SYSTEMATIC PALEONTOLOGY
Order LAGOMORPHA
Brandt, 1855
Family LEPORIDAE
Fischer de Waldheim, 1817
Genus
GOBIOLAGUS Burke, 1941
Type Species:
Gobiolagus tolmachovi
Burke, 1941.
Emended
Diagnosis:
Cheek tooth crown higher than that of Shamolagus but lower than that of
Desmatolagus except for D. vetustus; p3 and m3 are proportionally
smaller than those of Shamolagus but larger than those of Desmatolagus;
incisor extending to m1 or m2; lower cheek tooth trigonid
transversely extended; talonid shorter than trigonid except for m2; enamel
ridge connecting the trigonid and talonid at the lingual side; p4 significantly
smaller than m1; p4 talonid reduced and trigonid anteroposteriorly elongated; m2
the largest cheek teeth; m2 talonid much lower than the trigonid in contrast to
the condition in p4 or m1; P4 nonmolariform and significantly smaller than M1;
P3-4 not lingually extended; M3 reduction moderate.
Included
Species:
Gobiolagus andrewsi, G. major, G. lii, and G. burkei sp. nov.
Comments:
Gobiolagus is a leporid genus proposed by
Burke (1941) based on specimens
collected from central Asia, although the genus was
recently placed under Leporidae with a query (Zhang et al. 2001).
Compared to other early Tertiary lagomorphs, such as Desmatolagus,
specimens of Gobiolagus are relatively rare. Since
Burke (1941), possible
Gobiolagus specimens have been reported from several localities of Asia
(Li 1965;
Qi 1988;
Tong 1997;
Erbajeva 1999;
Meng et al. 1999;
Zhang et al.
2001), all represented by fragmentary specimens. A total of four species were
named at the time the genus was established: G. tolmachovi, G. andrewsi, G.
(?) major, and G. (?) teilhardi. All the type specimens
are partial lower jaws, and those of G. (?) major and G.
(?) teilhardi are particularly poor. Thus, these two species were named
with uncertainty. One of the species, G. (?) teilhardi, was later
assigned to a different genus, Ordolagus, because of its high crown cheek
teeth in which the tooth crown extends into the alveolus (de Muizon 1977;
Huang
1986). Ordolagus teilhardi is an early Oligocene species from the Hsanda
Gol Formation of Mongolia. Specimens that are identified with certainty to
Gobiolagus are mostly from the late Eocene Shara Murun Formation and Ulan
Gauche Formation of the Shara Murun region, Inner Mongolia, except a few
isolated teeth assigned to G. cf. major from the Oligocene of
Kazakhstan (Erbajeva 1999). Isolated teeth assigned to Gobiolagus are
reported from the Sharamurunian beds of Henna, south China (Tong 1997) and of
Inner Mongolia (Li 1965;
Meng et al. 1999), and from the upper Oligocene of Zaisan Depression, Kazakhstan (Erbajeva 1999), and the lower Oligocene Valley of
Lakes, Mongolia (Erbajeva and Daxner-Hoeck 2001).
Gobiolagus lii was reported recently from
the middle Eocene Yuli member of the Heti Formation, Yuanqu, Shanxi Province,
China (Zhang et al. 2001).
Gobiolagus
tolmachovi
Burke, 1941
Emended
Diagnosis: Smaller than
G. major but larger than G. lii and G. burkei sp. nov.; p3
internal reentrant at the anterolingual side of the tooth; p4 trigonid
pear-shaped.
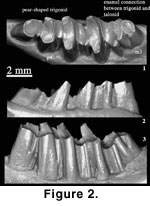
 Locality and
Age: The holotype (AMNH
26142, Figure 1.1,
Figure 2) and a referred specimen (AMNH 26143) are from the late
middle Eocene Shara Murun Formation near Baron Sog, Inner Mongolia (Burke 1941).
Additional specimens described here are from the Shara Murun Formation, Ula Usu,
Inner Mongolia.
Locality and
Age: The holotype (AMNH
26142, Figure 1.1,
Figure 2) and a referred specimen (AMNH 26143) are from the late
middle Eocene Shara Murun Formation near Baron Sog, Inner Mongolia (Burke 1941).
Additional specimens described here are from the Shara Murun Formation, Ula Usu,
Inner Mongolia.
 Referred
specimens: AMNH 141277,
right maxilla with P4-M2, alveoli of P2-3 and partial alveolus of M3; AMNH
141278, right maxilla with P4-M2 of a relatively young individual; AMNH 141279,
left maxilla with broken P4-M2; AMNH 141280, left maxilla with broken P4-M2;
AMNH 141281, right maxilla with P3-4 of a relatively young individual; AMNH
141282, right maxilla with P4 and broken M1-2; AMNH 141283, left maxilla with
P3-M1, alveolus of p2 and partial alveolus of M2; AMNH 141284, left maxilla with
broken P4-M2; AMNH 141285, left maxilla with broken P3-M2; AMNH 141286, left
maxilla with broken P4 and broken M1-2; AMNH 141287, left maxilla with M1-2;
AMNH 141288, left mandible with p3-m1; AMNH 141289, right mandible with broken
p4-m1; AMNH 141290, right mandible with broken p4-m2; AMNH 141291, left mandible
with m1-3; AMNH 141292, right mandible with p4-m1; AMNH 141293, right mandible
with broken m2; AMNH 141294, left mandible with m1; AMNH 141295, left mandible
with p4-m2. The referred specimens were collected in 1925 and were associated
with the field number 507 and catalog number AMNH 21750.
Referred
specimens: AMNH 141277,
right maxilla with P4-M2, alveoli of P2-3 and partial alveolus of M3; AMNH
141278, right maxilla with P4-M2 of a relatively young individual; AMNH 141279,
left maxilla with broken P4-M2; AMNH 141280, left maxilla with broken P4-M2;
AMNH 141281, right maxilla with P3-4 of a relatively young individual; AMNH
141282, right maxilla with P4 and broken M1-2; AMNH 141283, left maxilla with
P3-M1, alveolus of p2 and partial alveolus of M2; AMNH 141284, left maxilla with
broken P4-M2; AMNH 141285, left maxilla with broken P3-M2; AMNH 141286, left
maxilla with broken P4 and broken M1-2; AMNH 141287, left maxilla with M1-2;
AMNH 141288, left mandible with p3-m1; AMNH 141289, right mandible with broken
p4-m1; AMNH 141290, right mandible with broken p4-m2; AMNH 141291, left mandible
with m1-3; AMNH 141292, right mandible with p4-m1; AMNH 141293, right mandible
with broken m2; AMNH 141294, left mandible with m1; AMNH 141295, left mandible
with p4-m2. The referred specimens were collected in 1925 and were associated
with the field number 507 and catalog number AMNH 21750.
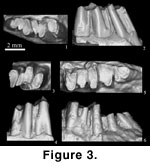 Description:
Measurements of the teeth are given in Table 1. The p3
is preserved in AMNH 141288 (Figure 3.1-2). The tooth is less worn than
in the holotype of G. tolmachovi (Figure 2) so that the lingual and
labial reentrants in the new specimen are deeper. However, because of relatively
smaller size of AMNH 141288 (Table 1), the absolute height of the tooth is lower
than that of the holotype. In occlusal view, the tooth is narrower anteriorly
than posteriorly. The lingual reentrant is at the anterolingual side of the
tooth crown and is about one third of the tooth height. The hypostriid (labial
reentrant) is much more distinct and over half of the tooth height.
Description:
Measurements of the teeth are given in Table 1. The p3
is preserved in AMNH 141288 (Figure 3.1-2). The tooth is less worn than
in the holotype of G. tolmachovi (Figure 2) so that the lingual and
labial reentrants in the new specimen are deeper. However, because of relatively
smaller size of AMNH 141288 (Table 1), the absolute height of the tooth is lower
than that of the holotype. In occlusal view, the tooth is narrower anteriorly
than posteriorly. The lingual reentrant is at the anterolingual side of the
tooth crown and is about one third of the tooth height. The hypostriid (labial
reentrant) is much more distinct and over half of the tooth height.
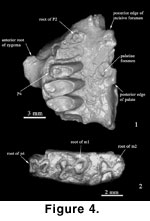 In labial or lingual view, the
cheek teeth appear to have two separate roots, but broken specimens (AMNH
141295; Figure 4.2) show that the roots in all cheek teeth are fused. The part
of the root supporting the trigonid is wider and is formed by fusion of two
smaller roots. The part of the root supporting the talonid is narrower but
longer and is in a circular contour in cross sectional view. The hypostriid and
the lingual reentrant extend continuously to the end of the fused root on the
labial and lingual sides of the tooth.
In labial or lingual view, the
cheek teeth appear to have two separate roots, but broken specimens (AMNH
141295; Figure 4.2) show that the roots in all cheek teeth are fused. The part
of the root supporting the trigonid is wider and is formed by fusion of two
smaller roots. The part of the root supporting the talonid is narrower but
longer and is in a circular contour in cross sectional view. The hypostriid and
the lingual reentrant extend continuously to the end of the fused root on the
labial and lingual sides of the tooth.
The p4 is
molariform but much smaller than m1 (Figure 3.1-4). The p4 trigonid is
pear-shaped, with its lateral side being wider and rounded. The talonid is short
and, in contrast to the trigonid, the lingual side is wider than the labial. The
trigonid and talonid are not connected at the lingual side, which differs from
the condition of the molars.
The m1 is
significantly larger than p4 (Figure 3.1-6). The m1 trigonid is transversely
wide. The talonid is shorter than the trigonid but is much larger than that of
p4. The two lobes are connected at the lingual side by a narrow band of enamel.
The m2 is the largest lower cheek tooth. Its talonid is longer than the trigonid
and is proportionally more expanded than in p4 and m1. The hypostriid reaches
the edge of the alveolus and is much wider than that of m1. The posterolateral
corner of the talonid is squared and the posterolingual corner is rounded. The
m3 trigonid is short and is also wider labially than lingually. The talonid is
oval-shaped and much longer than the trigonid. As in m1 and m2, the two lobes
are connected by the enamel ridge at the lingual side.
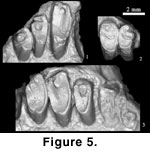 The maxilla
(Figure 4.1) and upper teeth (Figure 5.1-3) are associated with the mandible and
lower teeth because of their similar sizes and due to the distinctive size
increment between P4 and M1, which is consistent with that of the lower
dentition. A palatine foramen is completely within the palatine. There is no
premolar foramen. The posterior border of the incisive foramen levels with the
anterior edge of the alveolus for P2. The posterior edge of the palate is at the
level of M1. The anterior root of the zygomatic arch bears a ventral process. An
antorbital fossa is present anteromedial to the process or lateral to P3-4. The
posterior edge of the anterior zygomatic root is lateral to the anterior half of
M1. The external outline of the maxilla is not so convex as in the maxilla
assigned to G. tolmachovi by Qi (1988) and in G. lii (Zhang et al.
2001).
The maxilla
(Figure 4.1) and upper teeth (Figure 5.1-3) are associated with the mandible and
lower teeth because of their similar sizes and due to the distinctive size
increment between P4 and M1, which is consistent with that of the lower
dentition. A palatine foramen is completely within the palatine. There is no
premolar foramen. The posterior border of the incisive foramen levels with the
anterior edge of the alveolus for P2. The posterior edge of the palate is at the
level of M1. The anterior root of the zygomatic arch bears a ventral process. An
antorbital fossa is present anteromedial to the process or lateral to P3-4. The
posterior edge of the anterior zygomatic root is lateral to the anterior half of
M1. The external outline of the maxilla is not so convex as in the maxilla
assigned to G. tolmachovi by Qi (1988) and in G. lii (Zhang et al.
2001).
P2 is not
preserved, but a single rooted, triangular tooth can be estimated from the
alveolus (Figure 4.1,
Figure 5.3). P3 is trilobate and oval-shaped in occlusal view
(Figure 5.1-2). It has two roots, located lingually and labially. Its lingual
side is higher than the labial, but the difference is not so much as in P4. The
two valleys (reentrants) are simple. The labial valley is shallow and wide and
opens anterolabially. The lingual valley is narrow and extends posterolabially
around the central cusp; this condition is better seen in a worn specimen
(Figure 5.2). The lingual cusp, presumably the protocone, has a squared lingual
outline and bears no hypostria. As shown in AMNH 141283 (Figure 5.1), this cusp
is worn earlier than the central cusp in ontogeny. The central cusp is circular
in outline except at its posterolabial side where it connects to the rest of the
tooth. The posteroloph extends from the lingual cusp to the labial one. The
lingual arch of the tooth is not great; therefore, with increasing wear, the
transverse extension of the occlusal surface of the tooth is limited.
P4 is also
two-rooted, nonmolariform and with an oval-shaped occlusal view (Figure 5.1-2).
P4 is wider than P3, and its lingual side is much more hypsodont than P3. In a
worn specimen, such as AMNH 141286, P4 can be as wide as, or wider than the
molars. The anteroloph is complete so that the valleys on P4 become two enamel
fossettes that bound the central cusp lingually and labially, respectively. The
labial fossette is much shallower than the lingual one and bears very thin
enamel. The lingual fossette, or the crescentic valley, is curved and is framed
with a thick enamel wall labially and a thin wall lingually. The labial cusp is
small and does not show any sign of division.
M1 is
significantly larger than P4 (Figure 5). It has two small roots on the labial
side. The hypostria and crescentic valley are absent in all specimens. The crown
surface is characterized by an oval enamel contour on the occlusal surface. The
enamel along the anterior edge of the tooth is much thicker than that along the
posterior edge. The labial side of the tooth is an enamel loop indicating the
shape of the external valley. The round anterolabial corner suggests only one
labial cusp.
M2 is similar to
M1 (Figure 5.3). Its posterior half is narrower. A weak curvature at the lingual
edge of the tooth indicates presence of a shallow hypostria that marks the
boundary of the protocone and hypocone. A shallow, transverse groove separates
the trigon and the posteroloph of the tooth. M3 is not preserved in any of the
specimens, but the partial alveolus of 141277 indicates a sizeable tooth (Figure
4.1).
Comments:
The Shara Murun fauna is from the Shara Murun
Formation that overlies the “Tukhum” Fm. at the locality Ula Usu. When proposed,
the Shara Murun Fm. was not described and was believed to lie underneath the
Irdin Manha Fm. (Berkey and Granger 1923). Later these two formations were
considered to be roughly equivalent (Berkey and Morris 1924). The Shara Murun
Fm. was formally described by
Berkey and Morris (1927), with a faunal list
presented. Its younger age than Irdin Manha was based on faunal composition, not
on superposition of sediments (Radinsky 1964).
Russell and Zhai (1987, p. 210)
stated that: “nowhere in Inner Mongolia can the Shara Murun Formation be seen
overlying the Irdin Manha.” The superpositional relationship of Shara Murun and
Irdin Manha formations is probably present at the Bayan Ulan section (Meng et
al. 1999). The Sharamurunian is now recognized as the late middle Eocene land
mammal age of Asia (Tong et al. 1995;
Meng and McKenna 1998).
The dentitions
reported here are generally similar to those of the holotype of G. tolmachovi
(AMNH 26142). Compared to AMNH 26142, they have relatively lower crowns, in
which the hypostriid on the lateral surface of the cheek teeth usually ends
above the alveolus for specimens at different wear stages.
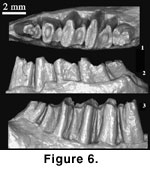 The p3 is critical
to distinguish G. tolmachovi from G. andrewsi (Burke 1941;
Figure
2, Figure 6). The new specimen (AMNH 141288;
Figure 3.1-2) is somewhat intermediate
between those of the holotypes of the two species. The lingual reentrant seems
more posteriorly positioned than that of G. tolmachovi but more
anteriorly than in G. andrewsi. In crown view, the tooth is narrower than
that of G. tolmachovi but wider than that of G. andrewsi. With
further wear, the tooth would become wider. These detailed features suggest that
the holotypes of the two species may actually represent individual variation
within the same species (see below).
The p3 is critical
to distinguish G. tolmachovi from G. andrewsi (Burke 1941;
Figure
2, Figure 6). The new specimen (AMNH 141288;
Figure 3.1-2) is somewhat intermediate
between those of the holotypes of the two species. The lingual reentrant seems
more posteriorly positioned than that of G. tolmachovi but more
anteriorly than in G. andrewsi. In crown view, the tooth is narrower than
that of G. tolmachovi but wider than that of G. andrewsi. With
further wear, the tooth would become wider. These detailed features suggest that
the holotypes of the two species may actually represent individual variation
within the same species (see below).
All species of
Gobiolagus were based on lower teeth except for G. lii (Zhang et al.
2001). Possible upper teeth of Gobiolagus (IVPP V3012) were first
described from the Shara Murun beds at Ula Usu, Inner Mongolia (Li 1965), but
these teeth were considered to be more similar to the North American
Mytonolagus (Qi 1988).
Qi (1988), erroneously citing specimen number V3011,
noted presence of the hypostria on the upper molars of Li’s specimen, which does
not seem comparable to the relatively low crown lower cheek teeth of
Gobiolagus. However, the partial hypostria on V3012 may be lost with further
wear. Moreover, absence of the hypostria on the upper molars does not
necessarily correspond to low crowned lower molars. For instance, Ordolagus
teilhardi has high crown lower teeth but its upper teeth lack the hypostria
(Huang 1986).
Qi (1988) instead
assigned a maxilla with P4-M3 (V8430) from the Shara Murun beds, Ula Usu, to
G. tolmachovi and recognized absence of the hypostria on cheek teeth and the
transverse extension of the upper cheek teeth as features by which V8430 was
assigned to G. tolmachovi. It has been demonstrated that tooth width and
condition of the hypostria in the upper teeth of early lagomorphs are highly
related to age and wear (Huang 1986,
1987). The specimen described by
Qi (1988)
is heavily worn so that its width and absence of the hypostria are at least
partly age related.
P4 of V8430 is
similar to the upper teeth, described above (Figure 4,
Figure 5) but is distinctively
smaller than M1. Qi (1988) considered the P4 of V8430 to be molariform. However,
because the cheek teeth of V8430 were extensively worn so that the tooth pattern
was not well preserved, it is uncertain whether the P4 is molariform. The only
structure that can be recognized from the P4 of V8430 is the anteroloph, which
is complete as in the specimens described in this study. A complete anteroloph
differentiates V8430 from Lushilagus, in which P4 is still trilobate, but
it alone does not necessarily make a molariform P4. The P4 in the specimens
assigned to G. tolmachovi in this study is similar to P3 in general
morphology except for being larger and having a complete anteroloph. It has
neither the division of the trigon and talon, nor division of cusps on the
labial side of the tooth. We consider such a P4 nonmolariform.
V8430 described by
Qi (1988) has the premolar foramen medial to the junction of P4 and M1 on the
palate, which differs from the maxilla described here. The premolar foramen is
usually within the maxilla and medial to the premolars in some lagomorphs
(Matthew and Granger 1923;
Bohlin 1942, figure 17;
Huang 1987). The premolar
foramen is not a stable feature and can vary within a species, such as in
Desmatolagus pusillus (Huang 1987).
Huang (1987, table 18) has surveyed
several species of Desmatolagus, Bellatona and Ochotona and
concluded that the premolar foramen shifts posteriorly in geologically younger
species. For instance, the foramen is medial to P3 in D. gobiensis, but
medial to P4 in Ochotona nihewanica. Compared to those species listed by
Huang, the position of the premolar foramen in V8430 seems to be quite
posteriorly positioned, at the junction of P4 and M1 (Qi 1988, figure 1).
Gobiolagus
andrewsi
Burke, 1941
Emended
Diagnosis: Similar to
Gobiolagus tolmachovi in morphology and size but differing from the latter
in having the p3 more compressed transversely with main lingual reentrant more
posterior (on worn teeth opposite buccal reentrant;
Figure 1.2,
Figure 6).
Locality and
Age: The holotype (AMNH
26091) and a referred specimen (AMNH 26092) were from Jhama Obo, East Mesa,
Shara Murun Region, Inner Mongolia. Another referred specimen (AMNH 26097) was
from Twin Obo, East Mesa of the same region. All of the specimens were from the
Ulan Gochu horizon, which is now considered to be late Eocene (Wang 1997a,
1997b;
Meng and McKenna 1998).
Comments:
The holotype of G.
andrewsi (AMNH 26091; Figure 6) is generally similar to that of G.
tolmachovi, as realized by
Burke (1941), and is from a relatively younger
individual, judging from the degree of tooth wear, than the holotype of G.
tolmachovi. The major difference between the two species is that in G.
andrewsi the p3 is more compressed transversely with the lingual reentrant
opposite the labial reentrant. Other differences listed by
Burke (1941, p. 8)
are relatively trivial, some of which are probably caused by preservation or
artifact. For instance, the lingual side of the anterior half of the p3 in G.
andrewsi was apparently damaged, and the enamel was peeled off, which makes
the tooth appear narrower. As shown in Figure
6, the trigonid and talonid of
cheek teeth in AMNH 26091 are pitted, an unusual condition for any known
lagomorph. The shearing wear of the lagomorph teeth does not seem able to create
such a pitted wear surface on a cheek tooth. The position of the lingual
reentrant varies with wear and probably does so from individual to individual.
As shown above, the new specimen of G. tolmachovi from Ula Usu has a p3
in which the lingual reentrant is more posterior than that in holotype of the
species. It is highly possible that G. andrewsi is but a junior synonym
of G. tolmachovi.
Gobiolagus
major
Burke, 1941
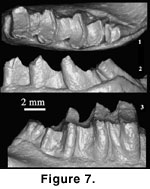 Emended
Diagnosis: Larger than other
species of the genus; p4 talonid less reduced; lower incisor ending at the level
of m2 talonid and forming a distinct protuberance on the lingual surface of the
mandible. Differs from G. tolmachovi and G. lii in having P3-4
triple-rooted; the P4 crescentic valley V-shaped and central cusp triangular; M1
oval in occlusal view (Figure 1.3,
Figure 7).
Emended
Diagnosis: Larger than other
species of the genus; p4 talonid less reduced; lower incisor ending at the level
of m2 talonid and forming a distinct protuberance on the lingual surface of the
mandible. Differs from G. tolmachovi and G. lii in having P3-4
triple-rooted; the P4 crescentic valley V-shaped and central cusp triangular; M1
oval in occlusal view (Figure 1.3,
Figure 7).
Locality and
Age: The holotype of
Gobiolagus major (AMNH 26098) was collected from the
late Eocene Ulan Gochu beds at Urtyn Obo, East Mesa, Shara Murun Region, Inner
Mongolia (Burke 1941). Additional specimens assigned to the species were from Yihesubu, about 18 km southeast of Nomogen village, at the north rim of the Ulan
Shireh platform, Inner Mongolia, of possible Upper Eocene age (Meng and Hu
2004).
Comments:
Burke (1941) named G. major with uncertainty. He noted that the cheek
teeth of G. major were similar to those of Desmatolagus but the p4
was of the Gobiolagus type. After re-examining the holotype and
additional specimens assigned to Gobiolagus major (Meng and Hu
2004), we
consider G. major a valid species that shares several dental features
with other species of the genus, including trigonids of lower cheek teeth
transversely extended, talonids transverse and shorter than trigonids except for
m2, enamel ridge connecting the trigonid and talonid at the lingual side, m2 the
largest cheek tooth, m2 talonid much lower than the trigonid, nonmolariform P4
significantly smaller than M1, P3-4 not lingually extended and M3 reduction
moderate.
Gobiolagus
lii
Zhang, Dawson, and
Huang, 2001
Diagnosis:
“Differs from G. tolmachovi in being smaller, having much less
well-developed buccal cuspules on P4 and M1, and nonmolariform P4” (Zhang et al.
2001, p. 70).
Locality and
Age: Yuli member of the Heti
Formation at Houshipo, Guojia village, Yuanqu County, Shanxi; middle Eocene
(Irdinmanhan).
Comments:
G. lii is the
earliest species of the genus (Zhang et al.
2001). The holotype, the only
specimen, of this species is a fragmentary maxilla with P3-M2 (IVPP V12755).
According to
Zhang et al. (2001, p. 259), taxonomic identification of G. lii
was based primarily on comparison with the maxilla assigned to G. tolmachovi
(Qi 1988). Both species are similar “in the shape of the maxilla, zygoma,
and infraorbital foramen, and in possessing a premolar foramen” (Zhang et al.
2001, p. 259). As we discussed above, the identification of the maxilla as G.
tolmachovi by
Qi (1988) is itself uncertain, which provides a weak basis for
the taxonomic identification of G. lii. However, the specimens we here
assign to G. tolmachovi resemble that of G. lii in some aspects of
dental morphology, e.g., P3-4 morphology. These specimens differ from the
holotype of G. lii in being larger, lacking the premolar foramen, and
having a less convex external outline of the maxilla.
Gobiolagus
burkei sp. nov.
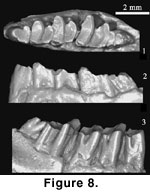 Holotype -
AMNH 141275, a fragmentary
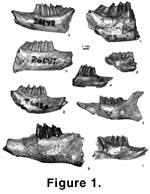
right mandible with p3-m3 (Figure 1.4,
Figure 8).
Holotype -
AMNH 141275, a fragmentary
right mandible with p3-m3 (Figure 1.4,
Figure 8).
Referred
Specimens: AMNH 141276, a
fragmentary mandible with m1-2. AMNH 141275-6 were collected in 1925 under the
same field and catalog numbers (507 and AMNH 92307) as those referred to G.
tolmachovi (see above) from the same locality.
Etymology:
The specific name is in
honor of Dr. J.J. Burke, who made the first systematic study of early Tertiary
lagomorphs from the Mongolian Plateau.
Diagnosis:
Resembles Lushilagus in relatively lower tooth crown and trilobate
lingual shape of p3; similar to Gobiolagus in having the pear-shaped p4
trigonid, short talonid on p4 and m1, lower cheek tooth
trigonid transversely extended, talonid shorter than trigonid except for m2,
enamel ridge connecting the trigonid and talonid at the lingual side, p4
significantly smaller than m1, and m2 the largest cheek tooth; differing
from other species of Gobiolagus in being smaller and having lower crown
cheek teeth, incisor ventral to cheek teeth and lingual protuberance absent, and
incisor extending posteriorly below the talonid of m2; and further differing
from G. tolmachovi and G. andrewsi in having less reduced p4
talonid.
Locality and
Age:
Shara Murun Formation at Ula Usu, Inner Mongolia; late
middle Eocene.
Description:
The smallest species of Gobiolagus (Figure 1.4,
Table 1). The posterior
portion of the incisor is preserved and ends posteriorly below the talonid of
m2. The incisor is ventral to the roots of cheek teeth; therefore, there is no
protuberance on the lingual surface of the mandible. On the lateral surface, two
small posterior mental foramina are under p4 and m1 with the anterior one being
larger than the posterior. The cast of matrix left at the breakage below the
diastema indicates an anterior mental foramen. The mandible measures 4.95 mm
deep and 2.67 thick at m1.
The cheek teeth
are low-crowned, rooted, and the height difference is not distinct for all cheek
teeth (Figure 1.4,
Figure 8). The p3 is longer than wide in occlusal view and its
width increases posteriorly. On the lateral surface a hypostriid extends a third
of the crown height from the occlusal surface and the tooth is divided into a
long, narrow anterior lobe and a short, wide posterior lobe. On the lingual
side, there are two weak grooves (internal reentrants) that are shallower and
shorter than the hypostriid. In occlusal view the tooth is trilobate.
The p4 is
molariform, but is notably smaller than m1. As in molars, the occlusal surface
is uneven, with the lingual side higher than the labial one in both the trigonid
and talonid. The trigonid and talonid are separated by a narrow groove filled
with cement except at the lingual side of the tooth where a narrow enamel band
connects the posterolingual corner of the trigonid with the anterolingual corner
of the talonid. The talonid is short but wide, with the labial side being
narrower than the lingual side. The hypostriid is similar to that of p3.
The m1 is similar
to p4 in general morphology, but is larger than the latter. The m2 is significantly
larger than m1. The trigonid and talonid of m2 are also proportionally larger
than those of m1. In lateral view the hypostriid of m2 is much wider and deeper
than that of m1. Unlike p4 and m1 where the trigonid is slightly longer than the
talonid, the talonid of m2 is longer than the trigonid, although the difference
is minor. The enamel on the lingual surface of m3 was broken so that the tooth
appears smaller than it is. The m3 is greatly reduced but still has both the
trigonid and talonid lobes. The trigonid of m3 is wider, but shorter, than the
talonid. The hypostriid of m3 is the narrowest and shortest of the cheek teeth.
Comments:
AMNH 141275 is considered a
species of Gobiolagus because it has the basic features of the genus,
including p3 and m3 proportionally smaller than those of Shamolagus but
larger than those of Desmatolagus, incisor extending to m2, lower cheek
tooth trigonid transversely extended, talonid transverse and shorter than
trigonid except for m2, enamel ridge connecting the trigonid and talonid at the
lingual side, p4 significantly smaller than m1, m2 the largest cheek tooth, and
talonid of m2 much lower than the trigonid in contrast to the condition in p4 or
m1.
Gobiolagus
burkei differs from other
species of the genus in being smaller and having relatively low-crowned cheek
teeth. Gobiolagus lii is probably similar to the new species in size.
However, because G. lii is represented only by upper teeth and G.
burkei by lower ones, the comparison between the two species is difficult.
In species of early lagomorphs where upper and lower dentitions from the same
individual are known, such as Shamolagus medius (Li 1965), a lower cheek
tooth is usually shorter than its upper counterpart. Given that relationship, it
may be estimated that G. burkei is slightly smaller than G. lii
(Zhang et al. 2001, table 1). Taking the different age and geographic
distributions into account, we recognize both G. lii and G. burkei.
Gobiolagus
burkei further differs from
G. tolmachovi and G. andrewsi, but is similar to G. major,
in having a less reduced p4 talonid. Its incisor extends to the level of the
talonid of m2 and is less lingually offset than in other species. The p3 of
G. burkei is similar to that of Shamolagus. One of the features
originally diagnosing Gobiolagus was p3 “with modified Shamolagus
pattern” (Burke 1941, p. 5). However, the p3 is unknown in G. major. In
G. tolmachovi the anterior lobe of p3 is large and has a rounded occlusal
outline. In G. andrewsi p3 is considered more compressed transversely
(Burke 1941).
Genus Lushilagus
Li, 1965
Type Species:
Lushilagus lohoensis Li, 1965.
Emended
Diagnosis: Small lagomorphs
with relatively low-crowned cheek teeth; P3 with large lingual lobe; P4
trilobate with an incomplete anteroloph; upper cheek teeth not transversely
extended; labial tooth enamel folds absent when worn; M3 not significantly
reduced; p3 anterior lobe conical and posterior lobe triangular, lingual side
with one or two reentrants; m3 with the third lobe.
Included
Species: Lushilagus
danjiangensis
Tong and Lei, 1987.
Locality and
Age: Lushih (=Lushi)
Formation, Menchiapu (=Mengjiapu), Lushih District, Honan (=Henan) (Li 1965); Hetaoyuan Formation, Shipigou, Xichun County, Henan (Tong and Lei 1987;
Tong
1997); Liyang, Jiangsu, China (Qi et al. 1991); middle Eocene.
Comments:
Lushilagus lohoensis was based on upper dentition (Li 1965;
Qi et al. 1991) and some isolated lower teeth were later assigned to the species (Tong
1997). All specimens of L. danjiangensis are isolated teeth. This genus
is primarily diagnosed by primitive characters and represents one of the
earliest lagomorphs known.
Erbajeva (1999)
transferred L. danjiangensis to Zaissanolagus. The type species of
Zaissanolagus, Z. gromovi, was based on a P3 discovered from the
late Oligocene Buran Formation in the Zaisan Depression (Erbajeva 1999). One of
the features that distinguish Zaissanolagus from other Paleogene genera
of the family Leporidae is the proportion of P3 dimensions. According to
Erbajeva (1999), the P3 ratio of length to width is 49.3% in Zaissanolagus,
98% in Shamolagus, 83% in Lushilagus, and 43.5% in Gobiolagus.
However, the length and width of P3 are 1.2 and 1.8 mm in Lushilagus
lohoensis, and 1.3 and 2.1 mm in Shamolagus (Li 1965, p. 28), which
give ratios of 67% and 62%, respectively. The P3 ratio is 46% (1.9 by 4.1 mm) in
Gobiolagus based on the measurements of
Qi (1988, table 1) and 45% (1.0
by 2.2 mm) in L. danjiangensis (Tong and Lei 1987, table 2). Moreover,
the P3 ratio of Zaissanolagus gromovi is 81% based on the measurements
given by Erbajeva (1999, 1.5 by 1.85 mm in table 1). These numbers are
drastically inconsistent. Because the sources of data and methods of calculation
used in Erbajeva (1999) are unclear to us, we are unable to verify the
significant differences among these numbers. Nonetheless, it is clear that the
P3 ratio is distinctive between Z. gromovi and L. danjiangensis;
thus we recognize L. danjiangensis until additional evidence proves
contrary.
Genus
Strenulagus
Tong and Lei,1987
Type Species:
Strenulagus shipigouensis
Tong and Lei,1987, the only known species of
the genus.
Diagnosis:
“Unilateral hypsodonty distinct. Anterior lobe of p3 conical, and narrower than
posterior lobe; trigonid of p4-m2 compressed anteroposteriorly; talonid of p4
smaller relative to its trigonid, with distinctly compressed labial side and
even lingual wall; on m1-2 talonid large, with a third lobe on little worn
teeth; m3 less reduced, with a clear hypoconulid; p3 and p4 double-rooted, with
trilobate occlusal surface; P3 labial lobe relatively developed; P4 metastyle
enlarged, usually connected with parastyle; preprotocrista of P4 frequently
unconnected with parastyle; metacone of M1 isolated, surrounded by U-shaped
enamel valley; M2 and M3 consisting of V-shaped trigon and obliquely elongated
hypocone; M3 less reduced” (Tong 1997, p. 202).
Locality and
Age: Hetaoyuan Formation,
Shipigou, Xichun County, Henan, China; Irdinmanhan, middle Eocene.
Comments:
Strenulagus shipigouensis was named by
Tong and Lei (1987) and additional
specimens led to a thorough revision of the species diagnosis (Tong 1997).
Genus
DITUBEROLAGUS Tong, 1997
Type Species:
Dituberolagus venustus
Tong, 1997, the only known species of the genus.
Diagnosis:
“A small primitive lagomorph. Trigonid of midcheek teeth made up of conical
protoconid and metaconid; valley between the two cusps deep, extending down
along anterior wall of tooth; talonid of m2? longer and wider than trigonid;
posterior lobe of p3 cingulum-like” (Tong 1997, p. 204).
Locality and
Age: Hetaoyuan Formation at
Shipigou, Xichun County of Henan, China; Irdinmanhan, middle Eocene.
Comments:
Averianov (1998) noted that
the lower teeth of Dituberolagus venustus are similar to those of
Bulatia aksyirica (Gabunia and Shevyreva 1994).
Bulatia aksyirica is based on an isolated m1 or m2 from the middle Eocene
Sargamyss Formation in Zaisan depression and was originally referred to
Mixodontia incertae familiae.
Averianov (1998) considered Bulatia aksyirica
a nomen dubium and referred it to as Mammalia incertae ordinis.
Averianov (1998,
p. 207) suggested that the M2 referred to Lushilagus? danjiangensis by
Tong (1997) may belong to Dituberolagus venustus and that the tooth is
“noticeably similar to upper molars of mimotonid Aktashmys montealbus
from the early Eocene of Kirghizia.”
Genus
Shamolagus
Burke, 1941
Type Species:
Shamolagus grangeri
Burke, 1941.
Emended
Diagnosis: Teeth relatively
low crowned; p3 trilobate; talonid of cheek teeth large; m3 not reduced and with
a distinct hypoconulid; P4 not significantly smaller than M1; P4 crescentic
valley with posterolabial extension; worn upper molar with circular central
enamel lake.
Included
Species:
Shamolagus medius
Burke, 1941 (see below for S. ninae
Gabunia, 1984).
Locality and
Age: Middle Eocene Ulan
Shireh beds at the Chimney Butte locality, North Mesa, Shara Murun Region, Inner
Mongolia; late middle Eocene Shara Murun Formation near Baron Sog, Inner
Mongolia; middle Eocene of Heti Formation, Yuanqu, Shanxi Province.
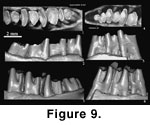 Comments:
Shamolagus
represents one of the earliest
lagomorph genus in Asia. The type species, Shamolagus grangeri, is based
on a mandible with p4-m3 (holotype, AMNH 26289;
Figure 1.6,
Figure 9.1-3) that came
from “Ulan Shireh, Upper Eocene. Irdin Manha Beds” at the locality “Chimney
Butte, North Mesa, Shara Murun Region, Inner Mongolia” (Burke, 1941, p. 1).
The Ulan Shireh beds at North Mesa of Shara Murun area
were originally equated to the “Tukhum”
Fm. (Berkey et al. 1929;
Szalay and Gould 1966), but were also considered to be
above the “Tukhum”
Fm., underlying an unnamed sandy formation (Russell and Zhai 1987). In his
faunal analysis, Ye (1983) used the term “Ulan Shireh beds” and thought that the
Ulan Shireh fauna was a taphocoenosis of the Irdin Manha fauna. He did not
explain why “beds” rather than “formation” was employed, but from the context it
seems implied that the “Ulan Shireh beds” is part of the Irdin Manha Fm.
Comments:
Shamolagus
represents one of the earliest
lagomorph genus in Asia. The type species, Shamolagus grangeri, is based
on a mandible with p4-m3 (holotype, AMNH 26289;
Figure 1.6,
Figure 9.1-3) that came
from “Ulan Shireh, Upper Eocene. Irdin Manha Beds” at the locality “Chimney
Butte, North Mesa, Shara Murun Region, Inner Mongolia” (Burke, 1941, p. 1).
The Ulan Shireh beds at North Mesa of Shara Murun area
were originally equated to the “Tukhum”
Fm. (Berkey et al. 1929;
Szalay and Gould 1966), but were also considered to be
above the “Tukhum”
Fm., underlying an unnamed sandy formation (Russell and Zhai 1987). In his
faunal analysis, Ye (1983) used the term “Ulan Shireh beds” and thought that the
Ulan Shireh fauna was a taphocoenosis of the Irdin Manha fauna. He did not
explain why “beds” rather than “formation” was employed, but from the context it
seems implied that the “Ulan Shireh beds” is part of the Irdin Manha Fm.
The holotype of S. medius (AMNH 26144;
Figure 1.5,
Figure 9.4-6) is a fragmentary right mandible with p3-m1from Shara Murun Formation near Baron Sog, Inner Mongolia, and additional material
was reported from the Ula Usu (Li 1965).
Shamolagus
grangeri and S. medius
are similar in size and general tooth structures (Figure 9). Both are primitive
in having relatively low-crowned teeth. An
important character of S. medius is its trilobate p3 (Burke, 1941; Li
1965;
Figure 9.4). Other diagnostic features seem to be minor. Because p3
is not preserved in the holotype of S. grangeri, the difference between
the two species is not fully revealed. The specimen from Ula Usu (Li 1965) and
those from the Heti Formation (Tong 1997) assigned to S. medius and S.
cf. medius, respectively, have a smaller m3 than that of S. grangeri.
A reduced m3 with an indistinct hypoconulid lobe is by far the most convincing
feature distinguishing S. medius from S. grangeri.
Isolated teeth
assigned to Shamolagus sp. and Shamolagus cf. medius were
also reported from the middle Eocene of Henan and Shanxi (Tong 1997).
Genus
Hypsimylus
Zhai, 1977
Type Species:
Hypsimylus beijingensis
Zhai, 1977.
Emended
Diagnosis: Large lagomorphs
with hypsodont cheek teeth; the third lobe of p4-m3 distinct in relatively young
individuals.
Included
Species: Hypsimylus
yihesubuensis
Meng and Hu,
2004. Hypsimylus yihesubuensis differs from H. beijingensis in
being larger and having relatively lower crown height and wider m1 (Meng and Hu
2004).
Locality and
Age: Late middle Eocene
Changxindian Formation, Beijing; Yihesubu locality, Inner Mongolia; late Eocene.
Comments:
The type species of the genus, Hypsimylus beijingensis, is based on lower
teeth and was originally assigned to Eurymylidae because of the high-crowned
cheek teeth that are somewhat similar to those of Rhombomylus (Zhai 1977). Although recognizing dental similarities between Hypsimylus and
lagomorphs, Zhai (1977) noted that the cheek teeth of Hypsimylus are
larger and more hyposodont than those of Eocene lagomorphs, and uniquely have a
third lobe on lower cheek teeth.
Li and Ting (1985) placed Hypsimylus in
Mimotonidae.
Dashzeveg and Russell (1988) found no significant character to
indicate a relationship of Hypsimylus to mimotonids and established the
subfamily Hypsimylinae under Eurymylidae.
McKenna and Bell (1997) regarded
Hypsimylus as a paleolagine lagomorph and placed it within Leporidae.
Averianov (1998) considered that Hypsimylus is similar to Valerilagus
reshetovi Shevyreva (1995; see below) and considered Hypsimylus as
Lagomorpha incertae familiae.
Meng and Hu (2004) described Hypsimylus
yihesubuensis from the late Eocene Yihesubu locality and confirmed that
Hypsimylus is not a eurymylid, but a lagomorph. The trilobate tooth pattern
seen in Hypsimylus is not uncommon in lagomorphs; it is usually present
on immature specimens, as noted in the North American Megalagus turgidus
(Wood 1940) and Palaeolagus philoi (Dawson 1958).
Genus
Desmatolagus
Matthew and Granger, 1923
Type Species:
Desmatolagus gobiensis
Matthew and Granger, 1923.
Included
Species: There are two
Eocene species of the genus: Desmatolagus vetustus and D. ardynense.
Locality and
Age: Eocene species are from
Ulan Gochu beds, Jhama Obo, East Mesa, Shara Murun Region, Inner Mongolia; Ardyn
Obo (Ergilin-Dzo), Mongolia; late Eocene.
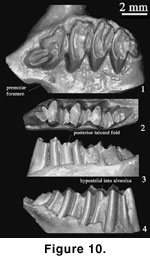 Comments:
Because the type species of the genus, D. gobiensis, is from the early
Oligocene Hsanda Gol Formation, which is beyond the scope of this study, we do
not provide a diagnosis for the genus except for the photographs of the type
specimens of D. gobiensis (Figure 1.9,
Figure 10). About 10 species of
Desmatolagus have been named from Asia and North America in various studies
(Matthew and Granger 1923;
Teilhard de Chardin
1926; Burke 1936,
1941;
Bohlin 1937).
Huang (1987) considered the taxonomic
position of the North American Desmatolagus uncertain and concluded that
there were five valid species of Desmatolagus in Asia, including D.
ardynense, D. vetustus, D. gobiensis, D. robustus, and D. pusillus.
The genus is mainly characterized by reduction of P2/p3 and M3/m3 (Huang 1987).
Among these species, D. vetustus and D. ardynense (Burke 1941) are
from late Eocene Ulan Gochu beds, Inner Mongolia and Ardyn Obo beds, Mongolia.
Comments:
Because the type species of the genus, D. gobiensis, is from the early
Oligocene Hsanda Gol Formation, which is beyond the scope of this study, we do
not provide a diagnosis for the genus except for the photographs of the type
specimens of D. gobiensis (Figure 1.9,
Figure 10). About 10 species of
Desmatolagus have been named from Asia and North America in various studies
(Matthew and Granger 1923;
Teilhard de Chardin
1926; Burke 1936,
1941;
Bohlin 1937).
Huang (1987) considered the taxonomic
position of the North American Desmatolagus uncertain and concluded that
there were five valid species of Desmatolagus in Asia, including D.
ardynense, D. vetustus, D. gobiensis, D. robustus, and D. pusillus.
The genus is mainly characterized by reduction of P2/p3 and M3/m3 (Huang 1987).
Among these species, D. vetustus and D. ardynense (Burke 1941) are
from late Eocene Ulan Gochu beds, Inner Mongolia and Ardyn Obo beds, Mongolia.
The holotype of
D. vetustus (AMNH 26089; Figure 1.7,
Figure 10.1-3) from the Ulan Gochu beds,
Jhama Obo, Inner Mongolia, is the earliest and most primitive species of
Desmatolagus (Huang 1987). Because of its primitive low-crowned cheek teeth,
this species has been removed from Desmatolagus and placed in
Procaprolagus (Gureev 1960;
Sych 1975;
de Muizon 1977). According to
Gureev
(1960) and de Muizon
(1977), Procaprolagus is characterized by
low-crowned teeth in which the lingual and labial grooves do not enter the
alveolus, lack of the premolar foramen on the palate medial to the premolar,
lack of the third loph on p4 and lower molars, transverse extension of the
trigonids of lower p4 and molars, and presence of an enamel spike projecting
posteriorly from the posterior wall of the trigonid.
De Muizon (1977) synonymized P. vetustus with Desmatolagus vetustus and P.
radicidens with D. radicidens, but
Sych (1975) regarded these two
species of Dasmatolagus as junior synonyms of Desmatolagus
gobiensis. Huang (1987), following
Sych (1975), synonymized D. radicidens
with D. gobiensis but retained D. vetustus as a valid and
primitive species of Desmatolagus; he considered the basic tooth pattern
and reduction of terminal teeth in D. vetustus as typical of
Desmatolagus. He also recognized that the premolar foramen is variably
present within individuals of the same species, such as D. pusillus.
Erbajeva and Sen (1998) again synonymized Procaprolagus with
Desmatolagus and pointed out that the third lophid on the lower cheek teeth
of some Desmatolagus is a juvenile feature, which is therefore not
adequate to distinguish Procaprolagus as a different genus from
Desmatolagus.
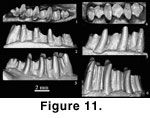 In addition, we note that the enamel spike on the posterior
wall of the trigonid is also present in the holotype of D. vetustus
(Figure 11.1).
In addition, we note that the enamel spike on the posterior
wall of the trigonid is also present in the holotype of D. vetustus
(Figure 11.1).
In a recent study
on lagomorphs from the late Eocene of Inner Mongolia,
Meng and Hu (2004) noted
that differences of D. vetustus from other species of Desmatolagus
are significant. In D. vetustus the lower teeth are relatively low
crowned, with the hypostriid and the lingual groove shallow and above the
alveolus. The height of cheek teeth decreases posteriorly to a lesser degree.
There is no posterior talonid fold in lower cheek teeth of known specimens. The
talonid is relatively narrow compared to the trigonid, or the trigonid is more
transversely extended. The premolar foramen is absent. In other species,
including D. gobiensis, D. robustus, D. pusillus, and D. ardynense,
the cheek teeth have much higher crowns and the hypostriid extends into the
alveolus. The anterior cheek teeth are much higher than m2 and m3. The posterior
talonid fold is usually present in relatively young individuals. The talonid is
relatively wider. Although the differences are truly present, most of those
listed for D. vetustus are primitive, except the relative narrower
talonid. In addition, the posterior border of the palate is shifted more
anteriorly than that in D. gobioensis. We follow
Huang (1987) to
recognize D. vetustus.
The holotype of
D. ardynense (AMNH 20373; Figure 1.8,
Figure 10.4-6) is
from Ardyn Obo, Mongolia. It was originally identified as D. robustus
(Matthew and Granger 1925) and was later named as D. ardynense by
Burke (1941).
Matthew and Granger (1925) noted the “minute vestigial stump of a tooth”
anterior to the alveolus of the p3, which was correctly indentified as the root
of the dp3 (Burke 1941). The species is generally similar to D. robustus
in morphology and size, but differs from the latter in having smaller p4, cheek
teeth less compressed anteroposteriorly, and m1 the largest lower cheek tooth.
Because the p3 and m3 are not preserved in the holotype, whether these teeth are
reduced in size is unknown. In addition, because the specimen is from a
relatively young individual, judging from retention of the dp3 root and presence
of the posterior internal reentrant on m2, the features that distinguish the
species from D. robustus, such as the relatively smaller p4 and longer
talonid, may be age-related. Therefore, it is still possible that AMNH 20373
belongs to D. robustus. Nonetheless, this specimen represents a hypsodont
species of lagomorph from the Eocene. Other species with similar hypsodonty are
assigned to the genus Hypsimylus.
The age of Ardyn
Obo beds is worth of a note. The “Ardyn Obo Formation”
was proposed by
Berkey and Granger (1923) and accepted thereafter (Berkey and
Morris 1924,
1927). The locality, marked by the Ardyn Obo, was found at mile 487
along the Kalgan-Uliasutai Trail (Berkey and Morris 1927, fig 84).
Berkey and
Morris (1927, p. 176) wrote: “The word ‘ardyn,’ meaning ‘jewels,’ proved to be
the same word which we had spelled ‘irdin’ farther east. It refers to the
glittering, highly polished pebbles of quartz and chalcedony which are found in
the upper sandstones of the formation.” The escarpment was afterwards explored
more extensively by several expeditions during a 30 year period from 1946 to
1974 (Russell and Zhai 1987). The entire escarpment was then called the Ergilin
Dzo, whereas the Ardyn Obo was equated to Ergil Obo.
Russell and Zhai (1987)
used “Ergilin-Dzo Svita” to include all the beds from the escarpment and cited
Desmatolagus robustus, actually D. ardynense, as a member of the
Ergilin-Dzo fauna. The Ergilin-Dzo fauna is now the basis for Ergilian, the late
Eocene Asiatic land mammal age that has been widely recognized (Emry et al.
1998; Meng and McKenna 1998).
LAGOMORPHA
incertae sedis
Several
fragmentary specimens collected from the Eocene of
Kazakhstan and Kyrgyzstan
were assigned to various lagomorph species. The taxonomy of these species
remains controversial, and the published data are insufficient to review them
meaningfully. We only provide brief comments on their documentation.
Shamolagus ninae
Gabunia (1984) is based on one m1 and one m2 from the late early or early middle
Eocene Obaila Formation, at the Aksyir locality, Zaisan Basin, Kazakhstan. The
teeth are small (m1 length / width, 1.8 x 2mm; m2, 1.8 x 1.9), brachydont, and
are characterized by high and narrow trigonids, which are distinctive from other
species of Shamolagus.
Shevyreva (1996) regarded S. ninae a eurymylid, but
Averianov (1998)
considered it a lagomorph and treated the species as Lagomorpha incertae
familiae because the poor specimens lack any diagnostic feature at the generic
level.
Romanolagus hekkeri
and Valerylagus reshetovi, purported lagomorphs, were mentioned by
Shevyreva (1994) but were described in
Shevyreva (1995), in which the latter
species was spelled as Valerilagus reshetovi. The specimens of the two
taxa are from the middle Eocene or possibly latest early Eocene Andarak 2
locality, Kyrgyzstan (Shevyreva 1995;
Averianov
1998; Averianov
and Godinot 1998).
Romanolagus hekkeri is based on a maxillary fragment with M1.
Averianov (1998, p. 207) noted that “the holotype of
R. hekkeri could not be distinguished by the size or morphology
from the known upper molars of the mimotonid Anatolimys rozhdestvenskii
described from the same locality.” Thus, Averianov (ibid.) regarded R.
hekkeri a junior subjective synonym of A. rozhdestvenskii, a
mimotonid. Valerilagus reshetovi is based on a fragmentary maxilla with
labial portions of P3-4. Additional specimens are available from the Andarak 2
locality (Averianov 1996) but have not been described to our knowledge.
Valerilagus reshetovi was placed in Mytonolagidae (Averianov 1996; Averianov and Godinot 1998), but the family is treated
as a synonym of Leporidae (McKenna and Bell 1997). The line drawing of the holotype of V. reshetovi (Shevyreva 1995,
figure 1) convincingly
shows lagomorph morphology, as endorsed by
Averianov
and Godinot (1998).
Annalagus
margarita is based on
several isolated teeth from the late Eocene Aksyir Formation, Baldys, Zaisan
depression, Kazakhstan (Shevyreva 1996). The holotype of the species is a dp3.
The species was assigned to Amphilaginae,
Palaeolagidae, Lagomorpha.
Averianov (1998, p. 207)
argued that dp3 is insufficient to diagnose the species and thus regarded
Annalagus margarita Shevyreva, 1996 as a nomen dubium and suggested
that the species “could be provisionally referred to the family ‘Mytonolagidae’
(paraphyletic group).”
