SYSTEMATIC PALEONTOLOGY
THERAPSIDA
Broom, 1905
BIARMOSUCHIA Sigogneau-Russell,
1989
BURNETIAMORPHA Broom, 1923
Definition. The most
inclusive clade including Burnetia mirabilis, but excluding
Biarmosuchus tener, Hipposaurus boonstrai, and
Ictidorhinus martinsi (from
Sidor and Wellman 2003, p. 631).
Diagnosis. Supraorbital
boss present, antorbital fossa or pit on lateral surface of lacrimal; median
frontal ridge present but variably expressed; boss on ventral surface of
squamosal lateral to level of quadrate; boss present on lower margin of
zygomatic arch at level of postorbital bar (from
Sidor and Wellman 2003, p. 632).
Unnamed New Genus and Species
Material. A nearly
complete skull, slightly distorted and damaged in the premaxillary region.
Diagnosis. Mal 290 has a
skull roof that is relatively unpachyostotic; posteriorly directed squamosal
boss at dorsal apex of lateral temporal fenestra present, but not developed into
squamosal horns; narrow median nasal boss present. Its autapomorphies include:
short facial region, large orbit, and frontal and supraorbital bosses of equal
height. For other characters see phylogenetic analysis.
Occurrence. Lower Bone
Bed (B1) of the Chiweta Beds, Malawi.
Comments. Mal 290
exhibits all of the features of Burnetiamorpha listed above, although the
zygomatic boss is either poorly preserved or poorly developed. The description
and phylogenetic analysis demonstrates that Mal 290 is a new taxon, but we are
working from a cast in this study. As stated in the Introduction, Elizabeth M.
Gomani will oversee final description, analysis, and naming of this taxon in
conjunction with a more detailed study of the Chiweta Beds and its fauna.
Description of Mal 290
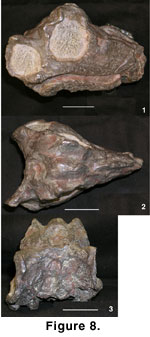 Mal 290 (Table 3,
Figure 8,
Figure 9) is nearly complete but slightly distorted and with the lower jaw
attached. The right side has been pushed posteriorly with the anterior portion
rotated ventromedially. The anterior premaxillary and nasal region is damaged so
incisor relationships are not obvious. The lower jaw is attached with the left
ramus in articulation. The right ramus is damaged and pushed posteriorly with
its posterior portion rotated medially more than the anterior portion. Sutures
are obscure, usually an indication that a specimen is not juvenile, but
burnetiamorph sutures tend to be difficult to discern and their obscurity may
not rule out Mal 290 being juvenile. Other features that might reflect a
juvenile state are the large size of the lateral temporal fenestra, large
orbits, and short facial region (Sidor personal commun., 2004).
Mal 290 (Table 3,
Figure 8,
Figure 9) is nearly complete but slightly distorted and with the lower jaw
attached. The right side has been pushed posteriorly with the anterior portion
rotated ventromedially. The anterior premaxillary and nasal region is damaged so
incisor relationships are not obvious. The lower jaw is attached with the left
ramus in articulation. The right ramus is damaged and pushed posteriorly with
its posterior portion rotated medially more than the anterior portion. Sutures
are obscure, usually an indication that a specimen is not juvenile, but
burnetiamorph sutures tend to be difficult to discern and their obscurity may
not rule out Mal 290 being juvenile. Other features that might reflect a
juvenile state are the large size of the lateral temporal fenestra, large
orbits, and short facial region (Sidor personal commun., 2004).
The broadest
transverse dimension of the skull is across the squamosals at the level of the
zygomatic arch at the ventral margin of the lateral temporal fenestra. The
posterior outline viewed from above is gently concave. The skull is narrow
across the frontals and tapers anteriorly. In lateral view, the skull is deep,
highest at the supraorbital bosses, but the lower jaw is shallow and not
expanded anteriorly to any great degree. The orbit is round and larger than the
lateral temporal fenestra, which is also nearly round and situated in the
posteroventral area of the skull. The narrow snout is short (length
approximately 120% of diameter of the orbit), but longer than the postorbital
length (approximately equal to orbital diameter). The occipital region is
vertical (perpendicular to the tooth row). The frontals contain the highest
point on the skull, from which the parietal region angles steeply to the dorsal
margin of the occipital region. The pineal foramen lies in a diffuse crater-like
swelling nearly in line with the posterior margin of the orbit and slightly
anterior to the plane of the occipital condyle. Anteriorly the skull slopes less
steeply toward the snout.
 There is minor
anteroposterior shearing deformation of the supraorbital bosses relative to the
frontal boss and minor dorsal extension of the left supraorbital boss relative
to the right. Assuming all deformation is recognized, the supraorbital bosses
are well developed and of comparable height to the broad median frontal boss.
The frontal boss is widely separated from a distinct, narrow, median nasal boss,
which extends toward the snout from the plane of the anterior margin of the
orbit. It is highest posteriorly, tapering gradually to the nose, terminating at
the posterior margin of the external nares. A boss occurs at the posterodorsal
margin of the orbit near the origin of the postorbital bar. A well-developed
boss is found on the squamosal at the dorsal margin of the lateral temporal
fenestra but it is not developed into a squamosal horn. A poorly preserved or
weakly developed boss occurs on the zygomatic arch at the level of the
postorbital bar. The zygomatic arch is bent strongly ventrally.
There is minor
anteroposterior shearing deformation of the supraorbital bosses relative to the
frontal boss and minor dorsal extension of the left supraorbital boss relative
to the right. Assuming all deformation is recognized, the supraorbital bosses
are well developed and of comparable height to the broad median frontal boss.
The frontal boss is widely separated from a distinct, narrow, median nasal boss,
which extends toward the snout from the plane of the anterior margin of the
orbit. It is highest posteriorly, tapering gradually to the nose, terminating at
the posterior margin of the external nares. A boss occurs at the posterodorsal
margin of the orbit near the origin of the postorbital bar. A well-developed
boss is found on the squamosal at the dorsal margin of the lateral temporal
fenestra but it is not developed into a squamosal horn. A poorly preserved or
weakly developed boss occurs on the zygomatic arch at the level of the
postorbital bar. The zygomatic arch is bent strongly ventrally.
The dorsal process
of the premaxilla is short and does not extend to a level posterior to the
canine as indicated by the narrowness of the snout. The septomaxilla is exposed
on the face. Fossae are present in the position of the lacrimal, although
sutures are difficult to discern. The posterior extent of the postorbital onto
the margin of the lateral temporal fenestra is unclear, but it does not appear
to extend to the posterior margin of the lateral temporal fenestra because of
the squamosal boss developed along the posterodorsal margin. The medial portion
of the squamosal overlaps the quadrate.
The anterior
portion of the palate has not been prepared so the vomer and the vomerine
process of the premaxillae cannot be seen, nor can the anterior portion of the
palatines. The ectoptygoids are not visible. The transverse flanges of the
pterygoids lie beneath the anterior half of the orbit and appear to have
denticles. The basicranial rami of the pterygoids exhibit a median trough
defined by parasagittal ridges. The parabasisphenoid has a ventral fossa and
apparently lacks a midline ridge. It extends back to the basioccipital at the
occipital condyle.
The occipital
condyle is small. The relationship of the exoccipitals on the dorsal side of the
foramen magnum is unclear. A ridge extends dorsally from the foramen magnum to
the dorsal margin of the postparietal. Given the width of the posterior portion
of the skull between the tabulars, the postparietal appears to be approximately
square. A posttemporal fenestra lies dorsal to the paroccipital process. The
stapes is not visible.
The mandibular
symphysis is unfused. The level of the base of the dentary follows a sinuous
curve, but the bone is relatively shallow. The shape of the dentary-angular
suture is unclear. Ridges and fossae occur on the angular. The articular is
unprepared. There is no fenestra between angular and dentary. The surangular has
a laterally projecting ridge. The jaw articulation lies below the level of the
tooth row. The medial surface of the jaw is unprepared.
The upper incisors
are not preserved, but they could not have been as large as the canine. At least
six postcanine teeth are visible on the left side of the skull, although
serrations cannot be discerned. The area around the left upper canine is
slightly crushed, but the remnants of the right canine suggest a slight anterior
orientation and a short diastema between the canine and postcanine dentition.
The lower canine is unprepared, and no dentary teeth can be seen.
Phylogenetic Position of Mal 290
Phylogenetic
analysis was performed using PAUP 4.0 Beta 10 and MacClade 4.06 on the data
matrix of 37 characters presented by
Sidor and Welman (2003), with character 30
(of Sidor and Welman 2003; ratio of dentary height in canine versus anterior
postcanine regions) deleted because we found the scoring confusing, and with Mal
290 added to bring the total number of taxa to 13 (Appendix,
Table 4). We
chose this data set because it includes taxa in which we are most interested,
notably Lemurosaurus and the burnetiids Proburnetia, Burnetia,
and Bullacephalus. This group, the Burnetiamorpha, falls within the
Biarmosuchia, originally considered to be defined on primitive characters but
now accepted as monophyletic (Hopson 1991;
Hopson and Barghusen 1986;
Rubidge
and Kitching 2003;
Rubidge and Sidor 2001,
2002;
Sidor 2001,
2003;
Sidor and
Hopson 1998;
Sidor and Welman 2003;
Sigogneau-Russell 1989). The sphenacodontid
synapsids Haptodus and Dimetrodon were specified as outgroups, as
in Sidor and Welman (2003). In most cases, the operational taxonomic units in
the data matrix are genera, except for the family level group Anteosauridae and
the single, taxonomically undesignated specimen Mal 290. Multistate characters
were treated as unordered. All characters were equally weighted. Unknown or
inapplicable characters were scored with a question mark. Branch and bound
searches found three most parsimonious trees with lengths of 55, consistency
indices of 0.82, and retention indices of 0.86.
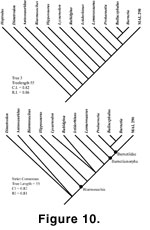 All three most parsimonious
cladograms and the strict consensus tree (Figure 10) exhibit identical
topologies among Ictidorhinus, Lemurosaurus, Mal 290,
Proburnetia, Bullacephalus, and Burnetia. Mal 290 forms a
monophyletic clade with the Burnetiidae, which includes Burnetia,
Proburnetia, and Bullacephalus. Mal 290 is united with the
Burnetiidae by two unambiguous characters, (Appendix, character 6) nasal
eminence (although the nasal eminence in Proburnetia is not like
Burnetia nor Bullacephalus) and (character 16) circum-pineal parietal
shape (which might be considered distinct in Bullacephalus), in two most
parsimonious trees. The burnetiids, exclusive of Mal. 290, are defined by
(character 10) thickening of the squamosal lateral to quadrate, (character 12)
squamosal horns present, and (character 13) squamosal thickened along its
posterior border with tabular. Within the Burnetiidae, Burnetia and
Bullacephalus consistently fall as a derived sister group to Proburnetia,
as was concluded by
Sidor and Welman (2003), because the nasal boss is scored as
transversely expanded (character 6).
All three most parsimonious
cladograms and the strict consensus tree (Figure 10) exhibit identical
topologies among Ictidorhinus, Lemurosaurus, Mal 290,
Proburnetia, Bullacephalus, and Burnetia. Mal 290 forms a
monophyletic clade with the Burnetiidae, which includes Burnetia,
Proburnetia, and Bullacephalus. Mal 290 is united with the
Burnetiidae by two unambiguous characters, (Appendix, character 6) nasal
eminence (although the nasal eminence in Proburnetia is not like
Burnetia nor Bullacephalus) and (character 16) circum-pineal parietal
shape (which might be considered distinct in Bullacephalus), in two most
parsimonious trees. The burnetiids, exclusive of Mal. 290, are defined by
(character 10) thickening of the squamosal lateral to quadrate, (character 12)
squamosal horns present, and (character 13) squamosal thickened along its
posterior border with tabular. Within the Burnetiidae, Burnetia and
Bullacephalus consistently fall as a derived sister group to Proburnetia,
as was concluded by
Sidor and Welman (2003), because the nasal boss is scored as
transversely expanded (character 6).
The definition of Burnetiamorpha
cited by Sidor and Welman (2003, p. 631) is, “The most inclusive clade including
Burnetia mirabilis, but excluding Biarmosuchus tener,
Hipposaurus boonstrai, and Ictidorhinus martinsi.”
Ictidorhinus, shown as the next taxon out from Burnetiamopha, is a poorly
known taxon with 57% of its characters scored as question mark. It is weakly
linked to Burnetiamorpha. The more basal portion of the tree follows
Sidor and Welman (2003) and needs not be discussed here.
