SYSTEMATIC PALEONTOLOGY
(continued)
Superfamily Giraffoidea Gray 1821
Family
?Giraffidae Gray 1821
Genus
Progiraffa Pilgrim 1908
Diagnosis
Moderate-sized pecoran (m2-m3 length: ~ 5.3 cm).
Ossicones present. Brachyodont teeth with rugose enamel. Lower molars with
prominent metastylid well separated from the metaconid, lingual face of
metaconid and entoconid convex, m3 with robust entoconulid filling the lingual
wall. “Palaeomeryx‑fold” absent. Upper molars with subsidiary spurs
separate from labially directed postprotocrista and premetaconule crista.
Postmetaconule crista with bifurcation. Low lingual cingulum on posterior of
protocone.
Progiraffa exigua
Pilgrim 1908
Diagnosis
As for the genus.
New Material
Locality Z 117: 2376, medial phalanx.
Locality Z 120: Z 593, fragment of a right p2 or
p3; Z 2276, left astragalus; Z 2392, right lower molar fragment.
Locality Z 121: Z 2274, right distal metatarsal.
Locality Z 124: Z 162, posterior portion of skull.
Locality Z 126: Z 2391, distal left humerus.
Locality Z 137: Z 202, right cuboid-navicular; Z
210, proximal left femur.
Locality Z 203: DGK 15, left astragalus.
Locality Z 205: DGK 23, right astragalus; DGK 45,
left proximal femur; DGK 148, fragment of an ossicone; Y 41662, left mandible
with incomplete m1 and m2.
Locality Z 211: DGK 145, left unciform; DGK 204,
astragalus; DGK 215, right m1 or m2; DGK 291, right magnum-trapezoid.
Locality Z 212: DGK 188, right astragalus; DGK
200, right distal metacarpal.
Locality H 8115: H 208, left m3; H 664, right
M3(?).
Locality H 8125; H 312, right maxilla with
erupting P3 and DP4-M3.
Locality S 2: S 88, very worn left m1 or m2; S
412, left p3.
Locality S 4: S 48, incomplete left m3; S 495,
fragment of a right m1 or m2.
Locality S 6: S 154, right m1 or m2.
Locality S 15: S 305, right mandible fragment with
m1 or m2.
Locality Y 591: Y 24013, broken left cuneiform; Y
24020, distal metapodial; Y 24021, fragment of a left P4; Y 31676, left m1 or
m2.
Locality Y 592: Y 17554, incomplete left
astragalus; Y 31204, fragment of a left calcaneum; Y 31206, right P4; Y 47224,
medial phalanx; Y 47225, right fibula.
Locality Y 652: Y 23429, left cuneiform.
Locality Y 687: Y 47366, proximal phalanx.
Locality Y 721: Y 26598, incomplete right
astragalus; Y 26599, fragments of a right calcaneum.
Locality Y 738: Y 31148, fragment of a very worn
left upper molar.
Locality Y 744: Y 40808, incomplete right
astragalus.
Locality Y 747: Y 31733, left cuboid-navicular; Y
31735, left unciform; Y 31736, left cuneiform; Y 31739, left cuneiform; Y 31743,
incomplete right astragalus; Y 31745, fragment of a right astragalus; Y 31748,
left astragalus; Y 31760, medial phalanx; Y 31764, proximal portion of a
proximal phalanx; Y 31766, medial phalanx; Y 31768, distal metapodial; Y 31771,
distal portion of a medial phalanx; Y 31772, distal fragment of a proximal
phalanx Y 31773, medial phalanx; Y 31774, medial phalanx; Y 31775, distal
metapodial; Y 31776, distal metacarpal; Y 31784, right distal femur; Y 31787,
proximal left tibial epiphysis; Y 31789, fragment of a proximal right tibial
epiphysis; Y 31794, fragment of a left distal radius; Y 31797, lower left p3; Y
41455, distal metapodial; Y 46276, distal metapodial; Y 46277, left cuneiform; Y
46284, fragment of a left astragalus; Y 47475, proximal portion of a proximal
phalanx; Y 47476, proximal phalanx; Y 47477, fragment of left m1 or m2; Y 47478,
distal portion of a proximal phalanx; Y 47479, distal metapodial epiphysis.
Locality Y 780: Y 32920, small fragment of a right
astragalus.
Locality Y 802: Y 46236, right fibula; Y 46237,
left lunar.
Locality Y 843: Y 41451, distal portion of a
proximal phalanx.
The presence of three left cuneiforms at Locality
Y 747 indicates there are at least three individuals at that site.
Localities and Age
Pilgrim’s type specimen is from the equivalent of
the Chitarwata Formation at Dera Bugti, but its age is not known with any
certainty. The Zinda Pir sites are in the upper unit of the Chitarwata Formation
and the overlying Vihowa Formation, with an age range of approximately 20 to 17
Ma (Lindsay et al. this issue). The Manchar sites (GSP-H and GSP-S) are in the
lower part of the sequence (Raza et al. 1984; Hussain, personal commun., 1986)
and are of Lower and Middle Miocene age. The oldest Potwar sites (GSP-Y 747 and
Y 721) are at the base of the terrestrial sequence in the Salt Range and are
estimated to be about 18.3 Ma, while the youngest (GSP-Y 591 and Y 592) are
estimated to be about 16.0 Ma (Johnson et al. 1985).
Ginsburg et al. (2001) have
reported additional specimens from level 6 of
Welcome et al. (2001) in the Chitarwata Formation at Dera Bugti, to which the latter authors assign an Early
Miocene age.
Description
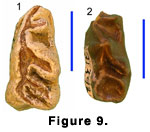 Lower Dentition. The available specimens
are all isolated premolars and molars, except for Y 41662, a mandible with
broken but otherwise well-preserved m1 and m2. The presumed p3’s (S 412 and
Y 31797) (Figure 9.1-9.2) are low and long, with faint labial incisions, a
large posterolingually directed metaconid that runs parallel to the equally
strong, oblique entoconid. The latter converges on a large, more transverse
entostylid to define a basin with a narrow lingual opening. The long anterior
crest is bifurcated, with a robust, lingually directed paraconid. In both
specimens there is a low cingulum just behind the base of the paraconid.
Lower Dentition. The available specimens
are all isolated premolars and molars, except for Y 41662, a mandible with
broken but otherwise well-preserved m1 and m2. The presumed p3’s (S 412 and
Y 31797) (Figure 9.1-9.2) are low and long, with faint labial incisions, a
large posterolingually directed metaconid that runs parallel to the equally
strong, oblique entoconid. The latter converges on a large, more transverse
entostylid to define a basin with a narrow lingual opening. The long anterior
crest is bifurcated, with a robust, lingually directed paraconid. In both
specimens there is a low cingulum just behind the base of the paraconid.
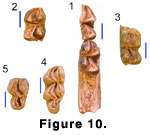 The lower molars (Figure 10.1-10.4) are brachyodont, with fine striations in the enamel. The obliquely situated
metaconid and entoconid have flattened labial faces but retain convex lingual
faces with weak development of lingual ribs in some cases. The premetacristid
contacts the anterolabial end of the preprotocristid, while the posthypocristid
reaches the labial side of the tooth, ending in a tubercle and nearly closing
off the posterolingual end of the median valley. In all specimens the
prehypocristid is separate from the preentocristid and trigonid cusps, even in
very advanced wear. None of the molars have even a vestige of a “Palaeomeryx‑fold,”
but all have large lingually displaced metastylids that are separated from the
metaconid by a strong vertical groove. The metastylid typically lies lingual to
the anterior end of the preentocristid. In two of the molars (S 305 and Y 31676)
(Figure 10.1-10.2) there is a small entostylid terminating the
postentocristid. This entostylid is distinct from the tubercle on the
posthypocristid. All the molars have low ectostylids and anterior and posterior
basal cingula.
The lower molars (Figure 10.1-10.4) are brachyodont, with fine striations in the enamel. The obliquely situated
metaconid and entoconid have flattened labial faces but retain convex lingual
faces with weak development of lingual ribs in some cases. The premetacristid
contacts the anterolabial end of the preprotocristid, while the posthypocristid
reaches the labial side of the tooth, ending in a tubercle and nearly closing
off the posterolingual end of the median valley. In all specimens the
prehypocristid is separate from the preentocristid and trigonid cusps, even in
very advanced wear. None of the molars have even a vestige of a “Palaeomeryx‑fold,”
but all have large lingually displaced metastylids that are separated from the
metaconid by a strong vertical groove. The metastylid typically lies lingual to
the anterior end of the preentocristid. In two of the molars (S 305 and Y 31676)
(Figure 10.1-10.2) there is a small entostylid terminating the
postentocristid. This entostylid is distinct from the tubercle on the
posthypocristid. All the molars have low ectostylids and anterior and posterior
basal cingula.
Both of the more complete m3’s (H 208 and S 48)
(Figure 10.3-10.4) have simple, crescent-shaped hypoconulids and a large
entoconulid that fills the space between the entoconid and hypoconulid, making
the lingual wall of the hypoconulid loop continuous. In S 48 the posthypocristid
contacts the entoconulid.
 Upper Dentition. H 312 (Figure 11.1) is a
well-preserved maxilla with undamaged crowns of all the molars, a worn dP4, and
P3 exposed but still in its crypt. An unerupted P4 is also present, but only the
forming roots are visible and the crown is hidden within the maxilla. P3 has a
tall, anteriorly situated paracone with a strong labial rib and foreshortened
anterior crest. The posterior crest of the paracone is, in contrast, elongated
and blade-like. A large parastyle is separated from the paracone by a deep
vertical groove. The anteriorly situated protocone is connected to the parastyle
by a low crest forming a convex lingual wall. Within the posterior fossette of
the median valley there is a complicated, low spur of enamel connecting to the
inner wall of the postprotocrista and running anteriorly.
Upper Dentition. H 312 (Figure 11.1) is a
well-preserved maxilla with undamaged crowns of all the molars, a worn dP4, and
P3 exposed but still in its crypt. An unerupted P4 is also present, but only the
forming roots are visible and the crown is hidden within the maxilla. P3 has a
tall, anteriorly situated paracone with a strong labial rib and foreshortened
anterior crest. The posterior crest of the paracone is, in contrast, elongated
and blade-like. A large parastyle is separated from the paracone by a deep
vertical groove. The anteriorly situated protocone is connected to the parastyle
by a low crest forming a convex lingual wall. Within the posterior fossette of
the median valley there is a complicated, low spur of enamel connecting to the
inner wall of the postprotocrista and running anteriorly.
The upper molars are low crowned, with large
metaconules on M1 and M2. On M3 the metaconule is smaller than the protocone.
The parastyles and mesostyles are prominent on all the molars, the paracones
have very strong labial ribs and the metacones flat labial faces. On the M1 the
postmetaconule crista has a distinct bifurcation, and there are faint traces of
this condition on M2 and M3. The three molars of H 312 have a complex of low
enamel spurs near the junction of the postprotocrista and premetaconule crista.
On M2 and M3 these appear as a bifurcation of the worn premetaconule crista, but
on M1 it has a more complex shape. These spurs clearly have formed as structures
separate from the postprotocone and premetaconule crista, although with wear
they appear to merge with the centrally directed crista. Both M2 and M3 have a
low cingulum on the posterior face of the protocone. This cingulum originates
between the protocone and metaconule. At the base of the metaconule there is an
entostyle-like structure.
H 664 (Figure 11.2) is judged to be an M3 because,
although in advanced wear, it has no posterior interdental facet. In addition,
the metaconule is smaller than the protocone, as is typical of an M3. The molar
has a bifurcated postmetaconule crista, but it does not have the small spurs,
entostyle, nor protocone cingulum seen in the other specimens.
 Cranial Remains. Ossicones: DGK 148 (Figure
12.1-12.3) is a small section of an ossicone (here used broadly to include the
branched structures of Climacoceras). The specimen is associated with a
mandible (Y 41662) and postcranials referred to Progiraffa exigua.
We assume, therefore, it also belongs to Progiraffa exigua. The ossicone
is possibly a midbeam segment, approximately 57.3 mm long. It is straight and
tapers toward the presumed distal end, with cross-section dimensions of 24.6 x
18.6 mm at the craniad end and 19.0 x 14.9 mm distally. In cross section it has
an oval shape, flattened on one side and with the greatest minor dimension
slightly off center. In cross section there is a gradual transition between a
very thin (3-6 mm at proximal end) outer zone of dense cortical bone and the
largely cancellenous interior. The surface has many fine grooves originating
from nutrient foramina at their proximal ends. These structures are scattered
uniformly about the surface and spiral counterclockwise about the long axis of
the bone when viewed from the distal end.
Cranial Remains. Ossicones: DGK 148 (Figure
12.1-12.3) is a small section of an ossicone (here used broadly to include the
branched structures of Climacoceras). The specimen is associated with a
mandible (Y 41662) and postcranials referred to Progiraffa exigua.
We assume, therefore, it also belongs to Progiraffa exigua. The ossicone
is possibly a midbeam segment, approximately 57.3 mm long. It is straight and
tapers toward the presumed distal end, with cross-section dimensions of 24.6 x
18.6 mm at the craniad end and 19.0 x 14.9 mm distally. In cross section it has
an oval shape, flattened on one side and with the greatest minor dimension
slightly off center. In cross section there is a gradual transition between a
very thin (3-6 mm at proximal end) outer zone of dense cortical bone and the
largely cancellenous interior. The surface has many fine grooves originating
from nutrient foramina at their proximal ends. These structures are scattered
uniformly about the surface and spiral counterclockwise about the long axis of
the bone when viewed from the distal end.
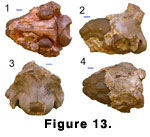 Skull: Z 162 (Figure 13.1-13.4) is the rear of a
broken skull. The specimen represents a medium-sized ruminant and, as
Progiraffa is the only known appropriate sized ruminant at this level, we
refer it to that species.
Skull: Z 162 (Figure 13.1-13.4) is the rear of a
broken skull. The specimen represents a medium-sized ruminant and, as
Progiraffa is the only known appropriate sized ruminant at this level, we
refer it to that species.
The specimen comprises the posterior of a damaged
cranium, lacking teeth and ossicones. Except for the nuchal crest and a small
remnant of the most dorsal part of the supraoccipital, the dorsal surface of the
cranium has been broken away, exposing a natural endocast of the brain. The
ventral surface has suffered only minor damage, while the supraoccipital region
is more or less intact. Although the frontals and skull roof have been
destroyed, the great thickness of the dorsal-lateral part of the lambdoidal
crest and orientation of the dorsal part of the supraoccipital suggest that the
skull roof was domed or that there may have been large posterior cranial
appendages. The supraoccipital is posteriorly projecting and laterally expanded
as it contacts the lambdoidal crest, with shallow pits for the semispinalis
capitis. Above the foramen magnum, the supraoccipital is slightly swollen, while
ventrally the paired occipital condyles are completely fused into a ring-like
structure, with the intercondyloid notch (incisura intercondyloidea of
Hamilton
[1973]) obliterated. The mastoid is narrowly exposed. On the ventral side of the
skull, the styloid process is posterolateral to and well separated from the
auditory bulla. The bulla is inflated and hollow, with an oval outline in
ventral view. It is well separated from the basioccipital and is not anteriorly
elongated. Consequently the middle lacerate foramen is left uncovered. The
basioccipital is posteriorly wide, narrowing anteriorly. It has well-developed,
laterally projecting condylar flexion stops (posterior tuberosities). A pair of
very subdued basilar tubercles is located posteriorly at about the level of the
posterior lacerate foramen, while anteriorly at the level of the foramen ovale
there is a second pair of very faint swellings. A median keel originates between
these latter swellings, running anteriorly onto the broken sphenoid. Anterior
and lateral to the posterior pair of basilar tubercles and just medial to the
auditory bulla there are paired irregular pits.
Postcranial Remains. Calcaneum: Y 31204 and
Y 26599 both preserve the fibular, cuboid, and much of the astragalar
articulations, and in the case of 26599 part of the calcaneal process. The
fibular articulation is typically pecoran, with a large convex posterior facet
and a smaller concave-convex anterior one. The bulbous posterior fibular
articulation has a medially projecting conical process with a distinct
posteromedial facet for the proximal lateral facet of the astragalus.
Fibula: Y 47225 is a distal fibula with a small
spine indicating it is a reduced remnant. Although damaged, the anterior tibial
facet is narrow and flat, and apparently did not curve medially.
Astragalus: The best preserved specimen is DGK
188, while all the others are broken or corroded by weathering. The
disto-lateral calcaneal facet is separated from the raised lateral edge of the
susentacular facet by a large sulcus, while the proximal lateral calcaneal facet
is confined to the edge of the susentacular facet and is separated from the
dorsal fibular facet. The cuboid condyle extends laterally, creating a distinct
notch on the lateral side of the astragalus, while the cuboid portion of the
condyle is cylindrical. The astragalus is narrow, with an approximate width to
length ratio of 1:1.7.
Cuboid-navicular: Z 202 is the more complete
specimen, but has been cracked and fractured, obscuring some of its features. Y
31723 is very well preserved, except for having lost the postero-ventral portion
of the astragalar articulation. In proximal view both are dorso-ventrally narrow
compared to the medial‑lateral diameter, with the cuboid‑astragalar facet being
slightly wider than the navicular‑astragalar one. The medial plantar process is
very prominent and the dorso‑medial angle of the navicular-astragalar
articulation is retracted. In distal view, the distal process does not extend to
the lateral margin of the anterior cuboid‑metatarsal facet and the relatively
broad posterior metatarsal articulation has a slightly inclined medial part
originating near the entocuneiform facet with a more horizontal lateral
extension. The facet for the 4th metatarsal lacks a posterior medial extension
bounding the groove for the tendon of the peroneus longus. Consequently the
peroneus groove is shallow and broad. The junction between the ectomesocuneiform
and endocuneiform articulations rises to form a low prominence, but both facets
are on the same level.
Lunar: The sole example (Y 46237) is too poorly
preserved to describe.
Cuneiform: With four well-preserved examples and a
fifth broken one, the cuneiform is the most common postcranial element. They
demonstrate slight morphological and size variation, but the stratigraphically
youngest (Y 23429) is typical in all respects. The unciform articulation is
relatively short and the disto-ventral process is directed ventrally rather than
being inflected distally. The pisiform articulation is narrow at its proximal
end, with its lateral margin being continuous rather than angled at the junction
with the ulnar articulation.
Magnum-trapezoid: The sole example (DGK 291) is
well preserved. Its dorsal ventral diameter is much larger than its
medio-lateral diameter, primarily because the lunar articulation is narrow and
not extended laterally. The junction between the lunar and scaphoid
articulations forms a high proximal keel. The inferior posterior unciform facet
is widely separated from the anterior one.
Unciform: The two referred specimens (DGK 145 and
Y 31735) differ somewhat in size and morphology, apparently reflecting
individual variation. DGK 145 is the smaller of the two. Its lunar articulation
has a flat, rectangular dorsal part that rises ventrally to form an almost
cylindrical condyle. The articulation for the cuneiform is relatively narrow,
without a marked lateral lip. The junction between the cuneiform and lunar
articulations is well differentiated, but low so that the two lie in nearly the
same horizontal plane. Y 31735 differs primarily in being about 10% larger and
in having a relatively wider posterior lunar articulation and a narrower one for
the cuneiform.
Metapodials: The fossils are all distal portions.
Three specimens (Z 2274, DGK 200, and Y 31776) preserve both the third and
fourth digits, while the remainder have only one side. Z 2274 is a metatarsal,
while DGK 200 and Y 31776 are metacarpals. The metatarsal gully appears to be
open in Z 2274, but the shaft is not preserved proximally enough to be certain.
In all specimens the keels of the distal articular surface extend dorsally but
do not project as far as they do ventrally. Consequently, in lateral view the
keels are strongly asymmetrical. The external condyle has a lipped rim which,
while stronger ventrally, also extends dorsally. The dorsal articular surface of
the external condyle is thus complete. Both the external and internal articular
surfaces terminate in a small pit on the dorsal side of the shaft.
Comparisons
The presence of ossicones leads us to tentatively
place Progiraffa in the Giraffidae.
Progiraffa exigua differs from middle
Miocene giraffids (sensu
Hamilton 1978) such as Giraffokeryx
punjabiensis Pilgrim 1911 and Injanatherium
Heintz, Brunet, and Sen
1981 in numerous ways (Colbert 1933;
Morales et al. 1987). These include the
presence in Progiraffa of a large, well-separated metastylid, a cingulum
on the protocone, and an enamel complex connecting the postprotocrista and
premetaconule crista. Progiraffa also seems to have a more sporadic
presence of a bifurcated metaconule and a more primitive basicranium with small
basilar tubercles and an oval auditory bulla that is well separated from the
basioccipital.
Progiraffa is similar to various primitive
giraffoids, such Propalaeoryx
Stromer 1926, Climacoceras
MacInnes
1936, Teruelia Moyà-Solà 1987, and Lorancameryx
Morales, Pickford,
and Soria 1993. The first of these, Propalaeoryx, is smaller and as far
as is known lacks cranial appendages. It also has a more anterior P3 protocone,
lacks entostyles on the molars, and the Namibian species P. austroafricanus
has a “Palaeomeryx‑fold” on the lower molars (Morales et al. 1999).
The ossicone described here for Progiraffa exigua could be a segment of
the branched, antler-like ossicone of Climacoceras, although it shows no
sign of secondary tines and its internal structure has a greater mass of
cancelleous bone and only a thin outer layer of dense cortical bone. The many
fine surface grooves and nutrient foramina on the Zinda Pir specimen are also
absent in Climacoceras africanus, although
Hamilton (1978) noted them in
C. gentryi. Dentally both species of Climacoceras differ from
Progiraffa. Their rather hypsodont lower molars have more compressed lingual
cusps with nearly parallel axes, metastylids that are not separated from the
metaconid, and m3 with the hypoconulid lingual wall incomplete. Climacoceras
also has a more anterior P3 protocone and narrower upper molars. Most of the
same dental characters differentiate Progiraffa from Lorancameryx
and Teruelia, the latter also lacking the bifurcated paraconid on p3.
The close similarity between Progiraffa and
Canthumeryx sirtensis
Hamilton 1973 (including Zarafa zelteni
Hamilton 1973) has been noted before (Moyà-Solà 1987,
Gentry 1994;
Ginsburg et
al. 2001). Canthumeryx is known principally from the dentition and
postcranials, to which a skull may be added if the synonymy of Zarafa is
accepted (Hamilton 1978, but see
Janis and Scott 1987). This taxon is near the
size of Progiraffa and nearly indistinguishable dentally. Possible points
of difference are a slightly stronger and more isolated metastylid in
Progiraffa and stronger bifurcation of the postmetacrista and greater
development of lingual basal structures in Canthumeryx (shown principally
in material originally attributed to Zarafa). The posterior and
basicrania of the skull of Progiraffa are also very similar to
Canthumeryx. Canthumeryx, however, has a flat skull roof, a large
swelling over the foramen magnum, an open intercondyloid notch, and it appears
to have a larger, more anterior pair of basilar tubercles. The slender ossicone
fragment (DGK 148) described here is also difficult to reconcile with the
massive bases seen in the referred skull of Canthumeryx, although
Churcher (1978; fig. 25.10) choose to reconstruct the specimen with slender,
spike-like ossicones.
Nyanzameryx pickfordi
Thomas 1984 and
Prolibytherium magnieri
Arambourg 1961 are two additional primitive giraffoids sharing some features of the skull and dentition with Progiraffa
(Geraads 1986). The isolated ossicone (DGK 148) is especially noteworthy, in
that it could be a segment of one of the slender ossicones of Nyanzameryx.
However, the nuchal area of the East African species is more gracile and less
projecting, while the basicrania has more anteriorly positioned basilar
tubercles. In addition, Nyanzameryx has lower molars with compressed
lingual cusps nearly parallel to the axis of the tooth, metastylids that are not
well separated from the metaconid, and m3 with the hypoconulid lingual wall
incomplete. It is also a much smaller taxon. Prolibytherium magnieri
shares one very distinctive feature with Progiraffa in that it has a
reduced intercondyloid notch, while the basilar tubercles are posteriorly
situated and very subdued. The great thickness of the skull roof of
Progiraffa suggests the presence of large and elaborate ossicones as in
Prolibytherium. The supraoccipital of Prolibytherium, however, does
not project so strongly posteriorly. There are also dental differences from
Progiraffa, principally in the possession of compressed lingual cusps that
are more parallel to the axis of the tooth, metastylids that are not well
separated from the metaconid, and an incomplete hypoconulid on m3.
Discussion
The initial description of Progiraffa was
very brief and included only one species (Pilgrim, 1908). In his subsequent,
more detailed account of Progiraffa
Pilgrim (1911) referred a second
species to the genus, one which
Lydekker (1883) had originally described as
Propaleomeryx sivalensis. Lydekker's single specimen is an isolated upper
molar, with few distinctive characters. It lacks the secondary complex of low
enamel spurs near the junction of the postprotocrista and premetaconule crista,
and is similar to the molar (H 664) from Sind. Except for its smaller size, the
latter specimen is virtually indistinguishable from Giraffokeryx punjabiensis.
The type specimen of Progiraffa exigua was until recently all that was
known of the taxon, but the new collections show the species is rather common.
The reference of both the ossicone (DGK 148) and
skull (Z 162) to one species is somewhat problematic. The construction of the
nuchal area of the skull (Z 162) suggests the cranial appendages were probably
robust and perhaps even similar to those of Prolibytherium. This
combination seems incompatible with the more spike-like ossicone DGK 148. The
skull fragment is about 19 Ma while the ossicone fragment is between 18 and 16
Ma (Lindsay et al. this issue). Both are from the Vihowa Formation.
The oldest specimen we attribute to Progiraffa
exigua is a poorly preserved distal humerus (Z 2391) from the upper unit
of the Chitarwata Formation, at a level we estimate to be 20 Ma. The specimen is
clearly a large pecoran, but might belong to the indeterminate large pecoran (Z
2031) from locality Z 127 rather than Progiraffa exigua. However,
it is appreciably larger than what we expect for a tooth the size of Z 2031 and
is an appropriate size for Progiraffa exigua.
On the Potwar Plateau Progiraffa exigua
persists up to at least 16 Ma. There are postcranial remains of a larger form
that is morphologically similar from sites between 16 and approximately 14 Ma.
While these might belong to another species of Progiraffa, we do not
describe nor discuss this larger form because it is only known from postcranial
elements. The cuneiform (Y 23429) from locality Y 652 is associated with a large
distal metapodial, which is likely to represent the larger pecoran, indicating
the two probably co-existed. After 14 Ma the still larger but morphologically
different remains of Giraffokeryx are found.
