The
two major environmental changes recorded in the Siwalik Group, the older
inception of the Asian monsoonal precipitation system and the younger transition
from C3 floras to C4 grasses, are broadly concurrent with the increase in
richness and evenness in the snake record, but precise correlations are
problematic. The timing of monsoonal inception is controversial: uplift of the
Tibetan Plateau drives the Asian monsoons, but the timing of the uplift is not
precisely constrained, with different lines of proxy data providing age
estimates from 7.0 to 8.0 Ma (Molnar et al. 1993) to 9.0 to 11.0 Ma (Kroon et
al. 1991). Oxygen-18 isotopic studies have demonstrated the onset of monsoonal
precipitation patterns in the Siwalik Group of Nepal beginning by 10.7 Ma
(Dettman et al. 2001), and
![]() 18O
values consistent with seasonal precipitation patterns began on the Potwar
Plateau at approximately 9.15 Ma (Quade and Cerling 1995 [dates refined in
Barry
et al. 2002]). Thus, the majority of data generally constrains the inception of
the monsoons to between 11.0 and 9.0 Ma, but does not unanimously provide a more
refined estimate.
18O
values consistent with seasonal precipitation patterns began on the Potwar
Plateau at approximately 9.15 Ma (Quade and Cerling 1995 [dates refined in
Barry
et al. 2002]). Thus, the majority of data generally constrains the inception of
the monsoons to between 11.0 and 9.0 Ma, but does not unanimously provide a more
refined estimate.
The
inferred monsoonal precipitation pattern is a progressive environmental event
that generally corresponds with the pattern of diversity increase in the Siwalik
snake record. However, temporal correlation is relatively weak within the
500-Kyr intervals. The beginning of diversity increase among Siwalik snakes at
10.2 Ma (Y-450) precedes consistent oxygen isotopic evidence for seasonality on
the Potwar Plateau. This may represent faunal response to increasing aridity
prior to the onset of seasonal evaporation in the western Siwalik Group.
Conversely, isolated 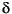 18O
values from the Potwar Plateau at 10.7 to 10.5 Ma are consistent with coeval
values indicating seasonal evaporation from Nepal (Dettman et al. 2001;
Barry
et al. 2002), and may represent an earlier increase in seasonality than previously
recorded.
18O
values from the Potwar Plateau at 10.7 to 10.5 Ma are consistent with coeval
values indicating seasonal evaporation from Nepal (Dettman et al. 2001;
Barry
et al. 2002), and may represent an earlier increase in seasonality than previously
recorded.
The transition from C3-dominated to C4-dominated vegetation is temporally well constrained at approximately 8.1 Ma, with fully C4 floras present by 7.4 Ma and the last occurrence of C3 floras at 7.0 Ma (Barry et al. 2002). This transition had a marked effect on mammalian feeding ecology (e.g., Flynn and Jacobs 1982) and is generally concurrent with increasing diversity in the snake record. More precise comparisons between the floral shift and the snake record are hampered by decreasing sample sizes from localities within the 6.5 Ma interval and younger, however.
Changes in richness and frequency in the Siwalik Group snake fauna are coeval with both increasing seasonality and the occurrence of the Dhok Pathan depositional environment, which are related and in turn ultimately driven by uplift histories of the Himalaya and Tibetan Plateau. Both events (and the subsequent evolution of C4 grasslands) would have increased the range of habitats within the area of the modern Potwar Plateau, which would have allowed an increasing diversity of taxa and habits within the Siwalik Group snake fauna.
The Siwalik Group is unique among Neogene fossil snake assemblages in its domination by a single taxon (compare with Szyndlar and Schleich 1993; Ivanov 2000) and in the absence of several taxa. There is no definitive occurrence of viperids within the Siwalik record, despite their extensive Neogene record throughout Europe (Szyndlar and Rage 1999), North America (Holman 2000), and other Asian localities (Rage and Ginsburg 1997; Ivanov 1999), and their presence in extant South Asian faunas (Minton 1966; Armed Forces Medical Intelligence Center, 1991). Their absence from the Siwalik section results from either taphonomic or ecological bias: the considerable sample of indeterminate colubroid specimens may include viperids, or viperids may not have actively inhabited the Siwalik environments proximal to depositional settings. Similarly, the absence of scolecophidians in the Siwalik Group is inconsistent with their presence in other Neogene strata (e.g., Szyndlar 1985, 1987, 1991). Scolecophidians are extremely small and fossorial to cryptic in habit. Their absence from the record likely reflects poor sampling of this habitat, an inference reinforced by the limited representation of similarly cryptic erycines in the Siwalik record, despite their abundance throughout the Neogene (Rage 1984; Holman 2000).
Extant European and South Asian snake faunas represent distinct biogeographic theaters (Boulenger 1913; Pope 1935; Minton 1966). The evolution of European snake faunas has been intensely studied (e.g., Ivanov 2000; Szyndlar and Rage 2003), but the Siwalik record is the only extensive fauna known for the Neogene of South Asia. Comparisons between the Siwalik fauna and approximately coeval faunas of Europe (Rage 1984; Szyndlar 1984, 1987; Ivanov 2000) demonstrate shared higher-order clades including Natricinae, Colubrinae, and possibly Erycinae, all of which were cosmopolitan by the end of the Paleogene (Rage 1984; Holman 2000). At more exclusive systematic levels, only a single genus, Python, is shared between Asia and Europe, a distribution that was achieved by the end of the Paleogene (Szyndlar and Rage 2003, this study). The rest of the Siwalik snake fauna is either endemic to the section or is singularly Asian. The fossil record of Acrochordus is only known from the Siwalik Group of South Asia and the Miocene of Thailand. Occurrence in Thailand indicates that the more eastern range of the genus was established early in its history, with physiographic barriers limiting the western distribution beyond Pakistan established by the onset of Siwalik sedimentation no later than 18 mya. The fossil record of Bungarus is also restricted to the Siwalik Group, which is geographically equivalent to the western margin of its extant distribution (Smith 1943; Minton 1966). That this very diagnostic taxon has not been recovered from the European record suggests that its past western distribution also did not extend beyond Pakistan, and the first occurrence of Bungarus, along with Chotaophis, Gansophis, and Sivaophis indicates that biogeographic barriers for derived snakes between South Asia and Europe were present by approximately 10.0 Ma.
The Siwalik snake record is different from the extant fauna of the Potwar Plateau. The modern snake fauna consists exclusively of colubroids, and only Bungarus is shared between the two intervals (western distributions of extant Python are just east of central Pakistan, Murphy and Henderson 1997). Acrochordus persists throughout the entire section, but A. dehmi is extinct, and there are no definitive historical records of extant species at the latitudes of Pakistan. Additionally, extant Acrochordus is not distributed as far inland as the Siwalik Group at any point in its current range. Extinction of A. dehmi on the Potwar Plateau is constrained as no older than 6.35 Ma and likely resulted from the draining of the Siwalik fluvial system off the plateau during the Pleistocene.
![]()