MAHENGE BAT
 The specimen reported here was found at the Mahenge locality, Singida Region, north-central Tanzania, and represents the oldest placental mammal ever found south of the equator on the African continent
(Figure 1). The Mahenge fossiliferous deposits
(Figure 2, Figure
3) are composed of alternating mudstones and variably silicified shales (Harrison et al.
2001) and represent crater lake sediments deposited in the top of a kimberlite pipe.
The specimen reported here was found at the Mahenge locality, Singida Region, north-central Tanzania, and represents the oldest placental mammal ever found south of the equator on the African continent
(Figure 1). The Mahenge fossiliferous deposits
(Figure 2, Figure
3) are composed of alternating mudstones and variably silicified shales (Harrison et al.
2001) and represent crater lake sediments deposited in the top of a kimberlite pipe.
 A diversity of plants and lower vertebrates is preserved at Mahenge
(Herendeen and Jacobs 2000;
Harrison et al.
2001). The fish assemblage from Mahenge suggests an early middle Eocene
(Lutetian) age (Murray
2000). A Pb-U date of 45.83 Ma (+/- 0.17) was recently obtained from a zircon crystal at the base of the Mahenge sequence (Harrison et al. 2001). Based on studies of crater lakes in Europe and Africa, it is likely that sediments began to accumulate rather quickly in the Mahenge pipe after initial formation of the crater
(Lorenz 1973;
Smith 1986;
Rayner 1987;
Giresse et al. 1991;
Cornen et al.
1992). Given sedimentation rates and compaction factors documented in modern
maars, it can be argued that the crater at Mahenge would have been filled by sediments in 0.2-1.0 million years
(Harrison et al.
2001).
A diversity of plants and lower vertebrates is preserved at Mahenge
(Herendeen and Jacobs 2000;
Harrison et al.
2001). The fish assemblage from Mahenge suggests an early middle Eocene
(Lutetian) age (Murray
2000). A Pb-U date of 45.83 Ma (+/- 0.17) was recently obtained from a zircon crystal at the base of the Mahenge sequence (Harrison et al. 2001). Based on studies of crater lakes in Europe and Africa, it is likely that sediments began to accumulate rather quickly in the Mahenge pipe after initial formation of the crater
(Lorenz 1973;
Smith 1986;
Rayner 1987;
Giresse et al. 1991;
Cornen et al.
1992). Given sedimentation rates and compaction factors documented in modern
maars, it can be argued that the crater at Mahenge would have been filled by sediments in 0.2-1.0 million years
(Harrison et al.
2001).  Conservatively, this would place an upper age limit of approximately 45 Ma for the Mahenge sequence.
Conservatively, this would place an upper age limit of approximately 45 Ma for the Mahenge sequence.
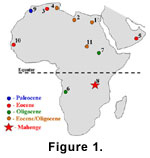
The bat consists of the anterior half of a skeleton (Figure
4) including skull, lower jaws, vertebral column, both shoulder girdles, and portions of both humeri and the left radius. The wrists, wing elements, most of the
pelves, and the hind limbs are missing. Despite repeated attempts, we were unable to locate any teeth.
 It may be that the expected (high) X-ray absorption of teeth is reduced in this specimen by demineralization in such a way that density differences between teeth and the surrounding matrix are minimal. The lack of any available tooth morphology limits our comparisons with other Eocene taxa to those that preserve comparable anatomical parts.
It may be that the expected (high) X-ray absorption of teeth is reduced in this specimen by demineralization in such a way that density differences between teeth and the surrounding matrix are minimal. The lack of any available tooth morphology limits our comparisons with other Eocene taxa to those that preserve comparable anatomical parts.
Higher level taxonomy follows Simmons
(1998) and Simmons and Geisler 1998;. However,
Springer et al. (2001) and
Teeling et al. (2002) have recently presented molecular evidence supporting microchiropteran
paraphyly. If this interpretation is true, then Tanzanycteris may be included in the suborder Yinpterochiroptera
(Springer et al.
2001).
Systematic Paleontology
Order CHIROPTERA Blumenbach, 1779
Suborder MICROCHIROPTERA Dobson, 1875
Family
TANZANYCTERIDIDAE, fam. nov.
Tanzanycteris gen. nov.
 Type Species. Tanzanycteris mannardi sp.
nov.
Type Species. Tanzanycteris mannardi sp.
nov.
Etymology. Tanzania and nykteris, Greek for bat.
Diagnosis. Differs from all comparable Eocene bats
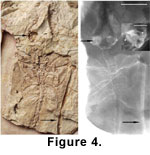
(Icaronycteris, Palaeochiropteryx, Archaeonycteris, and Hassianycteris) in having a much larger cochlear diameter relative to basicranial width
(Figure 4 and Figure
5). 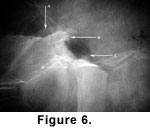 Differs from Icaronycteris and Palaeochiropteryx in having a first rib that is relatively broader than other ribs, and from Icaronycteris in having a clavicle that articulates with (or is in direct contact with) the coracoid
(Figure 6). Differs from all but Hassianycteris in having the trochiter (=greater
tuberosity) of the humerus extending proximally well beyond the humeral head (Figure 6). Further differs from Icaronycteris and Archaeonycteris in having anterior laminae on ribs and a manubrium with a bilaterally compressed ventral keel. Further differs from Palaeochiropteryx in having a narrow scapular infraspinous fossa and from Hassianycteris in having anterior laminae on ribs
(Figure 7).
Differs from Icaronycteris and Palaeochiropteryx in having a first rib that is relatively broader than other ribs, and from Icaronycteris in having a clavicle that articulates with (or is in direct contact with) the coracoid
(Figure 6). Differs from all but Hassianycteris in having the trochiter (=greater
tuberosity) of the humerus extending proximally well beyond the humeral head (Figure 6). Further differs from Icaronycteris and Archaeonycteris in having anterior laminae on ribs and a manubrium with a bilaterally compressed ventral keel. Further differs from Palaeochiropteryx in having a narrow scapular infraspinous fossa and from Hassianycteris in having anterior laminae on ribs
(Figure 7).
 Differs from all extant bats, except Rhinolophidae and Pteronotus parnellii, in having an extremely enlarged cochlea
(Figure 5). Differs from Rhinolophidae in having an enlarged cochlear
fenestra, anterior laminae on ribs, and two facets in the infraspinous fossa. Differs from P. parnellii in having a first rib that is relatively broader than other ribs, two facets in the infraspinous
fossa, a coracoid with a blunt (not flared) tip, and an iliac blade that extends dorsally beyond the level of the iliosacral articulation
(Figure 8).
Differs from all extant bats, except Rhinolophidae and Pteronotus parnellii, in having an extremely enlarged cochlea
(Figure 5). Differs from Rhinolophidae in having an enlarged cochlear
fenestra, anterior laminae on ribs, and two facets in the infraspinous fossa. Differs from P. parnellii in having a first rib that is relatively broader than other ribs, two facets in the infraspinous
fossa, a coracoid with a blunt (not flared) tip, and an iliac blade that extends dorsally beyond the level of the iliosacral articulation
(Figure 8).
We have chosen to place Tanzanycteris in a new family,
Tanzanycterididae, to emphasize the derived nature of its basicranial structures in comparison with all other Eocene
microbats.
Tanzanycteris mannardi sp. nov.
 Holotype. Tanzanian National Museum (TNM) MP-207, partial skeleton including skull, axial skeleton anterior to sacrum, partial left and right
humeri, and partial left radius. Only known specimen.
Holotype. Tanzanian National Museum (TNM) MP-207, partial skeleton including skull, axial skeleton anterior to sacrum, partial left and right
humeri, and partial left radius. Only known specimen.
Locality. Mahenge, north-central Tanzania, approximately 50 km west of
Singida. The coordinates of the Mahenge kimberlite pipe are 4º 47' 50.2" S; 34º 15' 54.5" E.
Age and Distribution. Middle Eocene, Lutetian, approximately 46 Ma
(Harrison et al. 2001).
Diagnosis. Only known species, as for genus.
Etymology. For George W. Mannard who explored the Singida Kimberlite Field and made the first excavation at the type locality in 1957
(Mannard
1962).
Discussion. Tanzanycteris mannardi is a relatively small bat
(Table 1) about the size of Palaeochiropteryx tupaiodon from the Lutetian of Europe
(Habersetzer and Storch
1987). T. mannardi (Figure
6, Figure 7, and Figure
8) has unfused vertebrae throughout the column, an iliac blade that extends dorsally beyond the level of the iliosacral articulation, a scapula with a double-faceted infraspinous
fossa, a ventrolaterally curving coracoid with a blunt tip, and ribs with posterior
laminae. Among Eocene bat taxa, Tanzanycteris is most similar to Hassianycteris, but differs in the size of the cochlea (much larger in Tanzanycteris) and presence of anterior laminae on ribs (absent in Hassianycteris).
In other features T. mannardi differs from archaic bats and resembles several extant echolocating bats. Among extant Old World bats, T. mannardi is most similar to rhinopomatoids and rhinolophoids (excluding
Nycteridae) (Simmons 1998;
Simmons and Geisler 1998;
Teeling et al.
2002). Features shared in common between rhinopomatoids and Tanzanycteris include an enlarged cochlear fenestra (also shared by mystacinids and
noctilionoids), a clavicle that articulates with the coracoid (craseonycterids only, also shared by
emballonurids, phyllostomatids, and mormoopids), and a trochiter that extends proximally well beyond the humeral head
(craseonycterids only, also shared by rhinolophoids and most yangochiropterans). None of these shared character states are exclusive to Tanzanycteris and
rhinopomatids.
Character states shared by Tanzanycteris and rhinolophoids include: extremely enlarged cochlea (absent in
megadermatids, present in one mormoopid); first rib broader than all other ribs; clavicle that articulates with the coracoid (also
craseonycterids, emballonurids, phyllostomatids, and mormoopids); trochiter that extends proximally well beyond the humeral head (also
craseonycterids, most yangochiropterans, and Hassianycteris); and iliac blade that flares dorsally resulting in a well-developed iliac fossa
(rhinolophids and Icaronycteris, Hassianycteris, and Archaeonycteris). Of these character states, the presence of a broad first rib is uniquely shared between Tanzanycteris and
rhinolophoids. Among other characteristics shared between Tanzanycteris and
rhinolophoids, all but the flaring iliac blade are present in other extant groups and several other Eocene
taxa. The flaring iliac blade with a well-developed iliac fossa is likely primitive for bats.