SYSTEMATICS
Family DIATOMYIDAE Mein and Ginsburg, 1997
Genus WILLMUS, new
Diagnosis: Large diatomyid, most high crowned absolutely and relative to size, strongly bilophodont with lophs closely positioned and cusps suppressed, lophs nearly straight (except anterior loph of dp4), no accessory cuspules, no hint of posterior cingulum on dp4.
Type Species: (only known species): Willmus maximus, new species.
Etymology: Named for famed fossil hunter Will Downs, plus mus, Latin for mouse.
Willmus maximus, new species
Holotype: GSP 33100, right dp4.
Hypodigm: GSP 33100, plus GSP 36161, left M3.
Type Locality: (only known occurrence): Y797, lower part of Nagri Formation, Gabhir section of the Potwar Plateau, 11.1 Ma.
Diagnosis: As for genus, only member of the genus.
Etymology: maximus, Latin for greatest, invoking the greatest member of this mouse lineage, and the greatest mouse hunter, Will Downs, or, if you will, "mighty mouse."
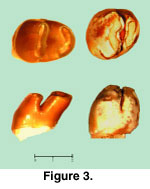 Description: Both specimens are strongly bilophodont and high crowned (Figure 3,
Figure 4). GSP 33100 is well preserved, moderately worn, and the enamel is light brown in color. GSP 36161 is less worn, but the enamel is corroded. It is possible, although improbable, that the two teeth represent the same individual. Many rodents, including rhizomyids, have delayed eruption of last molars such that anterior dentition is considerably worn before M3 erupts.
Description: Both specimens are strongly bilophodont and high crowned (Figure 3,
Figure 4). GSP 33100 is well preserved, moderately worn, and the enamel is light brown in color. GSP 36161 is less worn, but the enamel is corroded. It is possible, although improbable, that the two teeth represent the same individual. Many rodents, including rhizomyids, have delayed eruption of last molars such that anterior dentition is considerably worn before M3 erupts.
The lower premolar is narrow anteriorly, as is characteristic for the family (see below), with inclined lophs. The leading crest has a rounded V-shape derived from a linked protoconid-anteroconid-metaconid. The protoconid portion of the V is slightly thicker and the posterior cleft of the V is lingual to the midline of the tooth. The posterior loph is nearly straight and slightly oblique (entoconid end slightly anterior to hypoconid), and with gently invaginated anterior wall. There are no stylids or cingula. The valley between transverse lophs is equally open on each side, descending to about 0.9 mm above the base of the enamel on each side.
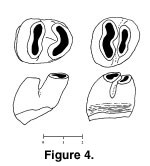 The base of the enamel undulates slightly. In lateral view, loph inclination is clear, and the occlusal wear planes on each loph slope anteriorly. Holding the tooth in anatomical position with the base of the enamel horizontal, the dimensions are 2.95 mm long and 2.33 mm wide (length would be slightly greater in an unworn tooth, due to the inclination of the crown). Root structure is unknown, but the base of the crown suggests two posterior roots. It is unknown whether GSP 33100 is p4 or dp4.
The base of the enamel undulates slightly. In lateral view, loph inclination is clear, and the occlusal wear planes on each loph slope anteriorly. Holding the tooth in anatomical position with the base of the enamel horizontal, the dimensions are 2.95 mm long and 2.33 mm wide (length would be slightly greater in an unworn tooth, due to the inclination of the crown). Root structure is unknown, but the base of the crown suggests two posterior roots. It is unknown whether GSP 33100 is p4 or dp4.
M3 appears slightly inclined posteriorly, with a vertical posterior loph and slanted anterior wall of the first loph. The base of the enamel undulates slightly and length equals width (2.43 mm). The first and larger loph has a gently convex anterior wall and posterior wall invaginated at the midline of the tooth. This loph represents the joined protocone-paracone of equal size. The second loph is a shorter figure-8 representing the smaller, fused hypocone-metacone. The base of the metacone appears to be larger than the base of the hypocone and is probably supported by an independent root. The hypocone is labial with respect to the protocone. The lophs are tightly appressed, but valleys at each end are open and rise to within 0.8 mm of the enamel-dentine junction. The two lophs in early attrition appear to share a common wear plane with some unevenness suggesting oblique power strokes; observed crown height, greater anteriorly, is 2.0 mm. At high magnification fine perikymata representing enamel prism decussation are visible on both sides of the M3, although they are more pronounced on the lingual side where the enamel surface is somewhat corroded.
Discussion: This unique taxon is distinguished by derived features of large size, considerable crown height, extreme bilophodonty, and absence of accessory cusps or cingula. It is derived with respect to
Diatomys from the late early and early middle Miocene of China, Thailand, and Pakistan. It is also derived with respect to the more bunodont
Fallomus and unnamed related Oligo-Miocene forms from Pakistan and India. These genera constitute the known record of this enigmatic Asian family of rodents.
Li (1974) described Diatomys shantungensis for a nearly complete, but deteriorated skeleton preserved in diatomites at Shanwang, a key reference locality for the early middle Miocene of China. In view of the transversely lophodont cheek teeth,
Li (1974) considered geomyoid and pedetoid affinity of this form but was not satisfied with familial identification. The skull was too poorly preserved to indicate higher taxon relationships; the teeth were clearly advanced in being quadrate, bilophodont, and multirooted.
D. shantungensis dentition is comparable in size to that of Willmus
maximus (see table in Li 1974).
The M3 (length, width = 2.65, 2.8 mm) is larger than that of Willmus, but
the premolar (length, width = 2.6, 1.9 mm) is smaller.
Flynn et al. (1986) named another bilophodont rodent, this one from the Bugti assemblage of Baluchistan, Pakistan.
Fallomus razae was so named, being mouse like in transverse arrangement of cusps, but obviously not a murine, being derived from a generalized ctenodactyloid dental pattern. Affinity with the more derived
Diatomys was recognized. Hypothesized membership of Fallomus in the ctenodactyloid Chapattimyidae led
Flynn et al. (1986) to support affinity to Pedetidae within Hystricognathi for these genera.
Mein and Ginsburg (1985) explicitly supported a pedetid relationship for diatomyids when they named
Diatomys liensis for material from the early Miocene of Thailand. They demonstrated diversity in the generic content of
Diatomys. They also noted that similar unnamed forms had been found in at least two sites in Pakistan (see
de Bruijn et al. 1981;
de Bruijn and Hussain 1984). Later,
Mein and Ginsburg (1985) described the variation present in a large sample of D. liensis and formally named Family Diatomyidae under Superfamily Ctenodactyloidea.
D. liensis was distinguished by smaller size and frequent presence of an ectostylid in m1 and by a reduced anterior cingulum on DP4 and posterior cingulum on dp4. This species, although smaller than
Willmus maximus, (M3 average length, width: 2.02, 2.18 mm; dp4: 2.00, 1.44) shows relative crown height rivaling that of
Willmus (height of unworn M3 about 1.5 mm).
By the end of the millennium, additional fossils known from various sites of the Indian subcontinent began to place
Fallomus in temporal context with Diatomys.
Kumar et al. (1996) found diverse and high crowned
Fallomus in the late Oligocene or early Miocene Kargil Formation, Ladakh, India, as well as a new rodent,
Wakkamys hartenbergeri. The latter, a primitive cricetid, tends to support Oligocene age of the locality. This fauna argues for considerable antiquity of
Fallomus predating Diatomys. Nanda and Sahni (1998) formally named the Kargil species as
Fallomus ladakhensis to emphasize its advanced nature over Fallomus razae.
Marivaux and Welcomme (2003) demonstrated Oligocene age of basal Bugti assemblages and recognized diversity in their large sample of
Fallomus from Paali, Bugti. The pre-Siwalik early Miocene Zinda Pir sequence (Downing et al. 1993) yielded undescribed material from several sites intermediate in morphology and age between
Fallomus and Diatomys. The Siwaliks of the Potwar Plateau also produced additional material of
Diatomys from locality Y747 near the base of the Kamlial Formation.
Evolution of the Diatomyidae on the Indian subcontinent involves suppression of the ctenodactyloid crown pattern by bilophodonty, through fusion of cusps, and increasing crown height. The bunodont ctenodactyloid pattern, with low ridges connecting cusps, is evident in
Fallomus. Fallomus has deciduous and permanent fourth premolars, as well as P3. Third molars are large, premolars are the smallest cheek teeth and primitively replaced, and lower teeth have a characteristic hypoconulid. In addition, upper molars (and P4 of
Fallomus) show an extra internal cusp, the entostyle, while lower teeth have an external ectostylid. The p4 and dp4 show three cusps in the anterior arcuate or V-shaped loph: the protoconid, anteroconid, and metaconid.
Diatomys joins opposite cusps in transverse lophs. Teeth are quadrate, and there is no indication of premolar replacement, meaning that the deciduous premolar is retained throughout life. Because
Willmus is derived with respect to Diatomys, we interpret its single premolar as deciduous. Hypoconulids are lost but evidenced in some specimens by a posterior cingulum. Crown height is somewhat increased. An intermediate stage is present in fossils from Zinda Pir locality Z113, and from the Murree Formation equivalent site HGSP 116 (de Bruijn et al. 1981). Lower premolars from each (Flynn 2000) show retention of the ectostylid and hypoconulid. Unpublished specimens from Z113 suggest that the entostyle is lost in upper teeth, but show two premolar morphologies, indicating replacement in this intermediate, pre-Diatomys form.
Willmus continues the trend of simplification. Lower premolars have a straight posterior loph and a V-shaped leading crest. No ectostylid or posterior cingulum is in evidence. The posterior wall of the tooth slants steeply and drops almost vertically near the base of the crown. The upper molar of
Willmus is simply bilophodont, with no indication of an entostyle. Both teeth are high crowned and large. Crown height appears to have been achieved to an equal extent on both the lingual and labial sides of the teeth; there is no unilateral development of hypsodonty. This interesting element of the Middle Siwalik fauna is the latest and most derived of its small group. A taxon of archaic origin, it persisted through the middle Miocene, evidently at low abundance in the Siwaliks.
