Li-640 | 38.2 | 37.4 | |
| Liko B | AMPG(V) | Li-642 | 33.8 | 29.8 | |
| Liko B | AMPG(V) | Li-644 | 36.6 | 34.4 | |
| Liko B | AMPG(V) | Li-645 | 37.1 | 35.6 | |
| Liko B | AMPG(V) | Li-646 | 34.6 | 31.4 | |
| Liko B | AMPG(V) | Li-647 | 41.6 | 37.8 | |
| Liko B | AMPG(V) | Li-648 | 33.8 | ||
| Liko B | AMPG(V) | Li-649 | 41.5 | ||
| Liko B | AMPG(V) | Li-650 | 36.2 | 34.1 | |
| Liko B | AMPG(V) | Li-651 | 42.8 | 36.2 | |
| Liko B | AMPG(V) | Li-652 | 37.2 | 33.1 | |
| Liko B | AMPG(V) | Li-653 | 40.2 | 40 | |
| Liko B | AMPG(V) | Li-654 | 48.8 | 46.1 | |
| Liko B | AMPG(V) | Li-655 | 35.6 | 34.8 | 166.9 |
| Liko B | AMPG(V) | Li-656 | 35 | 38.2 | |
| Liko B | AMPG(V) | Li-658 | 38 | 32.6 | |
| Liko B | AMPG(V) | Li-659 | 47.2 | ||
| Liko B | AMPG(V) | Li-660 | 61.4 | 50.4 | |
| Liko B | AMPG(V) | Li-661 | 36.2 | 33.4 | |
| Liko B | AMPG(V) | Li-662 | 47 | 43.6 | |
| Liko B | AMPG(V) | Li-663 | 24.1 | ||
| Liko B | AMPG(V) | Li-664 | 35.1 | ||
| Liko B | AMPG(V) | Li-665 | 36.2 | 39.5 | |
| Liko B | AMPG(V) | Li-666 | 37.1 | ||
| Liko B | AMPG(V) | Li-667 | 46.2 | 41.7 | |
| Liko B | AMPG(V) | Li-668 | 46.7 | 43.2 | |
| Liko B | AMPG(V) | Li-670 | 37.1 | 31.6 | |
| Liko B | AMPG(V) | Li-671 | 49.4 | 44.2 | |
| Liko B | AMPG(V) | Li-672 | 30.6 | 28.2 | |
| Liko B | AMPG(V) | Li-675 | 43.2 | 43 | |
| Liko B | AMPG(V) | Li-676 | 32.4 | 30.1 | |
| Liko B | AMPG(V) | Li-678 | 35.3 | 31.8 | |
| Liko B | AMPG(V) | Li-2518 | 32.8 | 31.8 | |
| Liko B | AMPG(V) | Li-2519 | 33.8 | 29 | |
| Liko B | AMPG(V) | Li-2520 | 39.9 | 37.8 | |
| Liko B | AMPG(V) | Li-2521 | 30.7 | 29.8 | |
| Liko B | AMPG(V) | Li-2522 | 35.6 | ||
| Liko B | AMPG(V) | Li-2523 | 40.8 | 36.8 | |
| Liko B | AMPG(V) | Li-2525 | 37.1 | 36.2 | |
| Liko B | AMPG(V) | Li-2526 | 30.8 | 29.6 | |
| Liko B | AMPG(V) | Li-2528 | 35.8 | 31.4 | |
| Liko B | AMPG(V) | Li-2529 | 35.2 | ||
| Liko B | AMPG(V) | Li-2530 | 38.7 | 34.1 | |
| Liko B | AMPG(V) | Li-2531 | 43.4 | 40 | |
| Liko B | AMPG(V) | Li-2532 | 49.8 | 40.4 | |
| Liko B | AMPG(V) | Li-2533 | 41.7 | 35.8 | |
| Liko B | AMPG(V) | Li-2535 | 41.6 | ||
| Liko B | AMPG(V) | Li-2536 | 38.8 | ||
| Liko B | AMPG(V) | Li-2537 | 39.8 | 35.2 | |
| Liko B | AMPG(V) | Li-2538 | 41.6 | ||
| Liko B | AMPG(V) | Li-2539 | 45 | 39.5 | |
| Liko B | AMPG(V) | Li-2540 | 30.6 | ||
| Liko B | AMPG(V) | Li-2541 | 42 | ||
| Liko B | AMPG(V) | Li-2543 | 33.6 | 30 | |
| Liko B | AMPG(V) | Li-2544 | 41.1 | ||
| Liko B | AMPG(V) | Li-2546 | 46.2 | ||
| Liko B | AMPG(V) | Li-2547 | 42.4 | 42.4 | |
| Liko B | AMPG(V) | Li-2549 | 39 | 36.2 | |
| Liko B | AMPG(V) | Li-2550 | 41.4 | ||
| Liko B | AMPG(V) | Li-2551 | 43.2 | 40.8 | |
| Liko B | AMPG(V) | Li-2553 | 49.8 | 46.2 | |
| Liko B | AMPG(V) | Li-2554 | 27.8 | 25.2 | |
| Liko B | AMPG(V) | Li-2555 | 29.8 | 27.9 | |
| Liko B | AMPG(V) | Li-2556 | 35.6 | 36.9 | |
| Liko B | AMPG(V) | Li-2557 | 27.4 | 32.2 | |
| Liko B | AMPG(V) | Li-2558 | 39.2 | 32.6 | |
| Liko B | AMPG(V) | Li-2559 | 32.6 | 30 | |
| Liko B | AMPG(V) | Li-2560 | 36.9 | 33.2 | |
| Liko B | AMPG(V) | Li-2561 | 40 | ||
| Liko B | AMPG(V) | Li-2563 | 41.4 | 37.6 | |
| Liko B | AMPG(V) | Li-2566 | 42.2 | 37.4 | |
| Liko B | AMPG(V) | Li-2567 | 42.2 | 38.8 | |
| Liko B | AMPG(V) | Li-2569 | 53.6 | 47.1 | |
| Liko B | AMPG(V) | Li-2571 | 51.7 | 40.2 | |
| Liko B | AMPG(V) | Li-2572 | 43 | 41.2 | |
| Liko B | AMPG(V) | Li-2573 | 40 | 38.2 | |
| Liko B | AMPG(V) | Li-2574 | 46.2 | 41.2 | |
| Liko B | AMPG(V) | Li-2575 | 49.6 | 49.4 | |
| Liko B | AMPG(V) | Li-2576 | 42.1 | 43 | |
| Liko B | AMPG(V) | Li-2583 | 38.2 | 37.4 | |
| Liko B | AMPG(V) | Li-2584 | 34.2 | 30.9 | |
| Liko B | AMPG(V) | Li-2586 | 39.4 | 37.2 | |
| Liko B | AMPG(V) | Li-2587 | 44.4 | 41.4 | |
| Liko B | AMPG(V) | Li-2588 | 47.6 | 40.4 | |
| Liko B | AMPG(V) | Li-2589 | 40.6 | 43.8 | |
| Liko B | AMPG(V) | Li-2591 | 38.2 | 30 | |
| Liko B | AMPG(V) | Li-2592 | 36.6 | 34.6 | |
| Liko B | AMPG(V) | Li-2593 | 41.6 | 41.8 | |
| Liko B | RGM | 442696 | 34 | ||
| Liko B | RGM | Li-2585 | 45.2 | 44.4 | |
| Liko B spikes for comparison | |||||
| Liko B | AMPG(V) | Li-680 | 16.6 | 15.6 | 118.8 |
| Liko B | AMPG(V) | Li-681 | 24.6 | 20.6 | 186 |
| Liko B | AMPG(V) | Li-682 | 34.6 | 29.8 | 180 |
| Liko B | AMPG(V) | Li-683 | 20 | 20.3 | |
| Liko B | AMPG(V) | Li-2610 | 16.1 | 18.6 | |
| Liko B | AMPG(V) | Li-2611 | 15.8 | 13.6 | |
| Liko B | AMPG(V) | Li-2612 | 18.2 | 18.1 | |
| Liko B | AMPG(V) | Li-2613 | 14.1 | 11.4 | |
| Liko B | AMPG(V) | Li-2614 | 17.2 | 15.2 | |
| Liko B | AMPG(V) | Li-2615 | 17.2 | 12.4 | 85 |
| Liko B | RGM | Li-2616 | 16.4 | 14.4 | 100 |
| Liko B | RGM | Li-2617 | 12.4 | 11.8 | 60 |
| Liko B | RGM | Li-2619 | 16.4 | 16 | |
| Liko B | RGM | Li-2620 | 12.6 | 9.1 | |
| Liko B | RGM | Li-2622 | 14.8 | 12.4 | |
| Liko B | RGM | Li-2623 | 15.1 | 9.4 | 92.8 |
| Liko B | RGM | Li-2625 | 13.6 | 12.2 | |
| Liko B | RGM | Li-2626 | 18.6 | 15.9 | |
| Liko B | RGM | Li-2627 | 20.4 | 20.2 | |
| Liko B | RGM | Li-2628 | 19.8 | 18.9 | |
| Liko B | RGM | Li-2630 | 29.4 | 23.6 | |
| Liko B | RGM | Li-2632 | 24.8 | 23.4 | |
| Liko B | RGM | Li-2633 | 25.4 | 24.8 | |
| Liko B | RGM | Li-2634 | 35.6 | 30.8 | |
| iko B | RGM | Li-2635 | 25 | 22.8 | |
| Liko B | RGM | Li-2636 | 42.4 | 37.2 | |
| Liko B variety sp. IIc of De Vos (1984) |
|||||
| Liko B | RGM | 442702 | 37 | 37.2 | 213 |
| Liko B | RGM | Li-759 | 39 | 40 | |
| Liko B | RGM | Li-759 | 40 | 40 | |
| Liko B variety sp. IIb of De Vos (1984) |
|||||
| Liko B | RGM | Li-669 | 40.1 | ||
| Liko B | RGM | Li-685 | 43 | 37.4 | |
| Liko B | RGM | Li-686 | 45 | 42 | |
| Liko B | RGM | Li-687 | 49 | 51 | |
| Liko B | RGM | Li-688 | 30.4 | 26.9 | |
| Liko B | RGM | Li-2595 | 37.2 | 35.8 | |
| Liko B | RGM | Li-2608 | 35 | 23 | |
| Liko B prox fragments | |||||
| Liko B | AMPG(V) | Li-689 | 40.2 | 41 | |
| Liko B | AMPG(V) | Li-690 | 37.1 | 34.5 | |
| Liko B | AMPG(V) | Li-691 | 33.3 | 28.2 | |
| Liko B | AMPG(V) | Li-692 | 38.1 | 32.6 | |
| Liko B | AMPG(V) | Li-693 | 34.3 | 29.6 | |
| Liko B | AMPG(V) | Li-694 | 41.6 | ||
| Liko B | AMPG(V) | Li-695 | 38.9 | 34.1 | |
| Liko B | AMPG(V) | Li-696 | 35.8 | 33.4 | |
| Liko B | AMPG(V) | Li-697 | 37.5 | 36 | |
| Liko B | AMPG(V) | Li-698 | 34.6 | ||
| Liko B | AMPG(V) | Li-699 | 43 | 43.1 | |
| Liko B | AMPG(V) | Li-700 | 48.6 | 46.8 | |
| Liko B | AMPG(V) | Li-701 | 45.2 | 44.3 | |
| Liko B | AMPG(V) | Li-702 | 50.7 | 47.4 | |
| Liko B | AMPG(V) | Li-703 | 39.2 | 35.3 | |
| Liko B | AMPG(V) | Li-704 | 38.2 | ||
| Liko B | AMPG(V) | Li-705 | 42.1 | 42 | |
| Liko B | AMPG(V) | Li-706 | 40.6 | 36.6 | |
| Liko B | AMPG(V) | Li-707 | 35.3 | 29.7 | |
| Liko B | AMPG(V) | Li-708 | 35.2 | 35.6 | |
| Liko B | AMPG(V) | Li-709 | 35 | 36.6 | |
| Liko B | AMPG(V) | Li-710 | 39 | 37.2 | |
| Liko B | AMPG(V) | Li-711 | 39 | 37.7 | |
| Liko B | AMPG(V) | Li-712 | 50.7 | 50.2 | |
| Liko B | AMPG(V) | Li-713 | 35 | 36.3 | |
| Gerani 4 | RGM | Ge4-2870 | 47.9 | 43.2 | 77 |
| Gerani 4 | RGM | Ge4-2871 | 33.4 | 36.7 | |
| Gerani 4 | RGM | Ge4-2872 | 42.4 | 33.5 | |
| Gerani 4 | RGM | Ge4-2876 | 37 | 32.4 | |
| Gerani 4 | RGM | Ge4-2877 | 33.9 | 30.8 | |
| Gerani 4 | RGM | Ge4-2883 | 44 | 42.2 | |
| Gerani 4 | RGM | Ge4-2884 | 44.5 | 35.8 | |
| Gerani 4 | RGM | Ge4-2885 | 36.6 | 31.7 | |
| Gerani 4 | RGM | Ge4-2886 | 45 | 41.2 | |
| Gerani 4 | RGM | Ge4-2887 | 40 | ||
| Gerani 4 | RGM | Ge4-2888 | 37.1 | 34.8 | |
| Gerani 4 | RGM | Ge4-2889 | 37.1 | 34.9 | |
| Gerani 4 | RGM | Ge4-2890 | 42.4 | 38.2 | |
| Gerani 4 | RGM | Ge4-2891 | 30.7 | 30.5 | |
| Gerani 4 | RGM | Ge4-2892 | 47 | 43.2 | |
| Gerani 4 | RGM | Ge4-2896 | 34.2 | 30.3 | |
| Gerani 4 | RGM | Ge4-2897 | 37.2 | 32.4 | |
| Gerani 4 | RGM | Ge4-2898 | 38.5 | 34.7 | |
| Gerani 4 | RGM | Ge4-2902 | 32.6 | 30.6 | |
| Gerani 4 | RGM | Ge4-2904 | 46 | 38.1 | |
| Gerani 4 | RGM | Ge4-2905 | 26.7 | ||
| Gerani 4 | RGM | Ge4-2906 | 44.2 | 38.9 | |
| Gerani 4 | RGM | Ge4-2907 | 38.9 | 33.9 | |
| Gerani 4 | RGM | Ge4-2908 | 33.8 | 28.3 | |
| Gerani 4 | RGM | Ge4-2910 | 36.4 | 35.4 | |
| Gerani 4 | RGM | Ge4-2911 | 36.2 | ||
| Gerani 4 | RGM | Ge4-2912 | 30 | ||
| Gerani 4 | RGM | Ge4-2913 | 42.8 | 41.5 | |
| Gerani 4 | RGM | Ge4-2914 | 39.8 | 41 | |
| Gerani 4 | RGM | Ge4-2915 | 40.5 | 38.8 | |
| Gerani 4 | RGM | Ge4-2916 | 38.6 | ||
| Gerani 4 | RGM | Ge4-2917 | 29 | 26.8 | |
| Gerani 4 | RGM | Ge4-2918 | 34.4 | 31.2 | |
| Gerani 4 | RGM | Ge4-2919 | 35 | ||
| Gerani 4 | RGM | Ge4-2920 | 34 | 31.4 | |
| Gerani 4 | RGM | Ge4-2921 | 39.8 | 38.7 | |
| Gerani 4 | RGM | Ge4-2922 | 41.7 | 43 | |
| Gerani 4 | RGM | Ge4-2923 | 42 | 37.4 | |
| Gerani 4 | RGM | Ge4-2924 | 33.2 | 30.9 | |
| Gerani 4 | RGM | Ge4-2925 | 33.9 | ||
| Gerani 4 | RGM | Ge4-2926 | 51.2 | 47.9 | |
| Gerani 4, spikes, just for comparison |
|||||
| Gerani 4 | RGM | Ge4-3514 | 14.6 | 16.2 | |
| Gerani 4 | RGM | Ge4-3515 | 14.4 | 11 | 59.6 |
| Gerani 4 | RGM | Ge4-3517 | 16.1 | 14.8 | |
| Various sites | |||||
| Gerani 1 | NHMC | 69.95 | 38.7 | 36.4 | |
| Gerani 1 | NHMC | 69.94 | 38.8 | 32 | |
| Gerani | AMPG(V) | 560 | 38 | 37 | |
| Gerani | AMPG(V) | GE764 | 47 | 39 | |
| Gerani | AMPG(V) | 2114 | 43 | 37 | |
| Grida Avlaki | NHMC | G.A.68 | 38 | 33 | |
| Gumbus B | AMPG(V) | 2147 | 41 | 36 | |
| Gumbus B | AMPG(V) | 2147 | 36 | 33 | |
| Gumbes C | AMPG(V) | 15 | 31.2 | ||
| Gumbes C | AMPG(V) | 16 | 35.5 | 30.3 | |
| Gumbes C | AMPG(V) | 17 | 31.4 | 27.4 | |
| Kalo Chorafi | NHMC | 109.63 | 40 | 35 | |
| Kalo Chorafi | NHMC | 110.63 | 41 | 40 | |
| Kalo Chorafi | AMPG(V) | 2138 | 44 | 39 | 269 |
| Kalo Chorafi | AMPG(V) | 2106 | 41 | 37 | |
| Liko C | AMPG(V) | 2101 | 34 | 31 | |
| Liko D | AMPG(V) | 2110 | 49 | ||
| Liko D | AMPG(V) | 2103 | 33 | 29 | |
| Liko D | AMPG(V) | 2153 | 41 | 39 | |
| Liko OD | AMPG(V) | 2104 | 37 | 43 | |
| Liko OD | AMPG(V) | 1738 | 44 | 36 | |
| Mavro Muri 4c | AMPG(V) | 498 | 218 | ||
| Mavro Muri 4c | AMPG(V) | 499 | 180 | ||
| Mavro Muri 4c | AMPG(V) | 500 | 28.7 | 25.4 | |
| Mavro Muri 4c | AMPG(V) | 501 | 38.6 | 33.9 | |
| Mavro Muri 4c | AMPG(V) | 502 | 36.1 | 31.6 | |
| Mavro Muri 4c | AMPG(V) | 503 | 35.6 | 35.4 | |
| Mavro Muri 4c | AMPG(V) | 504 | 35 | ||
| Mavro Muri 4c | AMPG(V) | 505 | 43 | 38.5 | |
| Peristeres 2 | AMPG(V) | 2137 | 43 | 36 | |
| Peristeres 2 | AMPG(V) | 2136 | 43 | 35 | |
| Rethymnon area | AMPG(V) | 2118 | 41 | 34 | |
| Rethymnon area | AMPG(V) | 2115 | 50 | 45 | |
| Rethymnon area | AMPG(V) | 2122 | 45 | 35 | |
| Simonelli | AMPG(V) | 126 | 39 | ||
| Largest specimens | |||||
| Sifanos | NHMC | F538/30 | 56 | 50 | |
| Grida Avlaki | NHMC | F538/53 | 56 | 54 | |
| Mavro Muri 4 | NHMC | MV4c.670 | 56.4 | 58.6 | |
| Melidoni | NHMC | skull M | 65.2 | 61.1 |
APPENDIX 2.
Scatter plot of burr diameters of Candiacervus antlers from various localities and size classes.
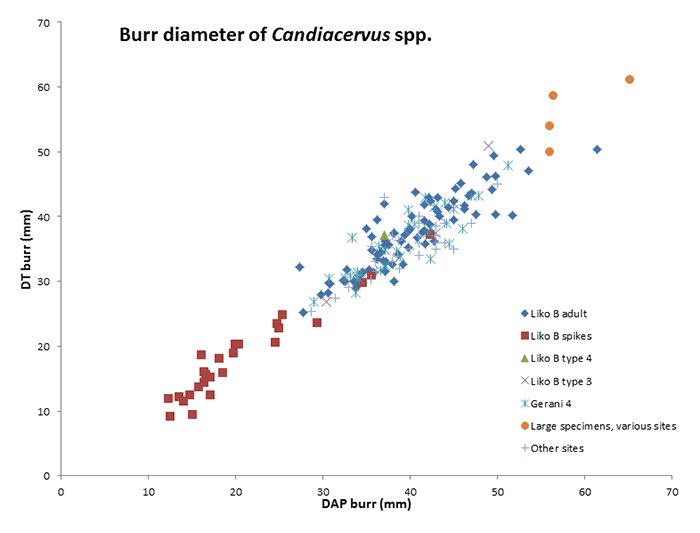
Uniformity in variety: Antler morphology and evolution in a predator-free environment
Plain Language Abstract
The Late Pleistocene mammal fauna of Crete was impoverished, as typical for oceanic islands, and consisted only of deer, dwarf elephants, an otter, a shrew and giant mice. Dwarf deer were the dominant herbivorous species. Here, I describe the adult antlers of this deer. Antler variety appears to be remarkably large, yet a few concise morphological groups without intermediate forms can be recognized, likely representing separate species. Three new species are described here, differing in antler and skull morphology, and the diagnosis of existing species is emended based on new material. Antler variation can be explained by two evolutionary trends: showiness versus a classic fighting type. Divergence is driven and accelerated by intra-specific competition among males. The classic type is best explained as a result of allometric down-scaling during dwarfism. The display type is best explained as a result of restructuring of antler architecture (simplification and extreme elongation of the main beam). In environments without mammalian predators, deer have the potential to evolve antler shapes and sizes and behavioural changes unknown on the mainland.
Resumen en Español
Uniformidad en la variedad: morfología y evolución de la cornamenta en un ambiente libre de depredadores
La fauna de mamíferos de Creta en el Pleistoceno tardío estaba empobrecida, como es típico en las islas oceánicas, y consistía únicamente en venados, elefantes enanos, una nutria, una musaraña y ratones gigantes. Los ciervos enanos (Candiacervus spp.) constituían las especies herbívoras endémicas dominantes. Se describe la morfología de la cornamenta adulta de este ciervo. La variedad de astas parece ser notablemente grande, aunque se pueden reconocer algunos grupos morfológicos concretos sin formas intermedias, los cuales probablemente representan especies separadas. La variedad de asta no es un producto de la variación aleatoria inducida por la escasa presión ecológica de un ambiente libre de depredadores. Aquí se describen tres nuevas especies (Candiacervus spp. nov.), que difieren en la morfología de la cornamenta y el cráneo, y se enmiendan las diagnosis de las especies actuales (C. ropalophorus, C. rethymnensis) a partir de material nuevo. La variación de la cornamenta puede explicarse por dos tendencias evolutivas: ostentación vs. un tipo de lucha clásica. La divergencia es impulsada y acelerada por la competencia intraespecífica entre los machos. El tipo clásico se explica mejor como resultado de la reducción alométrica durante el enanismo. El tipo referido a la exhibición (display) se explica mejor como resultado de la reestructuración del bauplan del asta (simplificación y alargamiento extremo del haz principal). Bajo escenarios libres de depredadores, los ciervos tienen el potencial de desarrollar morfologías de asta y cambios de comportamiento desconocidos en el medio terrestre continental.
Palabras clave: Candiacervus; venado endémico; nuevas especies; radiación adaptativa
Traducción: Enrique Peñalver (Sociedad Española de Paleontología)
Résumé en Français
L’uniformité dans la variété : morphologie des bois et évolution dans un environnement sans prédateurs
La faune de mammifères du Pléistocène récent de Crète était appauvrie, comme c’est typiquement le cas des îles océaniques, et comportait exclusivement des cerfs, des éléphants nains, une loutre, une musaraigne, et des souris géantes. Des cerfs nains (Candiacervus spp.) étaient les espèces herbivores endémiques dominantes. Dans cet article, je décris la morphologie adulte des bois de ce cerf. La variété des bois apparait remarquablement élevée, et pourtant, quelques groupes morphologiques sans formes intermédiaires peuvent être reconnus et représentent probablement des espèces différentes. La variété des bois n’est pas le produit d’une variation aléatoire provoquée par le relâchement écologique dans un environnement dénué de prédateurs. Trois nouvelles espèces sont décrites dans cet article (Candiacervus spp. nov.), discriminées par la morphologie du crâne et des bois, et la diagnose des espèces existantes (C. ropalophorus, C. rethymnensis) est émendée sur la base du nouveau matériel. La variation des bois peut être expliquée par deux tendances évolutives : un type de parade versus un type classique de combat. La divergence est contrôlée et accélérée par la compétition intra-spécifique entre mâles. Le type classique s’explique le mieux comme le résultat d’une réduction allométrique pendant le nanisme. Le type de parade s’explique le mieux comme le résultat d’une restructuration du bauplan des bois (simplification et élongation extrême du merrain principal). Dans des scénarios supposant l’absence de prédateurs, les cerfs ont le potentiel d’évoluer des morphologies des bois et des changements de comportement inconnus sur le continent.
Mots-clés : Candiacervus ; cerf endémique ; nouvelle espèce ; radiation adaptative
Translator: Antoine Souron
Deutsche Zusammenfassung
Homogenität bei der Vielfalt: Geweihmorphologie und Evolution in einer raubtierfreien Umwelt
Die spätpleistozäne Säugetierfauna von Kreta war, wie es für Inseln typisch ist, verarmt und bestand nur aus Hirschen, Zwergelefanten, einem Otter, einer Spitzmaus und aus Riesenmäusen. Zwerghirsche (Candiacervus spp.) waren die dominanten endemischen pflanzenfressenden Arten. Hier beschreibe ich die Morphologie der adulten Geweihe dieser Hirsche. Die Variabilität der Geweihe scheint ziemlich groß, es können jedoch einige prägnante morphologische Gruppen ohne Zwischenformen unterschieden werden, die wahrscheinlich unterschiedliche Arten repräsentieren. Die Variabilität ist keine zufällige Veränderung, die durch ökologische Freisetzung in einer raubtierfreien Umwelt ausgelöst wurde. Es werden hier drei neue Arten beschrieben (Candiacervus spp. nov.), die sich auch in Geweih-und Schädelmorphologie unterscheiden und die Diagnose von existierenden Arten (C. ropalophorus, C. rethymnensis) wird auf der Basis von neuem Material korrigiert. Geweihvariation kann durch zwei evolutionäre Trends erklärt werden: Ausdrucksverhalten contra klassischer Kampftypus. Die Divergenz wird durch intraspezifischen Wettkampf zwischen Männchen angetrieben und beschleunigt. Der klassische Typus kann am besten als das Ergebnis einer allometrischen Verkleinerung während der Verzwergung erklärt werden. Der Display-Typus kann am besten als das Ergebnis einer Umstrukturierung des Geweih-Bauplans erklärt werden (Vereinfachung und Verlängerung der Hauptstange). In Szenarien ohne Räuber haben Hirsche das Potential Geweihmorphologie und Verhaltensänderungen zu entwickeln, die auf dem Festland unbekannt sind.
Schlüsselwörter: Candiacervus; endemischer Hirsch; neue Art; adaptive Radiation
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024