Discussion
AND CONCLUSIONS
Dentaries and teeth demonstrate the
presence of at least two sauropods in Malawi, Malawisaurus and
Karongasaurus. Based on differences in the morphology of the caudal
vertebrae that have been generally recognized as significant, titanosaurian
caudal vertebrae from Malawi also represent at least two taxa including
Malawisaurus and Titanosauridae indet. Malawisaurus has platycoelous
medial caudal vertebrae, whereas a procoelous middle or posterior caudal
vertebra is distinct from Malawisaurus and referred to Titanosauridae
indet. A third taxon may be represented by vertebra referred to Titanosauria
indet.
Phylogenetic analyses
that include Aeolosaurus, Alamosaurus, Andesaurus,
Antarctosaurus, Argentinosaurus, Epachthosaurus,
Malawisaurus, Nemegtosaurus, Neuquensaurus,
Opisthocoelicaudia, Quaesitosaurus, Rapetosaurus,
Saltasaurus, and Titanosaurus (Upchurch 1995,
1998,
1999;
Salgado et
al. 1997;
Curry Rogers and Forster 2001; see also
Wilson 2002) indicate that
Andesaurus and Malawisaurus are basal titanosaurians. In fact,
Malawisaurus is the most complete Early Cretaceous titanosaurian known. It
is represented by cranial elements, 18 cervical, 10 dorsal, six sacral, and 51
caudal vertebrae, 24 chevrons, pectoral elements, pelvic elements, and dermal
armor. Phylogenetic analyses also indicate that taxa with cylindrical teeth and
strongly procoelous posterior caudal vertebrae (or opisthocoelous in the case of
Opisthocoelicaudia) are more derived than those with broad teeth and
platycoelous middle and distal caudal vertebrae.
Skull Shape and
Morphological Diversity in Malawi Sauropods. Cranial material attributed to
titanosaurians includes one or two partial braincases of Titanosaurus
indicus from the Late Cretaceous of India (Berman and Jain 1982;
Chatterjee
and Rudra 1996), a maxilla from India (von Huene and Matley 1933), a partial
braincase and partial skull roof of Saltasaurus (PVL 4017-161) from the
Late Cretaceous of Patagonia (Powell 1986,
2003), a premaxilla of Titanosauridae
indet. from the Late Cretaceous of Patagonia (Scuitto and Martinez 1994), a premaxilla (PVL 3670-12) that
Powell (1979) identified as Laplatasaurus
but later referred to as Titanosauridae indet. (Powell 1986,
2003), a
fragmentary braincase from the Late Cretaceous of France (Le Loeuff et al.
1989), the braincase, quadrate, quadratojugal, squamosal, and the lower jaws of
Antarctosaurus wichmannianus (MACN 6904) from the Late Cretaceous
of Patagonia (von Huene 1929;
Powell 1986,
2003), two partial braincases of
Antarctosaurus septentrionalis from the Late Cretaceous of India (von Huene and Matley 1933;
Chatterjee
and Rudra 1996), a nearly complete
disarticulated cranium of Rapetosaurus from the Late Cretaceous of
Madagascar (Curry Rogers and Forster 2001,
2004), and the specimens of
Malawisaurus from the Early Cretaceous of Malawi.
There are two basic
morphs of sauropod skulls: one high and short, the other low and elongate
(Wilson 2002). The high, short morph is referred to as a macronarian skull,
whereas the low, elongate morph is generally referred to as a diplodocoid skull
(Coombs 1975;
McIntosh and Berman 1975;
Berman and McIntosh 1978;
Salgado and Calvo 1997;
Wilson 2002). The macronarian skull is present in Brachiosaurus,
Camarasaurus, Datousaurus, Euhelopus, Mamenchisaurus,
Omeisaurus, Shunosaurus, and most prosauropods, whereas the
diplodocoid skull is present in Apatosaurus, Diplodocus, and
dicraeosaurids (Salgado and Calvo 1997; see also
Tidwell and Carpenter, 2003).
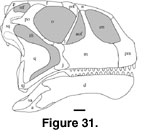 Comparison of cranial
features of Malawisaurus with macronarian and diplodocoid skulls
suggests that Malawisaurus had a high, short macronarian skull (Figure
31). The anterior section of the suture between premaxilla and maxilla appears
to have been nearly vertical as suggested by the highly angled articular surface
for the maxilla on the premaxilla. The high premaxilla also suggests that
Malawisaurus had a high, short, and blunt snout, and separate nares
positioned rostrally and facing laterally. The tooth row extends more than half
the length of the dentary. The mandibular symphysis is oblique to the long axis
of the mandible. The occipital condyle projected posteroventrally and the
basipterygoid processes projected ventrally. The quadrate axis was nearly
vertical, the pterygoid process of the quadrate was approximately perpendicular
to the long axis of the pterygoid, the pterygoid process was directed anteriorly
as in Brachiosaurus (Janensch 1935) and Camarasaurus (Osborn and Mook 1921), and the mandibular articulation was placed posteriorly beneath the
level of the occipital condyle. Thus, this study demonstrates that at least some
titanosaurians, including Malawisaurus, had high and short crania, as
compared to others such as Rapetosaurus, which had low and elongate
crania (Curry Rogers and Forster 2001,
2004).
Comparison of cranial
features of Malawisaurus with macronarian and diplodocoid skulls
suggests that Malawisaurus had a high, short macronarian skull (Figure
31). The anterior section of the suture between premaxilla and maxilla appears
to have been nearly vertical as suggested by the highly angled articular surface
for the maxilla on the premaxilla. The high premaxilla also suggests that
Malawisaurus had a high, short, and blunt snout, and separate nares
positioned rostrally and facing laterally. The tooth row extends more than half
the length of the dentary. The mandibular symphysis is oblique to the long axis
of the mandible. The occipital condyle projected posteroventrally and the
basipterygoid processes projected ventrally. The quadrate axis was nearly
vertical, the pterygoid process of the quadrate was approximately perpendicular
to the long axis of the pterygoid, the pterygoid process was directed anteriorly
as in Brachiosaurus (Janensch 1935) and Camarasaurus (Osborn and Mook 1921), and the mandibular articulation was placed posteriorly beneath the
level of the occipital condyle. Thus, this study demonstrates that at least some
titanosaurians, including Malawisaurus, had high and short crania, as
compared to others such as Rapetosaurus, which had low and elongate
crania (Curry Rogers and Forster 2001,
2004).
In addition, the titanosaurians
Nemegtosaurus (Nowinski 1971), and Quaesitosaurus (Kurzanov and
Bannikov 1983;
Curry Rogers and Forster 2001) had slender teeth restricted to
the anterior portion of the mandible, and also had low and elongate crania. If
that association of characters is general within titanosaurians, by implication
Karongasaurus would also have had a low and elongate cranium. Further,
phylogenetic analyses indicate that titanosaurians with slender teeth and which
have strongly procoelous middle and posterior caudal vertebrae are derived
relative to basal titanosaurians (Upchurch 1995;
Salgado et al. 1997;
Curry
Rogers and Forster 2001). Thus, both Karongasaurus and Titanosauridae
indet. from the Dinosaur Beds are derived relative to Malawisaurus. If
those characters are linked, the vertebra assigned to Titanosauridae indet. may
belong to Karongasaurus.
In any case, Malawisaurus and
Karongasaurus are two distinct titanosaurian taxa that coexisted in the
Early Cretaceous of Malawi and exhibited quite different morphological features,
certainly in their lower jaws and teeth, and probably also in their skull
shapes. Cylindrical teeth, an anteriorly restricted tooth row, and a long, low
skull shape evolved as a complex at least twice, once within diplodocoids and
once within titanosaurians. This character complex is functionally and
adaptively important for feeding. Its multiple origins, and its variance from
the macronarian skull pattern, implies that Malawisaurus and
Karongasaurus were ecologically distinct. If so, differences seen in the
lower jaw, teeth, and probably the skull of these herbivores were significant in
the ecological partitioning of their Early Cretaceous environment. Although the
macronarian and diplodocoid skull morphs are well known to occur together, for
example in the Jurassic of Africa, in the Early Cretaceous of Malawi
approximately equally divergent skull morphs are exhibited at a lower systematic
level among titanosaurians alone.
