SYSTEMATIC PALEONTOLOGY
(continued)
Order
CARNIVORA Bowdich, 1821
Infraorder ARCTOIDEA Flower, 1869
Family
AMPHICYONIDAE Haeckel, 1866
Genus
ICTIOCYON Crusafont-Pairó
et al., 1955
Ictiocyon cf. I. socialis
(Schlosser 1904)
?Ictiocyon sp.:
Qiu et al. 1997, p. 186.
Ictiocyon cf. I. socialis
(Schlosser 1904):
Qiu et al. 2001, p. 378.
Materials: IVPP V12185, partial right horizontal
ramus with anterior half of p4 and alveoli of c1-m3, collected from IVPP
locality 9308, N 36° 13' 52.1" E 103° 36' 23.9", south of the village of
Qujiachuan, on a small road cut in the first white sands, Middle Member of the
Xianshuihe Formation, Zhangjiaping l.f., early Miocene (see further discussion
in Associated Fauna and Age under Hyaenodon weilini).
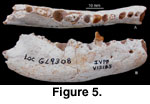 Description: This large fox-sized carnivoran has a
robust horizontal ramus (Figure 5), which is rather deep relative to its length.
The jaw depth is 22.8 mm below the talonid of m1 (on the lingual side). The
anteroventral corner of the masseteric fossa is preserved and located rather
high, about 10 mm above the ventral border of the ramus. There are two mental
foramina: one below the anterior root of the p2 and the other below the
posterior root of the p3. The premolar toothrow is short, and the p3-m1 are
apparently in close contact or even imbricate slightly. There is a short
diastema between the p2 and p3. The p1 and m3 are single rooted, whereas the
p2-m2 are all double rooted. The anterior half of the p4 indicates a short but
rather high-crowned tooth, its anterior ridge of the main cusp being rather
erect as compared to the more recumbent anterior ridges in many amphicyonids.
Its anterior cingulum swells slightly to form a low, cingulum cusp. Alveoli of
the corresponding teeth indicate a robust lower canine, a broad m1-2, an
elongated m2, and a reduced m3.
Description: This large fox-sized carnivoran has a
robust horizontal ramus (Figure 5), which is rather deep relative to its length.
The jaw depth is 22.8 mm below the talonid of m1 (on the lingual side). The
anteroventral corner of the masseteric fossa is preserved and located rather
high, about 10 mm above the ventral border of the ramus. There are two mental
foramina: one below the anterior root of the p2 and the other below the
posterior root of the p3. The premolar toothrow is short, and the p3-m1 are
apparently in close contact or even imbricate slightly. There is a short
diastema between the p2 and p3. The p1 and m3 are single rooted, whereas the
p2-m2 are all double rooted. The anterior half of the p4 indicates a short but
rather high-crowned tooth, its anterior ridge of the main cusp being rather
erect as compared to the more recumbent anterior ridges in many amphicyonids.
Its anterior cingulum swells slightly to form a low, cingulum cusp. Alveoli of
the corresponding teeth indicate a robust lower canine, a broad m1-2, an
elongated m2, and a reduced m3.
Measurements of IVPP V12185 (all in alveolar
length and in mm): p1 = 2.9, p2 = 7.9, p3 = 7.3, p4 = 9.8, m1 = 16.8, m2 = 8.7,
m3 = 3.8, and p1-m3 = 62.3.
Comparison: From what little is preserved, IVPP
V12185 possesses amphicyonid characters such as a high position (relative to the
ventral border) of the masseteric fossa and a large m2. The Lanzhou specimen is
slightly more derived than European and North American Oligocene small- to
medium-sized amphicyonids (such as Pseudocyonopsis, Cynelos,
Daphoenus, and Temnocyon) in its shortened premolar toothrow and
correspondingly broad and closely spaced premolars.
Ginsburg (1992) recognized two distinct lineages
of small, short-snouted amphicyonids in the early to middle Miocene of Europe:
an early Miocene Ictiocyon and a mostly middle Miocene Pseudarctos,
in contrast to an earlier hypothesis of a single anagenetic lineage of
Pseudarctos that decreases in size through time (Kuss 1965). The Lanzhou
specimen readily falls in the size range of both Pseudarctos and
Ictiocyon, and has similarly short and closely spaced premolars, derived
characters shared by both genera. In addition, V12185 has a rather long m2
relative to its m1, a derived feature that is well-developed in I. socialis
(Ginsburg 1992, figure 4, plate II), and still primitively retains a p1, which
was lost in Pseudarctos. Therefore, the Lanzhou specimen is here referred
to Ictiocyon based on its possession of a large m2 and its lack of a
Pseudarctos synapomorphy of the loss of p1. Caution, however, must be
exercised in an identification based on so little material.
A short rostrum and imbricated premolars also
characterize the European Oligocene Brachycyon. However, Brachycyon
is much larger and has a much more robust lower jaw than the Lanzhou specimen.
In addition, the horizontal ramus of Brachycyon tends to expand ventrally
near both its anterior and posterior ends (Ginsburg 1966) in contrast to a
rather straight ventral border in the Lanzhou specimen. Finally, the m2 of
Brachycyon is not enlarged as in the Lanzhou specimen.
In Europe, the monotypic Ictiocyon socialis
is known from several early Miocene localities (Ginsburg 1992;
Viranta 1996;
Ginsburg 1999a), Wintershof (MN3), Artenay (MN4a), Solnhofen fissures (MN4), and
El Canyet (MN4). Pseudarctos bavaricus, on the other hand, occurs later
in MN4b through MN9, and a series of three subspecies of P. bavaricus
were recognized by Ginsburg to represent a morphocline of increasing size
through time. The present recognition of a Chinese occurrence of Ictiocyon
extends its geographic range to East Asia and suggests an early Miocene age of
the first white sands in the Lanzhou strata.
Amphicyonidae gen. et sp. indet.
Material: IVPP V13574, posterior half of left p4,
acquired from local villagers, specimen probably from near IVPP loc. 8801 in
Duitinggou area, in basal white sandstones, Middle Member of the Xianshuihe
Formation, Zhangjiaping l.f., early Miocene (see further discussion in
Associated Fauna and Age under Hyaenodon weilini).
Comments: Although of large size, this tooth is
primitive in its retention of a well- developed posterior cingulum and posterior
accessory cusp, features that tend to occur in less derived species of
amphicyonids (most derived amphicyonids tend to reduce and simplify their
premolars through time). On the other hand, its broadened heel (11.0 mm wide)
reminds us of robust forms such as Gobicyon (Colbert 1939), which is
known only in the middle Miocene of China (a jaw from Serbia referred to
Gobicyon by
Pavlovic and Thenius [1959] was recently transferred to
Haplocyonoides by Ginsburg [1999a]). With the scanty material available, it
is not possible to further explore the identity of this form.
Musteloidea indet
?Plesictis sp.:
Qiu et al. 1997, p. 186.
Plesictis cf. P. vireti
Dehm,
1950: Qiu et al. 2001, p. 378.
Material: IVPP V12190, isolated right m1, from
IVPP locality 9309, N 36° 13' 44.1" E 103° 36' 18.6", Daochazigou, near a sand
and gravel quarry south of Qujiachuan, in the third white sands in the Middle
Member of the Xianshuihe Formation, Zhangjiaping l.f., early Miocene (see
further discussion in Associated Fauna and Age under Hyaenodon weilini).
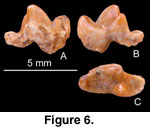 Comments: This isolated carnassial (Figure 6), 4.6
mm long and 2.1 mm wide, is the smallest carnivoran from the Lanzhou Basin,
smaller than any known early Miocene carnivoran in Asia. The metaconid is well
developed, slightly offset (located slightly posterior to) from the protoconid
in occlusal view, and of approximately the same height as the paraconid. The
talonid appears to be narrow, but this is partly due to damage to the labial
portion of the talonid. The talonid is dominated by a centrally located
hypoconid (only the lingual half is preserved), and the entoconid is poorly
differentiated. The possibility that this tooth is a deciduous p4 cannot be
ruled out.
Comments: This isolated carnassial (Figure 6), 4.6
mm long and 2.1 mm wide, is the smallest carnivoran from the Lanzhou Basin,
smaller than any known early Miocene carnivoran in Asia. The metaconid is well
developed, slightly offset (located slightly posterior to) from the protoconid
in occlusal view, and of approximately the same height as the paraconid. The
talonid appears to be narrow, but this is partly due to damage to the labial
portion of the talonid. The talonid is dominated by a centrally located
hypoconid (only the lingual half is preserved), and the entoconid is poorly
differentiated. The possibility that this tooth is a deciduous p4 cannot be
ruled out.
With the poor preservation, IVPP V12190 cannot be
easily identified. The relatively low-crowned trigonid and well-developed
metaconid suggest a caniform carnivoran (in contrast to higher crowned
feliforms). Among early Miocene caniforms, certain European basal musteloids,
such as Plesictis (Wolsan 1993;
Ginsburg 1999a), are closest in size and
overall morphology, although a definite match awaits discovery of additional
specimens.
