

 Re-evaluation of the very large Eomellivora fricki (Pia, 1939) (Carnivora, Mustelidae, Mellivorinae) from the Late Miocene of Austria
Re-evaluation of the very large Eomellivora fricki (Pia, 1939) (Carnivora, Mustelidae, Mellivorinae) from the Late Miocene of Austria
Article number: 20.1.17A
https://doi.org/10.26879/691
Copyright Palaeontological Association, April 2017
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 15 June 2016. Acceptance: 8 April 2017
{flike id=1830}
ABSTRACT
We re-evaluated the Austrian material for Hadrictis fricki Pia, 1939, from the localities Wien XII-Altmannsdorf and Gaiselberg (MN9, Vallesian, Late Miocene), concluding that Hadrictis can be considered as a synonymy of Eomellivora Zdansky, 1924; we therefore named it as Eomellivora fricki. This species is one of the earliest representatives of the genus, together with E. piveteaui Ozansoy, 1965. Our phylogenetic analyses indicate that Eomellivora forms a monophyletic group, establishing the sister clade of the large and derived Late Miocene Ekorus ekakeran. Eomellivora fricki shows a primitive dental morphology and is the largest species of the genus. This species shows the complexity of the genus Eomellivora, in which large and small species coexisted since the beginning of the Late Miocene.
Alberto Valenciano. Departamento de Geología Sedimentaria y Cambio Medioambiental. Instituto de Geociencias (CSIC, UCM). Calle José Antonio Novais 12, 28040. Madrid, Spain and Departamento de Paleontología Universidad Complutense de Madrid (UCM), Facultad de Ciencias Geológicas UCM. Calle José Antonio Novais 12, 28040, Madrid, Spain. alb3rtovv@gmail.com
Juan Abella. Universidad Estatal Península de Santa Elena. Kilómetro 1,5 de la Vía Santa Elena- La Libertad. Edificio Instituto de Investigacion Científica y Desarrollo Tecnológico INCYT), Segundo Piso. 240210 La Libertad, Ecuador. jabella@upse.edu.ec and Institut Català de Paleontologia Miquel Crusafont, Universitat Autònoma de Barcelona. Edifici Institut de Ciència i Tecnologia Ambientals - Institut Català de Paleontologia (ICTA-ICP), Campus de la UAB, Carrer de les Columnes s/n, 08193 Cerdanyola del Vallès, 08193, Barcelona, Spain. juan.abella@icp.cat
Ursula B. Göhlich. Naturhistorisches Museum Wien, Dept. Geology and Paleontology Burgring 7, 1010 Vienna, Austria. ursula.goehlich@nhm-wien.ac.at
M. Ángeles Álvarez-Sierra. Departamento de Geología Sedimentaria y Cambio Medioambiental. Instituto de Geociencias (CSIC, UCM). Calle José Antonio Novais 12, 28040. Madrid, Spain and Departamento de Paleontología Universidad Complutense de Madrid (UCM), Facultad de Ciencias Geológicas UCM. Calle José Antonio Novais 12, 28040, Madrid, Spain. masierra@ucm.es
Jorge Morales. Departamento de Paleobiología. Museo Nacional de Ciencias Naturales-CSIC, Calle José Gutiérrez Abascal, 2, 28006, Madrid, Spain. mcnm166@mncn.csic.es
Keywords: Eomellivora; Hadrictis; Mellivorinae; Mustelidae; Miocene
Final citation: Valenciano, Alberto, Abella, Juan, Göhlich, Ursula B., Álvarez-Sierra, M. Ángeles, and Morales, Jorge, 2017. Re-evaluation of the very large Eomellivora fricki (Pia, 1939) (Carnivora, Mustelidae, Mellivorinae) from the Late Miocene of Austria. Palaeontologia Electronica 20.1.17A: 1-22. https://doi.org/10.26879/691
palaeo-electronica.org/content/2017/1830-the-large-eomellivora-fricki
INTRODUCTION
Mustelidae is the most diverse family within the currently existing Carnivora, comprising 57 extant species of weasels, martens, polecats, badgers and otters (Larivière and Jennings, 2009). The Mellivorinae Gray, 1865, is the subfamily containing Mellivora capensis (Schreber, 1776) as the only living representative, commonly known as the ratel, or honey badger. It is the largest African terrestrial mustelid, weighing between 6.2 and 13.6 kg, and with a distribution range from Africa to India (Larivière and Jennings, 2009). It is a generalist and opportunistic predator, with a wide range of prey; it presents clear regional differences in its diet, which mostly involves rodents, other carnivorans (e.g., Suricata, Felis, Cynictis, Ictonyx), amphibians, reptiles, birds and invertebrates, but also includes roots, berries and fruits (Begg et al., 2003; Larivière and Jennings, 2009). Notwithstanding the fact that only one monospecific genus has survived up to the present, this subfamily was more diversified in the past. This mellivorine diversity is well reflected in the Neogene fossil record, although the remains are mostly fragmentary (Baskin, 1998; Morales et al., 2015, Werdelin and Peigné, 2010). The origin of the subfamily is unclear, but most likely arose in Eurasia or Africa, during the Middle Miocene and early Late Miocene with Sivamellivora Kretzoi, 1942, Mellalictis Ginsburg, 1977 and Eomellivora ?tugenensis Morales and Pickford, 2005. Sivamellivora necrophila (Pilgrim, 1932) comes from the Lower Siwaliks (India), Chinji formation ca. 14-11.2 m.y.a. (Patnaik, 2013) and is based on very few teeth and an edentulous mandible. Mellalictis mellalensis Ginsburg, 1977 from Beni Mellal (Morocco) ca. 12.5-11.2 m.y.a. (Werdelin and Peigné, 2010) is known by its isolated teeth and postcranial fragments. It is uncertain, however, whether it is a Mellivorinae or a mustelinae (Bonis et al., 2009). Eomellivora ?tugenensis Morales and Pickford, 2005 from the Ngorora formation (Kenya), ca. 12 m.y.a., is a poorly known mustelid of medium size that could constitute an ancestral form of Eomellivora (Valenciano et al., 2015).
Since the beginning of the Late Miocene, new genera of mellivorines radiated throughout Eurasia and Africa, e.g., Ekorus Werdelin, 2003 (assigned to Mellivorinae in this manuscript), Eomellivora Zdansky, 1924, Erokomellivora Werdelin, 2003, Hadrictis Pia, 1939, Howellictis Bonis et al., 2009, Mellivora Storr, 1780 and Promellivora Pilgrim, 1932. Some of them, such as Ekorus and Eomellivora (= Hadrictis), reached a significantly large size, which is why they are considered as giant mustelids. Eomellivora and Ekorus were one of the largest and most hypercarnivorous mustelids ever known; they were larger than the extant wolverine, Gulo gulo, but according to their cranium dimensions, they were smaller than the Oligobuninae Megalictis ferox (Werdelin, 2003; Valenciano et al., 2015, 2016). Eomellivora has been described in Asia, North America, Europe and Africa (e.g., Wolsan and Semenov 1996; Morales and Pickford, 2005; Valenciano et al., 2015), spanning from the Middle (MN8) to the Late Miocene (MN13). The first complete review of the genus was conducted by Wolsan and Semenov (1996); it concluded that this genus represents a single lineage of E. wimani Zdansky, 1924, which they subdivided into the chrono-subspecies E. wimani piveteaui for the Vallesian (MN9-10) specimens, and E. wimani wimani for the Turolian/Ventian (MN11-13) ones. More recently, a sample of E. piveteaui Ozansoy, 1965, from Batallones (Late Miocene, MN10, Madrid, Spain) has been described by Valenciano et al. (2015); this has enabled E. piveteaui, E. wimani, E. ursogulo (Orlov, 1948) and E. hungarica Kretzoi, 1942, to be accepted as valid species. However, neither Wolsan and Semenov (1996) nor Valenciano et al. (2015) analyzed the very large-sized mustelid Hadrictis fricki Pia, 1939.
Hadrictis fricki was described by Pia (1939) from the Austrian locality of Wien XII-Altmannsdorf, Late Miocene (MN9), by a largely fragmented skull that includes a worn P4-M1. Subsequently, Zapfe (1948) described a hemimandible with a worn dentition as H. fricki from Gaiselberg (Austria), Late Miocene (MN9). The systematic position of Hadrictis is doubtful - it constitutes a valid genus for some authors (Pia, 1939; Zapfe, 1948; Kretzoi 1942, 1965; Ozansoy, 1965; Webb, 1969; Ginsburg, 1977), whereas for others it is a synonym of Eomellivora (Werdelin, 1996, 2003; Peigné et al., 2006). However, the mandible of H. fricki described by Zapfe (1948) has been ignored and never compared with any other species of Eomellivora. The objectives of the present manuscript are twofold: (1) to re-describe and re-measure the material described by Pia (1939) and Zapfe (1948), and (2) to re-evaluate the systematic position of H. fricki in the light of the new evolutionary framework of Eomellivora proposed by Valenciano et al. (2015).
Localities and Geological Setting
 Both Austrian localities, Wien XII-Altmannsdorf and Gaiselberg, are situated in the Vienna Basin, which forms the northwestern part of the Pannonian Basin (Figure 1). During the Pannonian stage the foregone disintegration of the Central Paratethys has restricted Lake Pannon to the Pannonian Basin system. The first brackish-lacustrine and subsequent fluvial-influenced deposits of Lake Pannon are biostratigraphically subdivided into the Pannonian A to H biozones, based on molluscs (Papp, 1951). Both sites yielded a Late Miocene (Pannonian) vertebrate fauna (e.g., Pia, 1939; Thenius, 1948; Zapfe, 1949) including fossils of the three-toed horse Hippotherium sp., which FAD (First Appearance Datum) is MN9.
Both Austrian localities, Wien XII-Altmannsdorf and Gaiselberg, are situated in the Vienna Basin, which forms the northwestern part of the Pannonian Basin (Figure 1). During the Pannonian stage the foregone disintegration of the Central Paratethys has restricted Lake Pannon to the Pannonian Basin system. The first brackish-lacustrine and subsequent fluvial-influenced deposits of Lake Pannon are biostratigraphically subdivided into the Pannonian A to H biozones, based on molluscs (Papp, 1951). Both sites yielded a Late Miocene (Pannonian) vertebrate fauna (e.g., Pia, 1939; Thenius, 1948; Zapfe, 1949) including fossils of the three-toed horse Hippotherium sp., which FAD (First Appearance Datum) is MN9.
Wien XII-Altmannsdorf (Grießergasse clay and sand pit). Altmannsdorf is a quarter in the 12th urban district (Wien XII-Meidling) in the southwest of the city of Vienna. Vertebrate fossils in this area have been found mostly in historic sand and clay pits, in which the so-called “Congerientegel” (Schaffer, 1904) was exploited for brick production. Two nearby clay pits, one located in Grießergasse Street and the other in Oswaldgasse Street, have been active until about 1926 (Pia, 1939). At present, all the clay and sand pits have disappeared, and the area has become densely built up. Pia (1939) pointed out that the names of the nearby pits “Grießergasse,” “Oswaldgasse,” and even the general term “Altmannsdorf,” as the provenance for the vertebrate fossils, were used arbitrarily by the clay pit workers. However, the outcropping deposits in these nearby pits can be expected to be the same.
The fossil of Hadrictis fricki from Altmannsdorf was bought together with other vertebrate fossils (Table 1) by the Naturhistorisches Museum Wien (Vienna, Austria) in 1912 from a clay pit worker, who indicated the precise locality as “Grießergasse” (Pia, 1939). Due to the presence of the snail Melanopsis fossilis pseudoimpressa Papp, 1953, in the deposits of the “Oswaldgasse” pit (Papp, 1953), these deposits can be correlated with the biozone Pannonian D (Middle Pannonian), corresponding to the Mammalian Neogene Unit MN9 (Figure 1).
Gaiselberg. The Late Miocene deposits of Gaiselberg near Zistersdorf (Lower Austria) belong to the Hollabrunn-Mistelbach Formation, which represents sediments of a braid-delta system discharged by the paleo-Danube in the Vienna basin into Lake Pannon (Nehyba and Roetzel, 2004). Three sand pits in the village of Gaiselberg yielded an early Vallesian vertebrate assemblage (Table 2), which was studied by Zapfe (1949). The presence of the bivalve Congeria partschi, the more plesiomorphic dental morphology of Hippotherium sp. (Woodburne, 2009), and the co-occurrence of the two equids, Hippotherium (FAD) and Anchitherium (LAD), in Gaiselberg (Thenius, 1950; Daxner-Höck and Bernor, 2009) indicate an early Pannonian age, corresponding to the biozones earliest Pannonian C and earliest MN9, respectively (Figure 1).
MATERIAL AND METHODS
Nomenclature and Measurements
Dental nomenclature follows Ginsburg (1999) and Smith and Dodson (2003). Anatomical descriptions are based primarily on Scapino (1968), Turnbull (1970), Barone (1999, 2000), Waibl et al. (2005), and Evans and de Lahunta (2010, 2013). Measurements were made with Mitutoyo Absolute digital calipers accurate to 0.1 mm.
Institutional Abbreviations
AMNH, American Museum of Natural History, Division of Mammalogy, New York, USA; BAT, Batallones localities collection from the MNCN; IPUW, Institut für Paläontologie, Universität Wien, Vienna, Austria; MFGI, Geological and Geophysical Institute of Hungary, Budapest, Hungary; MNCN, Museo Nacional de Ciencias Naturales, Madrid, Spain; MNHN, Muséum National d´Historie Naturelle, Paris, France; NHMW, Naturhistorisches Museum Wien, Vienna, Austria; NRM, Naturhistoriska Rikmuseet, Stockholm, Sweden; PIN, Palaeontological Institute, Russian Academy of Sciences, Moscow, Russia; PMU, Palaeontological Museum, University of Uppsala, Uppsala, Sweden.
Studied Material
The fossil remains of E. fricki from Austria are stored in the fossil vertebrate collections of the Department of Geology and Palaentology at the NHMW, Austria: NHMW 2016/0065/0001: very fragmented skull with P4-M1 from Wien XII-Altmannsdorf, Vienna, Austria, MN9; NHMW 1977/1948/0130: partial hemimandible with broken c, alveoli of p1 and p2, a complete p4 and m1 and an alveolus for m2 from Gaiselberg, Lower Austria, Austria, MN9. For comparison we studied the lectotype of E. piveteaui (MNH-TRQ-1004), and E. piveteaui (MNH-TRQ-1005), both from Yassiören, Turkey, MN9, housed at MNHN; E. piveteaui from Batallones (see Valenciano et al., 2015), Spain, MN10, housed at MNCN; the holotype of E. ursogulo (PIN-No.268) from Grebeniki, Ukraine, MN11 housed at PIN (available as a cast at IPUW and NHMW); the holotype of E. hungarica (MFGI-Ob-2676) from Polgárdi 2, Hungary, MN13, housed at MFGI; the lectotype of E. wimani from Shangyingou (PMU-M3692 and PMU-M3693) and E. wimani from Liuwangou (PMU-M3847), both from China, MN12-13, housed at PMU; a cast of the holotype of Ekorus ekakeran from Lothagan, Kenya, (~MN12-13), housed at NRM (Lars Werdelin´s collection); Ischyrictis zibethoides (Blainville, 1842) from Sansan, France, MN6, housed at MNHN; Plesiogulo crassa Teilhard de Chardin, 1945, from Localities 30, 108 and 111, China, MN10-12 (Kurtén, 1970) housed at PMU. The fossils from Howellictis valentini from Toros Menalla 192 (Chad) (~MN12), were studied based on the publication of Bonis et al. (2009). The studied extant carnivorans were the mustelids Mellivora capensis, Pekania pennanti, Martes foina, G. gulo and the canid Canis lupus, housed at AMNH, NRM and MNCN.
Cladistic Analysis
We performed a cladistic analysis to better assess the relationships of E. fricki with other large Middle and Late Miocene taxa presenting similar hypercarnivorous dentition and with nearly complete remains, such as Ek. ekakeran, E. piveteaui, E. wimani, E. ursogulo, E. hungarica, H. valentini, I. zibethoides, P. crassa, and the living gulonines Martes foina, Pekania pennanti, G. gulo, the mellivorine M. capensis and the canid Canis lupus. We did not include in the analysis the African species of Eomellivora ?tugenensis or other mellivorines (e.g., Mellalictis, Promellivora, Erokomellivora, Mellivora benfieldi) due to the incompleteness of the preserved remains. The cladistic analysis includes 14 taxa and 70 equally weighted and unordered dental and cranial characters generated by us; some of them were modified from Bryant et al. (1993), Wolsan (1993), Baskin (2004) and Valenciano et al. (2015, 2016; Appendix 1). The matrix can be observed in Appendix 2 and Appendix 3. The analysis was performed using PAUP*4.0b10 (Swofford, 2002).
SYSTEMATIC PALAEONTOLOGY
Order CARNIVORA Bowdich, 1821
Suborder CANIFORMIA Kretzoi, 1943
Family MUSTELIDAE Fischer, 1817
Subfamily MELLIVORINAE Gray, 1865
Genus EOMELLIVORA Zdansky, 1924
Type species. Eomellivora wimani Zdansky, 1924 by original designation.
Included species. Eomellivora wimani Zdansky, 1924; Eomellivora fricki (Pia, 1939); Eomellivora hungarica Kretzoi, 1942; Eomellivora ursogulo (Orlov, 1948); Eomellivora piveteaui Ozansoy, 1965; Eomellivora ?tugenensis Morales and Pickford, 2005.
Diagnosis. Modified after Wolsan and Semenov (1996) and Werdelin and Peigné (2010). Mellivorine mustelid of large size; P1 present; P3 with one or two distal accessory cusps; P3 with the distal area thickened; P4 with a subconical protocone, and with paracone-protocone and paracone-parastyle crests; P4 protocone located in line with the parastyle; P4 parastyle poorly-developed but thickened; buccal wall of P4 with a concavity in the base of the crown between the paracone and the metastyle, exhibiting a variable degree of development; stylar area of M1 enlarged; M1 with a non-reduced metacone in the earlier species and a reduced one in the Turolian-Ventian; M1 with an arched ridge-shaped or conical protocone continuing into the mesial protocone crest, and a talon relatively equally expanded mesially and distally; premolar teeth crowned; p1 present; p2 turned buccolingually from the tooth row; p3 with a distal accessory cuspid and with the distal area thickened; p4 enlarged with a distal accessory cuspid and with a backward inclination of the main cuspid towards the m1; m1 enlarged with the m1 metaconid residual in some of the earliest species and absent in the others, in which it is replaced by a distinct crest; m1 talonid with single but strong, high and centrally positioned hypoconid; m2 elongated mesiodistally with a low crown surrounded by a cingulum and a central protoconid.
Eomellivora fricki (Pia, 1939)
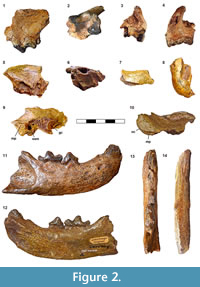
Figure 2, Table 3
1939 Hadrictis fricki Pia, p. 538, figures 1-5.
 1942 Hadrictis fricki Kretzoi, p. 319.
1942 Hadrictis fricki Kretzoi, p. 319.
1948 Hadrictis fricki Zapfe, p. 244, figure 1.
1965 Hadrictis fricki Ozansoy, p.25.
1965 Hadrictis fricki Kretzoi, p.131.
Holotype. NHMW 2016/0065/0001, comprising several small fragments of a cranium, including a portion of maxilla with P4 and M1.
Type locality. Wien XII-Altmannsdorf (Vienna, Austria).
Other locality. Gaiselberg (Lower Austria, Austria).
Stratigraphical distribution. Late Miocene (MN9).
Diagnosis. For cranium and upper dentition in Pia (1939) and for mandible and lower dentition in Zapfe (1948).
Emended diagnosis. Eomellivora of very large-size, with a relatively large P4 and M1. M1 with an enlarged stylar area, metacone not reduced, protocone ridge-edge located mesially with a prominent lingual platform. Very robust mandible, with a large c, presence of p1, rectangular p4 without a mesial accessory cuspid, a large m1, and a relatively large m2, according to its alveolus.
Differential diagnosis. Differs from all the species of Eomellivora in a larger dentition, in the presence of a residual m1 metaconid and in a stouter mandibular corpus. Differs from E. piveteaui in a bigger M1 metacone, in a larger distal platform in M1 and in a shorter mandible. Differs from E. wimani in a less concave buccal base of the P4, in a more developed M1 metacone and a non-cuspid-like M1 protocone; Furthermore, it differs in the absence of a mesial accessory cuspid in p4 and a shorter mandible. Differs from E. ursogulo in a better developed M1 metacone and a non-cuspid-like protocone; additionally it differs in the absence of a mesial accessory cuspid in p4, in possessing only one distal accessory cuspid in p4 and in a shorter mandible. Differs from E. hungarica in a much more developed M1 with a larger metacone and larger distal platform in M1 and in the absence of a mesial accessory cuspid in p4.
Description. NHMW 2016/0065/0001 comprises five little fragments of an incomplete cranium from Wien XII-Altmansdorf previously described by Pia (1939): (1) a fragment of the left maxilla with the distal part of the P3 alveoli and a worn P4-M1 (Figure 2.1-4; Table 3). It also shows the most rostral portion of the zygomatic arch and the ventral part of the infraorbital foramen; (2) an homologous edentulous fragment of the right maxilla (Figure 2.5-6) with the distal part of the P3 alveolus, the alveoli of P4 and one broken root of the M1; (3) a small fragment of the left zygomatic arch, fitting the frontal process of the zygomatic (Figure 2.7); (4) a small fragment of the left temporo-mandibular joint with a partial glenoid cavity (Figure 2.8); and (5) a fragment including the right mastoid process, the ventral part of the nuchal crest, the external auditory meatus and part of the glenoid cavity (Figure 2.9-10). Neither the alisphenoid canal nor a suprameatal fossa can be observed.
The distal part of the P3 alveolus is very wide, which indicates that the distal end of the P3 is wide. The P4 is strongly worn in the distal and occlusal area (Figure 2.1-4). It displays a very low but robust parastyle and a paracone-parastyle crest. The protocone, which is strongly worn, is subconical, robust and located in line with the mesial corner of the P4. Despite this wear, an inflection between the protocone and parastyle is visible. There is also a concavity in the buccal wall between the paracone and the metastyle. The M1 is a very large tooth with a typical morphology of Eomellivora, with the buccal wall narrower than the lingual one. It shows an enormous wear facet in the paracone, in the metacone and in the mesial part close to the protocone, as well as in the most distal corner of the tooth (Figure 2.2, 2.4). The mesial wear facet of the M1 is caused by the occlusion with the metaconid area of the m1, and the distal wear facet is produced by the occlusion with the m2. The stylar area is greatly enlarged. The paracone is conical and situated in the mesiobuccal corner. The metacone is also quite well developed and is surrounded by a distal expansion. The protocone is ridge-shaped, and it is mesiolingually located. There exists a swollen lingual platform that completely encloses the protocone.
The right hemimandible NHMW 1977/1948/0130 comes from Gaiselberg (Lower Austria) and was previously described by Zapfe (1948). The fragment has a total length of 123.37 mm. The mandible is very short, robust and dorsoventrally deep (Figure 2.11-14; Table 3). The height of the ramus, ventral to the m1, is 31 mm, and maximum width is 16.6 mm. The surface of the mandibular corpus is quite smooth, indicating some degree of alteration in its original morphology, likely due to a process of erosion by transport. The ventral margin is convex, with a medial bulge ventral to the distal part of the p4 on the ventromedial margin of the mandibular corpus. There are two rounded mental foramina, one ventral to the p2 and another ventral to the distal area of the p3. The mandibular symphysis is rather vertical and stout. The preserved dentition comprises c, p4 and m1 and the alveoli for i2-3, p1-2 and m2. The c is robust and elliptical in cross section, but the tip is broken off. The alveolus of the p1 is rounded. The p2 is two-rooted. The alveoli for the p2 are buccolingually rotated relative to the tooth row. The length of its alveoli suggests a long p2. This hemimandible has no alveoli for the p3; rather, there is a porous surface over the mandible, indicating that the tooth was lost when the animal was alive and reflects a complete closure of the alveoli. The p4 is long and sub quadrangular with a slight distal broadening. It presents no mesial accessory cuspid. The main cuspid is well worn and is inclined distally toward the m1. The distal accessory cuspid is high but also worn. The distal cingulum is high and shows a wear facet in the distobuccal corner of the p4, resulting from the occlusion with the P4. The m1 is very large and well worn. The trigonid occupies two-thirds of the total length of the tooth and bears a buccal wear facet on the trenchant blade. Between the paraconid and the protoconid there is a lingual concavity at the base of the crown. A residual metaconid can be observed (Figure 2.12). The talonid appears to have been high; however, due to its degree of wear this cannot be assured. The hypoconid is in a central position. The oval alveolus for m2 indicates a relatively large m2. It has a buccolingual constriction between the roots.
RESULTS
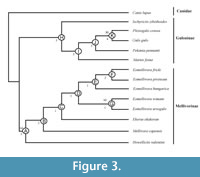 The results of our cladistics analysis provide one single most parsimonious tree (Figure 3). Apomorphies for each node are reported in Table 4. The topology of the tree clearly indicates the presence of two major clades corresponding to the subfamily Guloninae Gray, 1825, and Mellivorinae. The clade Mellivorinae comprises the taxa E. fricki, E. piveteaui, E. hungarica, E. wimani, E. ursogulo, Ek. ekakeran, M. capensis and H. valentini. This clade shares among others, the following traits (Table 4): (1) posterior lacerate foramen and jugular foramen with separate openings, the jugular foramen being in a distolateral position in relation to the lacerate foramen; (2) rostrolateral enlargement of the mastoid process; (3) high and thick mandibular corpus; (4) M1 metacunule absent; (5) p2 buccolingually rotated in relation to the tooth row; (6) p4 relatively long, with a backward inclination of the main cuspid towards the m1; (7) m1 hypoconid enlarged; and (8) absence of m1 entoconid or entocristid. Among the Mellivorinae, our analysis therefore reinforces the monophyly of Eomellivora, now also represented by E. fricki (Figure 3). We detected a Vallesian clade of Eomellivora consisting of the Euroasiatic E. piveteaui and the Centro-European E. fricki which conform the sister group of the Ventian E. hungarica. These three taxa are allied as a sister group of the Turolian/Ventian clade comprising the Euroasiatic E. wimani and the East-European E. ursogulo (Figure 3). The African Ekorus is nested as the sister group of Eomellivora. These two mustelids are associated with the following synapomorphies (Table 4): (1) straight shape of the upper incisor row; (2) position of the infraorbital foramen situated below the P4 parastyle; (3) P3 with a conspicuous concavity in the buccal wall; (4) enlarged M1 stylar area; (5) m1 hypoconid in a central position; and (6) m1 talonid without a basin, in which the hypoconid is orientated toward the m1 protoconid. Both Eomellivora and Ekorus appear as the sister group of the extant M. capensis, sharing a robust P3 with a strong cingulum, such as a widened p3. Howellictis valentini is placed in our tree as a primitive and basal mellivorine, which is in agreement with Bonis et al. (2009). The Guloninae clade (Table 4, node H), which in our analysis comprises M. foina, P. pennanti, G. gulo, P. crassa and I. zibethoides, shares among others, the following traits: (1) absence of the P3 distal accessory cusp; (2) length of the p2 relatively not reduced in relation to p3; and (3) m1 with an open and shallow talonid, with a low entocristid and a beveled lingual wall of the talonid (less marked in G. gulo). There is a significantly well-supported node composed of Plesiogulo crassa and the living wolverine G. gulo. Ischyrictis zibethoides is placed in the tree as a basal Guloninae.
The results of our cladistics analysis provide one single most parsimonious tree (Figure 3). Apomorphies for each node are reported in Table 4. The topology of the tree clearly indicates the presence of two major clades corresponding to the subfamily Guloninae Gray, 1825, and Mellivorinae. The clade Mellivorinae comprises the taxa E. fricki, E. piveteaui, E. hungarica, E. wimani, E. ursogulo, Ek. ekakeran, M. capensis and H. valentini. This clade shares among others, the following traits (Table 4): (1) posterior lacerate foramen and jugular foramen with separate openings, the jugular foramen being in a distolateral position in relation to the lacerate foramen; (2) rostrolateral enlargement of the mastoid process; (3) high and thick mandibular corpus; (4) M1 metacunule absent; (5) p2 buccolingually rotated in relation to the tooth row; (6) p4 relatively long, with a backward inclination of the main cuspid towards the m1; (7) m1 hypoconid enlarged; and (8) absence of m1 entoconid or entocristid. Among the Mellivorinae, our analysis therefore reinforces the monophyly of Eomellivora, now also represented by E. fricki (Figure 3). We detected a Vallesian clade of Eomellivora consisting of the Euroasiatic E. piveteaui and the Centro-European E. fricki which conform the sister group of the Ventian E. hungarica. These three taxa are allied as a sister group of the Turolian/Ventian clade comprising the Euroasiatic E. wimani and the East-European E. ursogulo (Figure 3). The African Ekorus is nested as the sister group of Eomellivora. These two mustelids are associated with the following synapomorphies (Table 4): (1) straight shape of the upper incisor row; (2) position of the infraorbital foramen situated below the P4 parastyle; (3) P3 with a conspicuous concavity in the buccal wall; (4) enlarged M1 stylar area; (5) m1 hypoconid in a central position; and (6) m1 talonid without a basin, in which the hypoconid is orientated toward the m1 protoconid. Both Eomellivora and Ekorus appear as the sister group of the extant M. capensis, sharing a robust P3 with a strong cingulum, such as a widened p3. Howellictis valentini is placed in our tree as a primitive and basal mellivorine, which is in agreement with Bonis et al. (2009). The Guloninae clade (Table 4, node H), which in our analysis comprises M. foina, P. pennanti, G. gulo, P. crassa and I. zibethoides, shares among others, the following traits: (1) absence of the P3 distal accessory cusp; (2) length of the p2 relatively not reduced in relation to p3; and (3) m1 with an open and shallow talonid, with a low entocristid and a beveled lingual wall of the talonid (less marked in G. gulo). There is a significantly well-supported node composed of Plesiogulo crassa and the living wolverine G. gulo. Ischyrictis zibethoides is placed in the tree as a basal Guloninae.
DISCUSSION
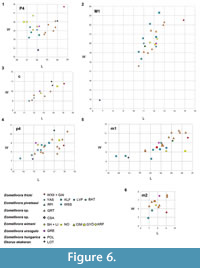
 Eomellivora (Hadrictis) fricki is one of the largest extinct mustelids from Europe, together with the gulonine Plesiogulo monspessulanus Viret, 1939, from Montpellier (France, MN14); they both remarkably exceed the size of the wolverine, the largest living terrestrial mustelid. Since Zapfe (1948), nobody has appropriately considered the giant Austrian taxon within the broader context of the large Miocene mustelids. Eomellivora fricki possesses a clearly Eomellivora-like morphology in its dentition, which relates it to the species of the genus Eomellivora (Figure 4, Figure 5, Table 4), more than with other extinct genera. It shares with the Eomellivora species the following characters: distal area of P3 thickened distally; P4 robust with a subconical protocone; M1 with an enlarged stylar area; M1 protocone higher than the metacone, with a lingual M1 platform mesiodistally enlarged; p4 relatively enlarged, main cuspid noticeably inclined towards the m1, and the distal area of p4 thickened buccolingually with a quadrangular shape in the occlusal view; narrow m1 talonid with an m1 hypoconid centrally positioned and orientated toward the m1 protoconid. According to all the features shared, and supported by our cladistics results, we conclude that Hadrictis is a synonymy of Eomellivora, and we therefore named it as Eomellivora fricki. However, E. fricki presents several diagnostic traits that make it a valid species of Eomellivora, e.g., a longer dentition (Figure 6), the presence of a residual m1 metaconid and a stouter mandibular corpus. Due to the very early Vallesian age of the Austrian species, the residual m1 metaconid is considered to constitute a primitive trait, which is lost in the other species of the genus. Such a loss is a very common feature along the evolution of mustelids, also occurring in some other genera (e.g., Hoplictis, Plesiogulo or Megalictis) in which some of their species became more hypercarnivorous (e.g., Hendey, 1978; Harrison, 1981; Ginsburg, 1999; Valenciano et al., 2016).
Eomellivora (Hadrictis) fricki is one of the largest extinct mustelids from Europe, together with the gulonine Plesiogulo monspessulanus Viret, 1939, from Montpellier (France, MN14); they both remarkably exceed the size of the wolverine, the largest living terrestrial mustelid. Since Zapfe (1948), nobody has appropriately considered the giant Austrian taxon within the broader context of the large Miocene mustelids. Eomellivora fricki possesses a clearly Eomellivora-like morphology in its dentition, which relates it to the species of the genus Eomellivora (Figure 4, Figure 5, Table 4), more than with other extinct genera. It shares with the Eomellivora species the following characters: distal area of P3 thickened distally; P4 robust with a subconical protocone; M1 with an enlarged stylar area; M1 protocone higher than the metacone, with a lingual M1 platform mesiodistally enlarged; p4 relatively enlarged, main cuspid noticeably inclined towards the m1, and the distal area of p4 thickened buccolingually with a quadrangular shape in the occlusal view; narrow m1 talonid with an m1 hypoconid centrally positioned and orientated toward the m1 protoconid. According to all the features shared, and supported by our cladistics results, we conclude that Hadrictis is a synonymy of Eomellivora, and we therefore named it as Eomellivora fricki. However, E. fricki presents several diagnostic traits that make it a valid species of Eomellivora, e.g., a longer dentition (Figure 6), the presence of a residual m1 metaconid and a stouter mandibular corpus. Due to the very early Vallesian age of the Austrian species, the residual m1 metaconid is considered to constitute a primitive trait, which is lost in the other species of the genus. Such a loss is a very common feature along the evolution of mustelids, also occurring in some other genera (e.g., Hoplictis, Plesiogulo or Megalictis) in which some of their species became more hypercarnivorous (e.g., Hendey, 1978; Harrison, 1981; Ginsburg, 1999; Valenciano et al., 2016).
 During the early Vallesian (MN9) E. fricki coexists in Central Europe with E. piveteaui, spanning from Europe and Turkey during the MN9-10. Both large mustelids are nested in the same clade (Figure 3), and share a primitive dentition, e.g., the two lack a marked concavity in the buccal base of the P4, which is present in the other species of Eomellivora; a M1 metacone not reduced, with an M1 protocone ridge-shaped and mesiolingually located, and absence of a mesial accessory cuspid in the p4 (Figure 2, Figure 4.2, Figure 5.3-4). Nonetheless, E. fricki differs from E. piveteaui in a shorter mandible and in an M1 with a stronger developed metacone and with a larger distal platform. Eomellivora fricki from Wien XII- Altmannsdorf is the largest Eomellivora (Figure 6), whereas the one from Gaiselberg overlaps with the size range of the larger specimens from Gritsev (Ukraine), MN9, which are only known by their dental dimensions (Wolsan and Semenov, 1996) (Figure 6). The dental sample of Eomellivora from Gritsev is the largest of the genus, even more so than the sample from Batallones (Wolsan and Semenov, 1996; Valenciano et al., 2015), and only a complete description and figuration of these fossils could elucidate the relationship between the Vallesian E. fricki and E. piveteaui ; this would provide valuable data on the intraspecific variability of Eomellivora.
During the early Vallesian (MN9) E. fricki coexists in Central Europe with E. piveteaui, spanning from Europe and Turkey during the MN9-10. Both large mustelids are nested in the same clade (Figure 3), and share a primitive dentition, e.g., the two lack a marked concavity in the buccal base of the P4, which is present in the other species of Eomellivora; a M1 metacone not reduced, with an M1 protocone ridge-shaped and mesiolingually located, and absence of a mesial accessory cuspid in the p4 (Figure 2, Figure 4.2, Figure 5.3-4). Nonetheless, E. fricki differs from E. piveteaui in a shorter mandible and in an M1 with a stronger developed metacone and with a larger distal platform. Eomellivora fricki from Wien XII- Altmannsdorf is the largest Eomellivora (Figure 6), whereas the one from Gaiselberg overlaps with the size range of the larger specimens from Gritsev (Ukraine), MN9, which are only known by their dental dimensions (Wolsan and Semenov, 1996) (Figure 6). The dental sample of Eomellivora from Gritsev is the largest of the genus, even more so than the sample from Batallones (Wolsan and Semenov, 1996; Valenciano et al., 2015), and only a complete description and figuration of these fossils could elucidate the relationship between the Vallesian E. fricki and E. piveteaui ; this would provide valuable data on the intraspecific variability of Eomellivora.
 The fossil material of E. hungarica from Polgárdi 2 (MN13) is poorly preserved (Figure 4.6, Figure 5.9-10) and hinders comparisons with E. fricki. Both species have a large c, p4 and m1, showing a stout m1 talonid with a quadrangular shape in the occlusal view. Eomellivora fricki, however, differs from E. hungarica in a much more developed M1 with a bigger M1 metacone, in a larger distal platform in the M1 and in the absence of a mesial accessory cuspid in the p4. Our phylogenetic hypothesis suggests a closer relationship between E. hungarica and the Vallesian clade than with the latest E. wimani and E. ursogulo (Figure 3). Eomellivora hungarica, E. fricki and E. piveteaui possess the subsequent characters (Table 4): (1) a non-cusp-like M1 protocone (synapomorphy); (2) a relatively robust p2; and (3) m1 protoconid and paraconid similar in height. This phylogenetic hypothesis could also be justified by the large size of the dentition and the apparently close morphology in the talonid of the m1 of E. hungarica and E. fricki and by the very similar morphology of the M1 between E. hungarica and E. piveteaui. Unfortunately, only a greater amount of material could clarify the systematic position of this Eomellivora from Polgárdi 2.
The fossil material of E. hungarica from Polgárdi 2 (MN13) is poorly preserved (Figure 4.6, Figure 5.9-10) and hinders comparisons with E. fricki. Both species have a large c, p4 and m1, showing a stout m1 talonid with a quadrangular shape in the occlusal view. Eomellivora fricki, however, differs from E. hungarica in a much more developed M1 with a bigger M1 metacone, in a larger distal platform in the M1 and in the absence of a mesial accessory cuspid in the p4. Our phylogenetic hypothesis suggests a closer relationship between E. hungarica and the Vallesian clade than with the latest E. wimani and E. ursogulo (Figure 3). Eomellivora hungarica, E. fricki and E. piveteaui possess the subsequent characters (Table 4): (1) a non-cusp-like M1 protocone (synapomorphy); (2) a relatively robust p2; and (3) m1 protoconid and paraconid similar in height. This phylogenetic hypothesis could also be justified by the large size of the dentition and the apparently close morphology in the talonid of the m1 of E. hungarica and E. fricki and by the very similar morphology of the M1 between E. hungarica and E. piveteaui. Unfortunately, only a greater amount of material could clarify the systematic position of this Eomellivora from Polgárdi 2.
Eomellivora ursogulo from Grebeniki (Ukraine), MN11 (Figure 4.4, 4.8, Figure 5.5-6) and E. wimani from Shangyingou (locality 12) (Figure 4.5, Figure 5.7-8) and Liuwangou (locality 31) MN12-13 from China are characterized by a relatively stouter dentition (Figure 6), with more accessory cusps and cuspids, and herein they differ from the Vallesian species (E. fricki and E. piveteaui). Eomellivora fricki differs from the Holarctic E. wimani and the Ukrainian E. ursogulo in a more developed M1 metacone, in a non-cusp-like M1 protocone, in the absence of a mesial accessory cuspid in the p4 and by a shorter mandible. Furthermore, E. fricki differs from E. wimani in a weaker concavity in the buccal base of the P4 and from E. ursogulo by a single distal accessory cuspid in the p4, whereas E. ursogulo possesses two.
The suprageneric taxonomy of M. capensis, G. gulo and other living mustelids has been discussed over time (e.g., Pia, 1939; Webb, 1969; Ginsburg, 1977; Ginsburg and Morales, 1992; McKenna and Bell, 1997; Baskin, 1998; Ginsburg, 1999). Mellivora capensis is currently considered to be the only living representative of Mellivorinae, and the subfamily Guloninae (previously referred to as Martinae) comprises the extant martens (Martes Pinel, 1792 and Charronia Gray, 1865), the South American tayra (Eira Smith, 1842) and the wolverine G. gulo (Linnaeus, 1758). The suprageneric taxonomy becomes an acute problem for the fossil forms, in some cases due to the fragmentary dental and cranial representations, and in others due to the presence of a mixture of characters shared by both subfamilies. The position of the Mellivorinae and the Guloninae subfamilies as a sister clade in our cladistics analysis enters into conflict with a previous cladistics analysis based on molecular data (e.g., Koepfli et al., 2008; Sato et al., 2012), in which Guloninae appears as the sister group of Mustelinae (weasel and relatives) and Lutrinae (otters); furthermore, the position of Mellivorinae, even it is not clear, shows a more basal position within mustelidae than Guloninae (e.g., Koepfli et al., 2008; Sato et al., 2012). However, the main goal of the study involves establishing how E. fricki is related to some similar large Miocene mustelids, and we therefore do not attempt to resolve the systematic position of the whole family.
The tree that we obtained reflects the evolutionary framework of Eomellivora proposed by Valenciano et al. (2015), in which Eomellivora is considered as a mellivorine. Werdelin (2003) noted that Ekorus, in spite of the differences in dental morphology between Eomellivora and Ekorus, could be in the same lineage. Our results confirm his idea and Ekorus is assigned to the Mellivorinae subfamily. The systematic position of the medium-sized ischyrictini Ischyrictis zibethoides from the middle Miocene of Europe has been ambiguous. The term Ischyrictini was erected by Pia (1939) as a “sub-subfamily,” a group of large mustelids that included the genera Ischyrictis, Laphictis and Hadrictis. Subsequently, Tobien (1955) considered the Ischyrictini as tribal level, constituting a valid rank. Concerning different authors (Pia, 1939; Tobien, 1955; Webb, 1969; Ginsburg, 1977, 1999), this tribe has been assigned to Mellivorinae and Guloninae, with different degrees of relationship with the genera Eomellivora, Hadrictis, Hoplictis, Iberictis, Gulo, Laphictis, Mellalictis, Mellivora and Plesiogulo. Additionally, Ginsburg and Morales (1992) suggested a direct relationship between Eomellivora and I. zibethoides, a consideration that is rejected on the basis of our analysis. We considered I. zibethoides as a basal guloninae without a sister group relationship with Eomellivora. Furthermore, the primitive dentition of I. zibethoides bears a resemblance to the living martens (e.g., M. foina or M. martes), with a close morphology in the P4-M1 and m1 , but with a bigger size and a more primitive dentition within the guloninae clade (e.g., presence of a distal accessory cuspid in p3, a distal accessory cuspid in p4 not reduced, m2 paraconid present), including some derived traits such as a high mandibular corpus, a robust p2, a robust P4 or an enlarged M1 stylar area. The systematic relationship of the wolverine-like mustelid Plesiogulo, ranging in body size from large to gigantic, is controversial. Some authors (e.g., Viret, 1939; Kurtén, 1970; Kurtén and Anderson, 1980) considered it to be directly related to G. gulo, whereas others have seen it as a separate phylogenetic lineage without descendants (Zdansky, 1924; Hendey, 1978; Harrison, 1981; Xiaofeng and Haipo, 1987; Alcalá et al., 1994; Sotnikova, 1995; Montoya et al., 2011). The morphological similarities between G. gulo and P. crassa, suggest a close relationship between both genera, supported by the synapopomorphy p2-4, which presents a strong cingula surrounding the entire tooth, with a thickening of the mesial and distal cristids of these teeth; as for several synapomorphies (Table 4), the most remarkable one involves a P4 more individualized protocone, located in a distal position in relation to the P4 parastyle. Nonetheless, further in-depth study of additional related taxa (e.g., other Plesiogulo species, Iberictis, Dehmictis, Ischyritis) are likely vital with regard to elucidating the relationship between the living wolverine and Plesiogulo.
CONCLUSIONS
The species Hadrictis fricki from the Austrian early Vallesian localities (MN9) can be referred to the genus Eomellivora, and we therefore synonymize Hadrictis with Eomellivora. This species represents the largest one, as well as one of the most primitive Eomellivora, which shows the complexity of the genus, in which larger (E. fricki) and relatively smaller (E. piveteaui ) species coexisted since the early Vallesian. For the first time, the phylogenetic analysis resulted in the assignation of Ek. ekakeran to the Mellivorinae subfamily, and Ekorus is allied with Eomellivora as a sister genus. Additionally, we propose a new reassignment of some extinct large mustelids at the subfamily level, I. zibethoides being placed in our phylogeny as a basal guloninae, and P. crassa as sister to the extant wolverine.
ACKNOWLEDGEMENTS
We would like to thank the curators J.O.R. Ebbestad and V. Berg-Madsen of the Palaeontological Museum, University of Uppsala, (Uppsala, Sweden), D. Kalthoff of the Naturhistoriska Rikmuseet (Stockholm, Sweden), E. López Errasquin of the Museo Nacional de Ciencias Naturales (Madrid, Spain), K. Rauscher from the Institut Fur Palaontologie der Universitat Wien, Department of Palaeontology (Vienna, Austria), C. Argot of the MNHN, and E. Westwig of the American Museum of Natural History, Division of Mammalogy (New York, USA), for access to comparative material under his care. We are grateful to L. Werdelin of the Naturhistoriska Rikmuseet (Stockholm, Sweden) for kindly allowing us to study the cast of the skull and mandibles of the Holotype of Ek. Ekakeran. Our thanks also to S. Peigné and P. Loubry (photographer UMR7207), both of the Muséum National d´Historie Naturelle (Paris, France), for pictures of the Lectotype of Eomellivora piveteaui from Yassiören, to M. Sotnikova, Geological Institute, Russian Academy of Sciences (Moscow, Russia), for pictures of the holotype of E. ursogulo from Grebeniki, and to L. Kordos and K. Palotás, both of the Geological and Geophysical Institute of Hungary (Budapest, Hungary), for pictures of the holotype of E. hungarica from Polgárdi 2. Thanks are due to Cormac de Brun for the review of the manuscript. A.V. is a trainee researcher in the CSIC program JAE-PRE_CP2011 (CSIC program "Junta para la 707 ampliación de estudios"), co-funded by the European Social Fund. A.V. also received support for his research through the project SYNTHESYS3, http://www.synthesys.info/ (SYNTHESYS; AT-TAF-5457), financed by the European Community Research Infrastructure Action under the FP7 “Capacities” Programme, and the European Union´s Seventh Framework Programme (FP7/2007-2013) under grant agreement nº 226506 (SYNTHESYS; SE-TAF-3637). A.V. also received support from an AMNH Collection Study Grant Program (2014). J.A. wishes to thank the “Proyecto Prometeo” of the “Secretaria de Educación Superior, Ciencia Tecnología e Innovación”, Republic of Ecuador. This study was also supported by the Spanish Ministerio de Economia y Competitividad (Economy and Competitiveness Ministry) research project CGL2015-68333 (MINECO/FEDER, UE), CGL2011-28681, CGL2011-28877 and the research group BSCH-UCM 910607. We are also indebted to the Handling Editor A. Souron, the Style and Proofing Editor C. Grohé and two anonymous reviewers for their useful comments and suggestions, which greatly improved the original manuscript.
REFERENCES
Alcalá, L., Montoya, P., and Morales, J. 1994. New large mustelids from the Late Miocene of Teruel Basin (Spain). Comptes Rendus de l’Académie des Sciences de Paris, 319:1093-1100.
Barone, R. 1999. Anatomie Comparée des Mammifères Domestiques, Tome 1, Ostéologie4th edition. Vigot, Paris.
Barone, R. 2000. Anatomie Comparée des Mammifères Domestiques, Tome 2, Antrologie et Myologie4 th edition. Vigot, Paris.
Baskin, J.A. 1998. Mustelidae, p. 152-173. In Janis, C.M., Scott, K.M., and Jacobs, L.L. (eds.), Evolution of Tertiary Mammals of North America, Volume 1: Terrestrial Carnivores, Ungulates, and Ungulate-Like Mammals. Cambridge University Press, Cambridge.
Baskin, J.A. 2004. Bassariscus and Probassariscus (Mammalia, Carnivora, Procyonidae) from the Early Barstovian (Middle Miocene). Journal of Vertebrate Paleontology, 24:709-720.
Begg, C.M., Begg, K.S., Du Toit, J.T., and Mills, M.G.L. 2003. Sexual and seasonal variation in the diet and foraging behavior of a sexually dimorphic carnivore, the honey badger (Mellivora capensis). The Zoological Society of London, 260:301-316.
Blainville, H. 1842. Ostéographie ou description iconographique comparée du squelette et du système dentaire des cinq classes d’animaux vertébrés récents et fossiles pour servir de base à la zoologie et à la géologie. Mammiferes Carnassiers: Des Mustelas (Genre Mustela L.), 10:1-83.
Bowdich, T.E. 1821. An Analysis of the Natural Classifications of Mammalia, for the Use of Students and Travellers. J. Smith, Paris.
Bryant, H.N., Russell, A.P., and Fitch, W.D. 1993. Phylogenetic relationships within the extant Mustelidae (Carnivora): appraisal of the cladistic status of the Simpsonian subfamilies. Zoological Journal of the Linnean Society ,108:301-334.
Crusafont-Pairó, M. and Ginsburg, L. 1973. Les carnassiers fossiles de Los Valles de Fuentidueña. Bulletin du Muséum National d´Histoire Naturelle, 3e série, 131:29-45.
Daxner-Höck, G. and Bernor, R.L. 2009. The Early Vallesian vertebrates from Atzelsdorf (Austria, Late Miocene). 8. Anchitherium, Suidae and Castoridae (Mammalia). Annalen des Naturhistorischen Museums in Wien, Serie A, 111:557-584.
de Bonis, L., Peigné, S., Guy, F., Likius, A., Makaye, H.T., Vignaud, P., and Brunet, M. 2009. A new mellivorine (Carnivora, Mustelidae) from the Late Miocene of Toros Menalla, Chad. Neues Jahrbuch für Geologie und Paläontologie , 252:33-54.
Evans, H.E. and de Lahunta, A. 2010. Miller´s Guide to the Dissection of the Dog 4th edition. W.B. Saunders Company, Philadelphia.
Evans H.E. and de Lahunta A. 2013. Miller´s Anatomy of the Dog 4th edition. W.B. Saunders Company, Philadelphia.
Fischer, G. 1817. Adversaria zoological. Mémoires de la société Imperiale des Naturalistes de Moscou, 5:368- 428.
Ginsburg, L. 1977. Les carnivores du Miocène de Beni Mellal. Géologie Méditerranéenne, 4:225-240.
Ginsburg, L. 1999. Order Carnivora, p. 109-148. In Rössner, G.E. and Heissig, K. (eds.), The Miocene Land Mammals of Europe. Friedrich Pfeil, München.
Ginsburg, L. and Morales, J. 1992. Contribution à la connaissance des Mustélidés (Carnivora, Mammalia) du Miocène d’Europe Trochictis et Ischyrictis, genres affines et genres nouveaux. Comptes Rendus de l’Académie des Sciences de Paris, 315:111-116.
Gray, J.E. 1825. An outline of an attempt at the disposition of Mammalia into tribes and families, with a list of the genera apparently appertaining to each tribe. Annals of Philosophy (New Ser.), 10:337-344.
Gray, J.E. 1865. Revision of the genera and species of Mustelidae contained in the British Museum. Proceedings of the Geological Society of London, 1865:100-154.
Harrison, J.A. 1981. A review of the extinct wolverine, Plesiogulo (Carnivora: Mustelidae), from North America. Smithsonian Contributions to Paleobiology, 46:1-27.
Harzhauser, M., Daxner-Höck, G., and Piller, W.E. 2004. An integrated stratigraphy of the Pannonian (Late Miocene) in the Vienna Basin. Austrian Journal of Earth Sciences, 95/96:6-19.
Hendey, Q.B. 1978. Late Tertiary Mustelidae (Mammalia, Carnivora) from Langebaanweg, South Africa. Annals of the South African Museum, 76:329 - 357.
Koepfli, K.P., Deere, K.A., Slater, GJ., Begg, C., Begg, K., Grassman, L., Lucherini, M., Veron, G., and Wayne, RK. 2008. Multigene phylogeny of the Mustelidae: resolving relationships, tempo and biogeographic history of a mammalian adaptive radiation. BMC Biology 6:10.
Koufos, G.D. 2012. New material of Carnivora (Mammalia) from the Late Miocene of Axios Valley, Macedonia, Greece. Comptes Rendus Palevol, 11:49-64.
Kretzoi, M. 1942. Eomellivora von Polgárdi und Csákvár. Földtani Közlöny, 72:318 - 323.
Kretzoi, M. 1943. Kochictis centennii n. g. n. sp., ein altertümlicher Creodonte aus dem Oberoligozän Siebenbürgens. Földtani Közlöny, 73:190-195.
Kretzoi, M. 1965. Die Hipparion-Fauna von Györszentmárton in NW-Ungarn. Annales Historico-Naturales Musei Nationalis Hungarici, 57:127-143.
Kurtén, B. 1970. The Neogene wolverine Plesiogulo and the origin of Gulo (Carnivora, Mammalia). Acta Zoologica Fennica, 131:1-22.
Kurtén, B and Anderson, E. 1980 . Pleistocene Mammals of North America. Columbia University Press, New York.
Larivière, S. and Jennings, A.P. 2009. Family Mustelidae (Weasels and Relatives), p. 564-656. In Wilson, D.E. and Mittermeier, R.A. (eds.), Handbook of Mammals of the World. 1. Carnivores. Lynx Editions, Barcelona.
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secumdum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Laurentii Solvii, Stockholm.
Lungu, A.N. 1978. Hipparion Fauna of Middle Sarmatian of Moldova: Carnivores. Shtintsa, Kishinev.
McKenna, M.C. and Bell, S.K. 1997.Classification of mammals above the species level. Columbia University Press, New York.
Montoya, P., Morales, J., and Abella, J. 2011. Musteloidea (Carnivora, Mammalia) from the Late Miocene of Venta del Moro (Valencia, Spain). Estudios Geológicos, 67:193-206.
Morales, J. and Pickford, M. 2005. Carnivores from the Middle Miocene Ngorora Formation (13-12 Ma), Kenya. Estudios Geológicos, 61:271 - 284.
Morales, J., Cantalapiedra, J.L., Valenciano, A., Hontecillas, D., Fraile, S., García Yelo, B.A., Montoya, P., and Abella, J. 2015. The fossil record of the Neogene canivore mammals from Spain. Palaeobiodiversity and Palaeoenvironments, 95:373-386.
Nehyba, S. and Roetzel, R. 2004. The Hollabrunn-Mistelbach Formation (Upper Miocene, Pannonian) in the Alpine-Carpathian Foredeep and the Vienna Basin in Lower Austria - An example of a coarse-grained fluvial system. Jahrbuch der Geologischen Bundesanstalt, 144:191-221.
Orlov, J.A. 1948. Perunium ursogulo Orlov, a new gigantic extinct mustelid (A contribution to the morphology of the skull and brain and to the phylogeny of Mustelidae). Acta Zoologica, 29:63 - 105.
Ozansoy, F. 1965. Étude des gisements continentaux et des mammifères du Cénozoïque de Turquie. Mémoires de la Société Géologique de France, Nouvelle Série, 44:1-92.
Papp, A. 1951. Das Pannon des Wiener Beckens. Mitteilungen der Geologischen Gesellschaft in Wien, 39-41:99-193.
Papp, A. 1953. Die Molluskenfauna des Pannon des Wiener Beckens. Mitteilungen der geologischen Gesellschaft in Wien, 44:85-222.
Patnaik, R. 2013. Indian Neogene Siwalik Mammalian Biostratigraphy: An Overview, p. 423-444. In Wang, X., Flynn, L.J., and Fortelius M. (eds.), Fossil Mammals of Asia: Neogene Biostratigraphy and Chronology. Columbia University Press, New York.
Peigné, S., Chaimanee, Y., Yamee, C., Srisuk, P., Marandat, B., and Jaeger J.J. 2006. A new member of the mustelida (Mammalia: Carnivora) from the Paleogene of Southern Asia. Journal of Vertebrate Paleontology, 26:788-793.
Pia, J. 1939. Ein riesiger Honingsdachs (Mellivorine) aus dem Unterpliozän von Wien. Annalen des Naturhistorischen Museums in Wien, 50:537-583.
Pickford, M. 2016. Biochronology of European Miocene Tetraconodontinae (Suidae, Artiodactyla, Mammalia) flowing from recent revision of the Subfamily. Annalen des Naturhistorischen Museums in Wien, Serie A, 118:175-244.
Pilgrim, G.E. 1932. The fossil Carnivora of India. Memoirs of the Geological Survey of India. Palaeontologia Indica, 18:1-232.
Pinel, P. 1792. Recherches sur une nouvelle méthode de classification des quadrupèdes, fondée sur la structure mécanique des parties osseuses qui servent à l’articulation de la mâchoire inférieure. Actes de la Societé d’Histoire Naturelle de Paris, 1:50-60.
Sato, J.J., Wolsan, M., Prevosti, F.J., D´Elía, G., Begg, C., Begg, K., Hosoda, T., Campbell, K.L., and Suzuki, H. 2012. Evolutionary and biogeographic history of weasel-like carnivorans (Musteloidea). Molecular Phylogenetics and Evolution 63:745-757.
Scapino, R.P. 1968. Biomechanics of feeding in Carnivora. PhD dissertation, University of Illinois, Chicago, Illinois, USA.
Schaffer, F.X. 1904. Geologie von Wien, Teil I. Wien. R. Lechner, Vienna.
Schreber, J.C.D. 1776. Die Säugethiere in Abbildungen nach der Natur, mit Beschreibungen. Wolfgang Walther, Erlangen, Germany.
Smith, C.H. 1842. Introduction to the Mammalia. The Naturalist’s Library, 13:1-299.
Smith, J.B. and Dodson, P. 2003. A proposal for a standard terminology of anatomical notation and orientation in fossil vertebrate dentitions. Journal of Vertebrate Paleontology, 23:1-12.
Sotnikova, M.V. 1995. Extinct wolverine from the Former USSR: Review of the genus Plesiogulo (Carnivora: Mustelidae). Lutreola, 6:1-8.
Stock, C. and Hall, E.R. 1933. The Asiatic genus Eomellivora in the Pliocene of California. Journal of Mammalogy, 14:63-65.
Storr, G.C.C. 1780. Prodromus methodi mammalium. Litteris Reissianis, Tubingae, Germany.
Swofford, D.L. 2002. PAUP*: Phylogenetic Analysis using Parsimony (*and other methods), version 4. Sinauer Associates, Sunderland, Massachusetts.
Teilhard de Chardin, P. 1945. Les Mustélidés de Chine. Institut de Géobiologie, 12:1-56.
Thenius, E. 1948. Über die Entwicklung des Hornzapfens von Miotragocerus. Sitzungsberichte der Österreichischen Akademie der Wissenschaften, math.-naturw. Klasse, 157:203-221.
Thenius, E. 1950. Über den Nachweis von Anchitherium aurelianense im Pannon des Wiener Beckens. Anzeiger der mathematisch-naturwissenschaftlichen Klasse der Österreichischen Akademie der Wissenschaften, 1950/8:174-181.
Tobien, H. 1955. Neue und wenig bekannte Carnivoren aus den unterpliozänen Dinotheriensanden Rheinhessens. Notizblatt des Hessischen Landesamtes fur Bodenforschung zu Wiesbaden, 83:7-31.
Turnbull, W.D. 1970. Mammalian masticatory apparatus. Fieldiana: Geology, 18:149-356.
Valenciano, A., Abella, J., Sanisidro, O., Hartstone-Rose, A., Álvarez-Sierra, M.A., and Morales, J. 2015. Complete description of the skull and mandible of the giant mustelid Eomellivora piveteaui Ozansoy, 1965 (Mammalia, Carnivora, Mustelidae) from Batallones (MN10), Late Miocene (Madrid, Spain). Journal of Vertebrate Paleontology, 35:e934570,
doi: 10.1080/02724634.2014.934570.
Valenciano, A., Baskin, J.A., Abella, J., Pérez-Ramos, A., Álvarez-Sierra, M.A., Morales, J. and Hartstone-Rose, A. 2016. Megalictis, the bone-crushing giant mustelid (Carnivora, Mustelidae, Oligobuninae) from the Early Miocene of North America. PLoS ONE 11(4):e0152430,
doi: 10.1371/journal.pone.0152430.
Viret, J. 1939. Monographie Paléontologique de la Faune de Vertébrés des Sables de Montpellier. III. Carnivora Fissipedia. Travaux du Laboratoire de Géologie de la Faculte des Sciences de Lyon, 37:7-26.
Waibl, H., Gasse, H., Hashimoto, Y., Burdas, K.D., Constantinescu, G.M., Saber, A.S., Simoens, P., Salazar, I., Sotonyi, P., Augsburger, H., and Bragulla, H. 2005. Nomina Anatomica Veterinaria 5th edition. International Committee on Veterinary Gross Anatomical Nomenclature. World Association of Veterinary Anatomists, Columbia, Missouri.
Webb, S.D. 1969. The Burge and Minnechaduza Clarendoian mammalian faunas of north-centra1 Nebraska. University of California Publications, Geological Sciences Bulletin, 78:1-191.
Werdelin, L. 1996. Carnivoran ecomorphology: A phylogenetic perspective, p. 582-624. In Gittleman J.L. (ed.), Carnivore Behavior, Ecology, and Evolution. Volume 2. Cornell University Press, Ithaca, New York.
Werdelin, L. 2003. Mio-Pliocene Carnivora from Lothagam, Kenya, p. 261-328. In Leakey, M.G. and Harris, J.M. (eds.), Lothagam, the Dawn of Humanity in Eastern Africa. Columbia University Press, New York.
Werdelin, L. and Peigné, S. 2010. Carnivora, p 603-657. In Werdelin, L. and Sanders, W.J. (eds.), Cenozoic Mammals of Africa. University of California Press, Berkeley.
Wolsan, M. 1993. Phylogeny and classification of early European Mustelida (Mammalia: Carnivora). Acta Theriologica, 38:345-384.
Wolsan, M. and Semenov, Y.A. 1996. A revision of the late Miocene mustelid carnivoran Eomellivora. Acta Zoologica Cracoviensia, 39:593-604.
Woodburne, M.O. 2009. The Early Vallesian vertebrates from Atzelsdorf (Austria, Late Miocene). 9. Hippotherium (Mammalia, Equidae). Annalen des Naturhistorischen Museums in Wien, Serie A, 111:585-604.
Xiaofeng, X. and Haipo, W. 1987. Fossil Gulo in China. Vertebrata Palasiatica 25:306-314 (In Chinese with summary in English)
Zapfe, H. 1948. Neue Funde von Raubtieren aus dem Unterpliozän des Wiener Beckens. Sitzungsberichte der Österreichische Akademie der Wissenschaften: Mathematisch-naturwissenschaftliche Klasse, 157:243-262.
Zapfe, H. 1949. Die Säugetierfauna aus dem Unterpliozän von Gaiselberg bei Zistersdorf in Niederösterreich. Jahrbuch der Geologischen Bundesanstalt, 93:83-97.
Zdansky, O. 1924. Jungtertiäre carnivoren Chinas. Palaeontologica Sinica, 2:1-149.
