DISCUSSION
Coexisting fossil plants, vertebrates, and paleosols have seldom been reported from Paleogene sites in tropical Africa. At Chilga, the abundance of fossil assemblages, and their association with a range of paleosols, allow us to apply multiple proxies to reconstruct floral and faunal communities all within the context of their paleo-landscape.
The most striking characteristics of
paleosols in the Chilga sedimentary strata are gley coloration, or the
dominance of low chroma colors (e.g., gray or green), and accumulation of
organic material (Mack et al. 1993). These features are found within all of
the Chilga paleosols, but they are most prominent in paleosol Types A-C.
Such soils occur most commonly today in areas with a high water table, where
soil horizons are waterlogged and undergo anoxic, or aquic conditions for
most of the year (Mack et al. 1993;
Buol et al. 1997;
Soil Survey Staff
1998). Specifically, the combination of (1) gley colors, (2) diffuse
accumulation of organic material, (3) tabular rooting systems, (4)
accumulation of minerals associated with low redox potential, such as
siderite, and (5) lenticular bodies of organic-matter accumulation at the
surface of Type B and C paleosols, is interpreted to represent the poorest
soil drainage conditions across the Chilga floodplains, probably
corresponding to local ponding of water or very slow surface runoff. In
contrast, the presence of redoximorphic features, such as drab soil matrix
colors with red and orange color mottling and accumulation of fine hematitic
nodules in some of the Type C and D paleosols, suggests seasonal
groundwater-table fluctuation and alternating periods of reducing and
oxidizing conditions. Furthermore, the wedge-shaped aggregate structures and
slickenside formation in some of the Type C and all of the Type D paleosols
likely indicate periodic wetting and drying of the soil profile, suggesting
that these profiles formed upon portions of the landscape that experienced
periodic fluctuation of the local groundwater table.
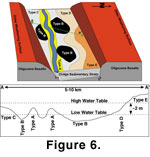 Finally, argillic
horizons in Type E paleosols require fairly stable, better-drained
conditions well above the groundwater table. Thus, we consider most of the
lateral and stratigraphic variation in the distribution of paleosol profiles
to record differences in the landscape position of soil-forming processes,
with respect to the paleo-groundwater table (Figure 6A-B).
Finally, argillic
horizons in Type E paleosols require fairly stable, better-drained
conditions well above the groundwater table. Thus, we consider most of the
lateral and stratigraphic variation in the distribution of paleosol profiles
to record differences in the landscape position of soil-forming processes,
with respect to the paleo-groundwater table (Figure 6A-B).
The composition and physiognomy of plant fossils from Chilga reflect local depositional conditions within the context of regional climate. The plants found in overbank or pond deposits close to a stream (in sediments associated with paleosol Type A) consist of taxa often found in those settings in tropical Africa today. These include palms, particularly calamoid or climbing palms, Sorindeia, and cf. Alismataceae. However, the entirety of this assemblage, including provisional identifications of Sterculiaceae, Fabaceae, and Dioscoreaceae, is indicative of a climate wet enough to support forest vegetation. Thus, a riparian association does not eliminate the necessity for ample rainfall, which must have been at least 1100 mm/year on average (the minimum needed to support forest vegetation today; e.g., Jacobs 1999).
Initial results from collection and study of plant specimens from the airfall tuffs (e.g., tuff IV) indicate that these plants grew in substrates that were very wet, perhaps swampy. The fern genus Acrostichum, found in ash IV in at least two locations across the basin, is represented today by three living species. These are pantropical and occur in either freshwater swamps or in association with mangrove vegetation (Mabberly 1993). Another pinnatifid fern occurs in this context at Chilga, associated with the leaves of dicots and legume fruits. Elsewhere, the same tuff preserves upright tree stumps, including palms, large palmate palm fronds, and legume leaves associated with pedogenic modification (Type A paleosols), a clear indication of a stratigraphic hiatus within the tuffaceous unit. In addition, tuff IV exhibits trough cross-bedding and sedimentary structures similar to those found in point bar deposits today, consistent with deposition of the airfall tuff in meandering stream channels. Thus, the airfall tuff bed, which is at least 7 m thick in most places, represents episodic ash deposition across a landscape characterized by a range of edaphic settings from swampy to subaerial.The fossil forests at Chilga are a good indication of wet conditions upon interfluvial settings. This inference is consistent with interpretations based on the plant assemblages known from overbank deposits, the presence of waterlogged bog or swamp deposits preserved as lignites, and the paleosol data, all of which point to poorly drained conditions in a generally wet climate.
With respect to mammalian assemblages, it is interesting to note that despite the somewhat more derived aspect of the individual taxa of the Chilga fauna, its general composition of arsinoitheres, hyracoids, and paleomastodonts largely matches that of the faunas from older Paleogene localities found around the continental margins of Afro-Arabia (Kappelman et al. 2003). The occurrence of this fauna in the highlands of Ethiopia suggests that these herbivores enjoyed a very widespread distribution across Afro-Arabia and probably occupied generalist niches with broad ecological tolerances. This observation is especially interesting in light of their overall low species diversities across a long temporal interval. In contrast, the dramatic diversification of the Proboscidea in the absence of any overall changes in faunal composition demonstrates that other, probably environmental, conditions were acting to drive their continued evolution. The question as to whether or not the arrival of Eurasian immigrants was responsible for driving the later extinction of the archaic endemics can only be tested by the recovery of new fossil localities dating closer to the Paleogene-Neogene boundary (23 Ma).
In summary, we envision an Oligocene landscape consisting of a generally flat and broad floodplain in a humid, tropical climate. Despite limited relief, landscape position played a primary role in the distribution of plants and in soil development due to variable but poor drainage conditions across the basin (Figure 6A-B).We hypothesize that stratigraphic changes in paleosol morphology and plant composition are primarily a reflection of intrabasinal physiography rather than extrabasinal controls, such as regional or global climate change. In addition, episodic ashfalls were literally superimposed upon these evolutionary geomorphic processes and provided a significant (perhaps primary) source of sedimentary material. The ash, depending on where it fell, was reworked to varying degrees, became part of a stable land surface upon which vegetation became established, or was incorporated into standing or flowing water.
Additional, quantitative paleoclimate proxies will provide a basis for evaluating the influence of climate on Chilga fossils and sedimentary characteristics and will allow us to test the hypothesis that climate change was not a primary driver of the physical and biotic changes identified in the sedimentary strata. Fossils and sediments have the potential to provide independent estimates of paleotemperature from the stable isotope composition of pedogenic minerals and fossil wood anatomical characters. Paleotemperature estimates from leaves at tropical sites have been shown to be unreliable (Jacobs 2002), but paleoprecipitation estimates can be derived from fossil leaf assemblages and from elemental composition of paleosols. To test for the occurrence of climate change, the accuracy of paleoclimate estimates derived from plant fossils will be evaluated by intrabasinal comparison of estimates from contemporaneous (stratigraphically equivalent) and facies-equivalent localities (Burnham 1994). If the plant fossils prove to be reliable climate proxies, then comparisons between facies-equivalent assemblages from different stratigraphic levels can assess the occurrence of climate changes through the sequence. Further analysis of plant fossil taxonomy will provide greater resolution of temporal and spatial variability of community composition, yield valuable biogeographic information, and provide historical documentation to complement and test systematic hypotheses generated by molecular phylogenetics.