INTRODUCTION
After many years examining field and well samples of Nummulites
and Discocyclina dominated limestones (Eocene, generally shallow marine) from eastern Indonesia, an association of secondary larger foraminifera was noted to correlate with plate tectonic divisions. A review of published data and proprietary industry records strengthened and extended this palaeogeographic correlation.
In order to partly test this hypothesis, a visit was made to Eocene limestones of Christmas Island. Christmas Island faunas occur between the two aforementioned faunas but had not yet been described in detail. The short accounts available mentioned neither of the palaeogeographic marker
taxa. The new samples showed that the apparent low diversity implied by previous summaries was real and not just a limitation of effort in study. The large number of polished and thin sections made for this study made it possible to identify that an abundant Heterostegine form was quite distinct and unusual, being a new species of the Boreal genus
Grzybowskia. This is the first record of this genus in the Indo-Pacific area or the southern hemisphere, and a summary of the distribution of this taxon also shows a strong palaeogeographic control. The reasons for including this species in
Grzybowskia, and hence for adding an additional palaeogeographic index taxa to the biogeographic reconstruction, are discussed below.
 A few reports of the APB fauna from the Australian plate in oil industry biostratigraphy studies (mostly from western Papua) have been investigated and, when original thin sections were still available, found to be incorrect. It should be noted that the APB
genera have some difficulties in identification, especially in random thin sections.
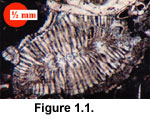
Assilina can be confused for flat forms of Operculina, and the usually distinctive
Pellatispira (Figure 1.1) has been confused with ornamented rotaliid forms, especially by industry biostratigraphers who are accustomed to examining planktonic
foraminifera. Lacazinella is strikingly unique and hard to mistake.
A few reports of the APB fauna from the Australian plate in oil industry biostratigraphy studies (mostly from western Papua) have been investigated and, when original thin sections were still available, found to be incorrect. It should be noted that the APB
genera have some difficulties in identification, especially in random thin sections.
Assilina can be confused for flat forms of Operculina, and the usually distinctive
Pellatispira (Figure 1.1) has been confused with ornamented rotaliid forms, especially by industry biostratigraphers who are accustomed to examining planktonic
foraminifera. Lacazinella is strikingly unique and hard to mistake.
 In the records discussed here no attempt has been made to distinguish
Assilina d'Orbigny 1839
from Planocamerinoides Cole 1957, in the sense of
Haynes (1988) and others, who place fully evolute forms (in the adult stages,
Figure 1.4) in
Assilina, and quasi-evolute forms (with spiral sheets extending over the previous whorls) in the latter genus (e.g.,
Figure 1.2 and Figure
1.3). Both taxa occur in the Indo-Pacific region, but most work was done during prior to the original or revised definitions of
Planocamerinoides.
In the records discussed here no attempt has been made to distinguish
Assilina d'Orbigny 1839
from Planocamerinoides Cole 1957, in the sense of
Haynes (1988) and others, who place fully evolute forms (in the adult stages,
Figure 1.4) in
Assilina, and quasi-evolute forms (with spiral sheets extending over the previous whorls) in the latter genus (e.g.,
Figure 1.2 and Figure
1.3). Both taxa occur in the Indo-Pacific region, but most work was done during prior to the original or revised definitions of
Planocamerinoides. 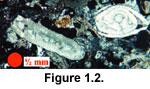 Not all records are illustrated, and therefore new taxonomies cannot be suggested.
Not all records are illustrated, and therefore new taxonomies cannot be suggested.
Environmental range of the genera
Both the APB and Lacazinella larger foraminifera usually occur as minor components in Eocene carbonates where the cosmopolitan genera
Discocyclina and Nummulites are dominant. This similar minority role suggests that the presence or absence of either of the markers is probably not related to gross lithologic or sedimentological effects. However, the genera
Assilina, Pellatispira, and Biplanispira usually occur only in a sub-set of samples from any particular Eocene locality, suggesting there is some local environmental control on its distribution.
 Lacazinella has a different local distribution
pattern, occuring commonly in low diversity assemblages (e.g., Figure
6), interbedded with more coralline limestones without
Lacazinella but containing Discocyclina and Nummulites. Because of this more stratified distribution, and the fact that it is a miliolid form,
Lacazinella has been interpreted as an indicator of back-reef facies. In my experience I have never observed more than a trace of planktonic foraminifera in the micritic limestone that hosts the most abundant
Lacazinella faunas. However, Lacazinella is reported from several locations on the Onin Peninsular area of west Papua
(Visser and Hermes 1962) and nearby Pisang Islands
(Rutten 1936), in a facies trend mapped by Visser and Hermes (enc. 10, part V) as "open marine, .. non reefal aphanitic limestones with predominance of planktonic
foraminifera." Also Apthorpe (1988) describes the widespread Hibernia limestone of the Australian Northwest shelf as being characterised by monospecific
Lacazinella assemblages - a wide distribution not typical for restricted back-reef
facies. Other supposed back reef miliolid indicators such as Alveolina are found in some Australian T.a stage (Middle Eocene) limestones but usually with
Nummulites and not with Lacazinella faunas. All this information suggests the
Lacazinella assemblages may have occupied a much more widely developed environmental setting than restricted back reefs habitats. More methodical observations are needed to clarify this hypothesis.
Lacazinella has a different local distribution
pattern, occuring commonly in low diversity assemblages (e.g., Figure
6), interbedded with more coralline limestones without
Lacazinella but containing Discocyclina and Nummulites. Because of this more stratified distribution, and the fact that it is a miliolid form,
Lacazinella has been interpreted as an indicator of back-reef facies. In my experience I have never observed more than a trace of planktonic foraminifera in the micritic limestone that hosts the most abundant
Lacazinella faunas. However, Lacazinella is reported from several locations on the Onin Peninsular area of west Papua
(Visser and Hermes 1962) and nearby Pisang Islands
(Rutten 1936), in a facies trend mapped by Visser and Hermes (enc. 10, part V) as "open marine, .. non reefal aphanitic limestones with predominance of planktonic
foraminifera." Also Apthorpe (1988) describes the widespread Hibernia limestone of the Australian Northwest shelf as being characterised by monospecific
Lacazinella assemblages - a wide distribution not typical for restricted back-reef
facies. Other supposed back reef miliolid indicators such as Alveolina are found in some Australian T.a stage (Middle Eocene) limestones but usually with
Nummulites and not with Lacazinella faunas. All this information suggests the
Lacazinella assemblages may have occupied a much more widely developed environmental setting than restricted back reefs habitats. More methodical observations are needed to clarify this hypothesis.
Stratigraphic range of the genera
 In the Indo-Pacific area Assilina is used to identify Middle Eocene carbonates, and a short overlap with
Pellatispira then occurs during latest Middle Eocene followed by Pellatispira and
Biplanispira in later Eocene carbonates (pers. obs., in contrast to Adams
1970). This biostratigraphy is one of the fundamental components in identifying Letter Stages T.a and T.b
(Figure 2; van der Vlerk and Umbgrove 1927 and later revisions).
In the Indo-Pacific area Assilina is used to identify Middle Eocene carbonates, and a short overlap with
Pellatispira then occurs during latest Middle Eocene followed by Pellatispira and
Biplanispira in later Eocene carbonates (pers. obs., in contrast to Adams
1970). This biostratigraphy is one of the fundamental components in identifying Letter Stages T.a and T.b
(Figure 2; van der Vlerk and Umbgrove 1927 and later revisions).
Dating Australian shallow marine carbonates using the Indo-Pacific Letter Stages is therefore difficult in the absence of the APB lineage.
Lacazinella is reported from beds considered as T.a (approximates Middle Eocene) on the basis of associated
Nummulites species such as N. bagelensis or large forms with meandrine septal traces
[Rutten 1936, Crespin 1962, and possibly
Bain and Binnekamp 1973]. The other T.a age index genus,
Alveolina, is found in both Sundaland and Australian regions but it is an uncommon form, suspected to be environmentally restricted (see notes above).
 Lacazinella continues through T.b (Late Eocene) and appears to share the same abrupt extinction as general Eocene forms such as
Discocyclina. However, Bursch (1947) described Lacazinella occurring with reticulate
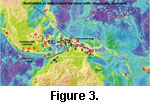
Nummulites in Kai Besar (Figure 3, location 3 on
map). Adams (1970) confirmed
Nummulites fichteli with Lacazinella in Bursch's material, which therefore appears to extend the range of the genus into Letter Stage
T.c, Early Oligocene.
Lacazinella continues through T.b (Late Eocene) and appears to share the same abrupt extinction as general Eocene forms such as
Discocyclina. However, Bursch (1947) described Lacazinella occurring with reticulate
Nummulites in Kai Besar (Figure 3, location 3 on
map). Adams (1970) confirmed
Nummulites fichteli with Lacazinella in Bursch's material, which therefore appears to extend the range of the genus into Letter Stage
T.c, Early Oligocene.
Lacazinella has been reported from the Late Paleocene of Oman by
White (1994), but the significance of this temporally and geographically isolated species in not known.
Geographic range of the genera
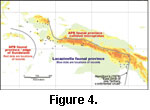 Figure
3, Figure 4, Appendix
1, and Appendix 2 illustrate the geographic spread of records of the genera.
Figure
3, Figure 4, Appendix
1, and Appendix 2 illustrate the geographic spread of records of the genera.
To summarise here I need only to stress that there is just a single known exception to the mutually exclusive distribution of the two faunas
(Crespin 1938, in the Chimbu Gorge of Papua New Guinea).
Even if this exception is evidence that the APB fauna could migrate in small numbers to the northern fringes of the high
palaeo-latitude Australian plate, it is a minor exception and unrelated to the total absence of
Lacazinella from low latitudes in the Eocene.
Both Boussac (1906)
and Umbgrove (1928) considered Pellatispira (which then still included
Biplanispira) and Assilina sensu lato as being "closely allied"
taxa, with the implication that the thick-walled assiline (Planocamerinid) forms could be both stratigraphically and morphologically
the ancestor to the pellatispirine types. Since then no missing link such as a pellatispirine species associated with an assiline juvenile stage has been found. Furthermore, the most recent work of
Hottinger et al. 1991 on pellatispirines has increased the morphologic differences between the two taxa by clearly showing the so-called marginal cord of
Pellatispira is a very different structure from the nummulitic cord (although some evolutionary link between the two features cannot be ruled out). The similar latitudinal association of the taxa presented here is weak data supporting the idea of a phylogenetic link between the two forms.
The work of Hottinger et al. 1991
clearly shows the close relationship of
Pellatispira and Biplanispira, as emphasised by the adult of the former often forming the juvenile stage of the latter.
