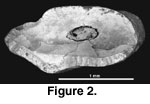
 Referred material. BUMP 1021, trigonid of left lower
cheek tooth (either a p4, m1, or m2) from Bukwa IIB.
Referred material. BUMP 1021, trigonid of left lower
cheek tooth (either a p4, m1, or m2) from Bukwa IIB.Order LAGOMORPHA
Family Ochotonidae Thomas, 1897
Genus and species
undetermined
Figure 2
Referred material. BUMP 1021, trigonid of left lower
cheek tooth (either a p4, m1, or m2) from Bukwa IIB.
Description. The only specimen is fragmentary. It is wider than long: width is 1.80 mm and length is 0.92 mm. Most of the anterior border is convex anteriorly and there is a distinct antero-external fold, which is not filled with cement. The posterior border has a sharp posterior inflection (postprotoconid crista of Mein and Pickford 2003, or “rib”) about one-third of the way from the labial side. Enamel is relatively thin along the anterior border of the tooth but is thickened labially. The posterior border has thick enamel, which projects as a ridge. Enamel along the ridge is smooth and unbroken, indicating that it was not continuous with the talonid (i.e., the trigonid and talonid were separated by cementum).
Remarks. The earliest African lagomorphs are early Miocene records of the ochotonids (pikas) Kenyalagomys (Kenya: MacInnes 1953; Namibia: Hendey 1978; Morocco: Janvier and De Muizon 1976; Israel: Tchernov et al. 1987; Libya: Wessels et al. 2003, cf. Kenyalagomys) and Austrolagomys (Namibia; Stromer 1926; Hopwood 1929; Hendey 1978; Mein and Pickford 2003). Mein and Pickford (2003) considered Kenyalagomys to be a junior synonym of Austrolagomys. The leporids (rabbits and hares) are unknown in Africa until the late Miocene (Winkler 2003). Fossil lagomorphs are extremely rare from Uganda. Earlier collecting at Bukwa by A. Walker and colleagues produced at least one specimen (Walker, personal commun., 2002), but this specimen(s) has not yet been relocated.
BUMP 1021 is extremely fragmentary and cannot be assigned confidently to species. It is assigned to Family Ochotonidae based on presence of a distinct antero-external fold. Comparable development of this fold is more commonly, although not exclusively, seen on ochotonids than on leporids (e.g., may be distinct in the leporids Serengetilagus and Pronolagus). Development of the antero-external fold on BUMP 1021 is within the range of variation seen on “Kenyalagomys” minor and “K.” rusingae. The fold is a more gentle curve on the type of Austrolagomys hendeyi (AD 418’99; Mein and Pickford 2003, figure 1) and on the holotype of A. simpsoni (AMNH 22528; this species considered synonymous with A. inexpectatus by Mein and Pickford 2003).
Order RODENTIA
Suborder Hystricognathi
Superfamily Thryonomyoidea
The small rodents from Bukwa are most similar in morphology and size with members of the Family Phiomyidae. Membership within the Phiomyidae, and the higher level systematics of this group are controversial (Holroyd 1994) and beyond the scope of the present paper. We follow Holroyd’s (1994) proposed phylogeny for the Fayum thryonomyoids, which includes Phiomys, Metaphiomys, and a new genus as members of the Phiomyidae. Holroyd (1994) did not include a detailed study of the East African Miocene phiomyids. The Bukwa rodents described here are compared with published descriptions and illustrations (primarily) and casts of phiomyids from Uganda (Bukwa; Lavocat 1973), Kenya (Lavocat 1973), Egypt (Wood 1968; Holroyd 1994), Libya (Fejfar 1987), Oman (Thomas et al. 1989; 1999) and Algeria (Jaeger et al. 1985). Stevens et al. (2004; in press) have reported phiomyids from Tanzania, but only a lower molar has been described (no comparable tooth position from Bukwa is reported here).
The best sample of phiomyids, both numerically and taxonomically, is from the late Eocene to early Oligocene Jebel Qatrani Formation, Fayum, Egypt. The seminal work on the Fayum rodents was by Wood (1968). Extensive new collecting in the Fayum lead to a major revision of the Fayum rodents by Holroyd in her dissertation (1994). This revision has not yet been formally published. Holroyd (1994) conservatively left the Fayum rodents in the Superfamily Thryonomyoidea, and proposed grouping them into two families, the Phiomyidae (which she revised) and the Thryonomyidae.
Fejfar (1987) reported 28 molars and premolars and 40 incisors of phiomyids from Oligocene deposits near the Zallah Oasis, southern Libya. The rodents and large mammals from these deposits suggested correlation with the Jebel Qatrani Formation, Fayum, Egypt. The Libyan rodents include Metaphiomys schaubi, Phiomys andrewsi, and Phiomys paraphiomyoides, with Metaphiomys the dominant taxon. Fejfar (1987) illustrated, but did not describe the specimens. Holroyd (1994, p. 324) disagreed with the identification of Phiomys andrewsi and suggested that M. schaubi was synonymous with M. beadnelli.
Phiomyids are known from two early Oligocene localities in Oman (Thaytiniti and Taqah; Thomas et al. 1989; 1999). Thomas et al. (1999) provided the most recent faunal list, but did not describe or illustrate any specimens. Holroyd (1994, p. 326) suggested a late Eocene age for the sites. The phiomyid fauna from Thaytiniti includes Phiomys cf. P. andrewsi, P. cf. P. lavocati, and cf. Metaphiomys spp. 1 and 2. The phiomyid fauna from Taqah includes those species plus M. cf. M. schaubi (M. sp. in Thomas et al. 1989).
Jaeger et al. (1985) described Protophiomys algeriensis based on 23 isolated teeth from the late Eocene Bir el Ater (= Nementcha) locality, eastern Algeria. Holroyd (1994) suggested that the hypodigm of Protophiomys algeriensis may actually include more than one taxon.
Stevens et al. (2004) reported multiple phiomorph rodent taxa from a site in the Mbeya Region, southwestern Tanzania. They noted that these taxa suggest a late Paleogene age for the locality. An isolated incomplete lower molar of Metaphiomys cf. M. beadnelli was described (Stevens et al., in press).
Family PHIOMYIDAE
Wood, 1955
Ugandamys, gen.
nov.
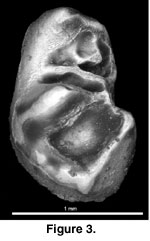
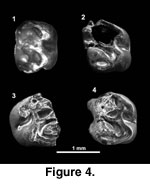
Figure 3,
Figure 4
 Type and only species. Ugandamys downsi.
Type and only species. Ugandamys downsi.
Diagnosis. As for type and only species.
Etymology. “Uganda,” the country where it is found plus “mys,” Greek for mouse.
Ugandamys downsi, gen. and sp. nov.
Holotype. BUMP 1022, right Dp4.
Referred material. BUMP 1023, RM1 or M2; BUMP 1024, LM1 or M2; BUMP 1025, RDP4. All from Bukwa IIB.
Diagnosis. Small size; proportionally elongate teeth. Dp4 – trigonid narrower than talonid; preprotocristid absent (primitive); metaconid opposite protoconid (derived); protoconid significantly reduced in size relative to metaconid; short metalophid; ectolophid present (derived) and relatively labial in position; strong mesolophid originating from the ectolophid and extending transversely to the lingual border of the tooth (derived); mesoconid present (primitive); triangular shaped talonid with a deep circular basin. Upper fourth premolar and M1 or M2s – strong mesoloph present.
Type locality and age. Green clay unit, Bukwa IIA, 4.0 km east of the village of Bukwa, Kapchorwa District, eastern Uganda (34°47’East longitude, 1°17’North latitude); early Miocene, about 22 Ma (Walker 1969).
 Etymology. “downsi” for Will Downs who
recovered the specimen; in gratitude for his dedication to the recovery of
small mammal fossils from Uganda and elsewhere.
Etymology. “downsi” for Will Downs who
recovered the specimen; in gratitude for his dedication to the recovery of
small mammal fossils from Uganda and elsewhere.
Description. Tooth terminology for the Bukwa rodents is adapted from Flynn et al. (1986). Polarity of character states is from Holroyd (1994, table 4.14).
Lower Fourth Premolar
The Dp4 is 1.44 mm long x 0.92 mm wide. This tooth is long and slender, with the trigonid narrower than the talonid. The mesial edge of the tooth is incomplete, but there appears to be a very low indistinct shelf along the metaconid. The metaconid occupies much of the mesiolingual portion of the tooth. It is the second largest cusp. The protoconid is significantly reduced. Some enamel along its mesiolabial edge is missing, but even if complete the protoconid would still be much smaller than the metaconid. The protoconid and metaconid are adjacent to each other; i.e., the metaconid is not anterior to the protoconid. Short low crests extend distally from the more medial aspects of the metaconid and protoconid. Each crest contacts a cuspule. These cuspules are joined as a short metalophid, and the lingual cuspule has a tiny crest continuing distally to the mesial side of the mesolophid, just lingual to its midpoint. A low crest projects distally from the metaconid to contact the entoconid, which is the largest cusp on the tooth. The ectolophid is low and extends distally from the protoconid along the labial side of the tooth. It curves slightly lingually to contact a long mesolophid, which continues transversely to the crest along the lingual side of the tooth. This crest is called a mesolophid (versus the posterior arm of the protoconid, protospur, or metalophulid II) because of its close proximity to the mesoconid. Continuing distally from the mesolophid, the ectolophid contacts a small mesoconid at the junction with the hypolophid. The hypolophid joins the entoconid to the hypoconid; the latter is the third largest cusp on the tooth. The hypoconid is staggered distolabial to the entoconid. There is a strong mesially concave posterolophid connecting the hypoconid and entoconid. At the distal apex of the posterolophid the enamel is thickened and likely represents a hypoconulid. These three cusps (plus the mesoconid) and their associated lophs define a roughly triangular talonid, which borders a deep circular basin. Roots of the Dp4 are not preserved.
Upper Fourth Premolar, M1, and M2s
BUMP 1023 (M1 or M2) is 1.44 x 1.16 mm and BUMP 1025 (DP4) is 1.52 x 1.20 mm. BUMP 1024 is too incomplete to measure but is similar in size to BUMP 1023 and 1025. These teeth are approximately rectangular in occlusal outline and longer than wide; BUMP 1025 (DP4) is proportionally slightly longer. The mesial and distal sides are gently convex. None of the teeth are complete. The delicate crown of BUMP 1025 (Figure 4.1-4.2) was originally complete but was damaged during preparation for SEM study. Much of the detailed structure of the occlusal surface of BUMP 1024 (Figure 4.3) is obscured by breakage.
These teeth have four distinct cusps, plus other minor cusps, and five crests. The lingual side of the occlusal surface has worn more quickly than the labial side. Thus, on BUMP 1023 (Figure 4.4) and 1025 the lingual side is relatively flattened, but the major labial cusps (paracone, metacone) are still prominent and bulbous. The protocone and hypocone are crescent shaped with occlusal wear. The protocone-paracone and metacone-hypocone pairs are approximately opposite each other. The paracone is larger than the metacone, and the protocone is slightly larger than the hypocone. An oblique anteroloph extends from the protocone to the labial side of the tooth. On BUMP 1025 this crest has two cuspids; the more lingual is heavily worn. There is a short protoloph connecting the protocone to the longitudinal crest. This short longitudinal crest, just lingual to the midline of the tooth, connects the protocone to the metaconule, and then continues from the metaconule to the hypocone. The metaconule is smallest on BUMP 1025, but more distinct on BUMP 1023 and 1024. The mesoloph is long and slightly oblique. It originates from the metaconule and continues to the labial side of the tooth where it ends in a mesostyle on BUMP 1024 and 1025. This area of the tooth is damaged on BUMP 1023. A distinct posteroloph extends from the hypocone to the labial side of the tooth, contacting the metacone. There is a hypoconule at the junction of the posteroloph with the metaloph on BUMP 1025. A short but distinct metaloph is present on BUMP 1023 and 1025. A metaloph is lacking, or at most represented only by an extremely low indistinct crest, on BUMP 1024.
There is a strong lingual root supporting both the protocone and hypocone. Separate smaller roots are present under the paracone and metacone.
Ugandamys downsi is best placed within the Family Phiomyidae. Holroyd (1994) recently proposed an emended diagnosis of the Phiomyidae (originally diagnosed by Wood 1955; emended by Wood 1968). Her revised diagnosis uses characters of the Dp4 to distinguish it from the Dp4 of the Family Thryonomyidae: 1) generally smaller size; 2) Dp4 replaced by p4 primitively; 3) metaconid less anteriorly placed; 4) relatively stronger hypolophid; and 5) less lingually placed ectolophid. It is unknown if Ugandamys replaced the Dp4. The metaconid of the Dp4 of Ugandamys is adjacent to the protoconid, similar to the condition in the Phiomyidae, and unlike the anteriorly placed metaconid of the Thryonomyidae. The hypolophid of the Dp4 of Ugandamys is distinct, and the ectolophid is along the labial border of the tooth. The small size and presence of a mesoloph on the DP4 and M1 or M2s is consistent with assignment to the Phiomyidae.
Holroyd (1994, p. 97) proposed placement of the genera Diamantomys, Metaphiomys, and a new genus (and excluding Pomonomys) into the subfamily Diamantomyinae (new rank, rediagnosed), within the Family Phiomyidae. The Diamantomyinae were distinguished in part from the Phiomyinae (new rank, rediagnosed by Holroyd 1994, p. 83) by their larger size and consistently having a mesolophid. The diamantomyine Dp4 has the protoconid and metaconid aligned labiolingually, a preprotocristid consistently present, and a relatively wider trigonid. The Dp4 of Ugandamys has a strong mesolophid, the protoconid and metaconid are aligned labiolingually, and size is close to, but still smaller than, that of Holroyd’s (1994, table 4.6) proposed new genus. Although the Dp4 of Ugandamys lacks a preprotocristid, and has a relatively narrow trigonid, the Dp4 of Ugandamys fits best within Holroyd’s proposed Subfamily Diamantomyinae. Based on their strong mesoloph and size close to that of Holroyd’s (1994, table 4.6) proposed new genus, the BUMP DP4 and these two upper molars are also best considered within this subfamily.
Dp4: Of the published phiomyids, the Bukwa Dp4 is most similar to, but still distinct from, the morphology represented by the Dp4 of AMNH 13271, “Phiomys andrewsi,” from the Fayum (Wood 1968, figure 1G). As originally noted by Wood (1968), and supported by Holroyd (1994), the original hypodigm of “Phiomys andrewsi” actually consists of more than one taxon. Holroyd (1994) suggested that material from the original hypodigm, plus additional specimens, represent the genus Phiomys (emended diagnosis) and a new unpublished genus (Holroyd 1994, p. 102). AMNH 13271 was considered by Holroyd (1994) to belong to the new genus, within her Subfamily Diamantomyinae.
Like AMNH 13271, the Bukwa Dp4 is elongate, has the metaconid and protoconid opposite with a metalophid connecting them posteriorly. The mesolophid is long and extends to the lingual side of the tooth. In comparison with AMNH 13271, BUMP 1022 has a distinctly smaller protoconid, lacks a preprotocristid, probably has a more labially placed ectolophid, has a mesoconid, and has a triangular talonid defining a circular basin. In addition, BUMP 1022 is from 7 to 17% shorter (N of the new genus is 6 to 16 for two species) and from 18 to 22% narrower (N = 6 to 18) than the mean values given by Holroyd (1994, table 4.7) for the genus to which AMNH 13271 belongs. Compared to species of the genus Phiomys from the Fayum (Holroyd 1994, table 4.2), Ugandamys is 10% shorter to 17% longer (N=1 for each of two species, 3 for a third species), and ranges from 23% narrower (N=4), comparable in width (N=2), to 24% wider (N=1).
Ugandamys is distinct from a Dp4 illustrated, but neither specifically named nor described, from prior collections at Bukwa (Lavocat 1973, p. 158, figure 28.7; specimen B9). Lavocat (1973, p. 158) assigned his right Dp4 to “Paraphiomys stromeri of small size.” Compared to Lavocat’s specimen (B9), BUMP 1022 is comparable in length but is slightly narrower. Approximate measurements of B9 from Lavocat’s photograph are 1.5 x 1.1 mm. On BUMP 1022 the protoconid is much smaller than the metaconid; these cusps are of more comparable size on B9. On B9 the area between the protoconid/metaconid and hypoconid/entoconid is obscured, so it is unknown if there is a metalophid or mesolophid. The strong labial ectolophid observed on BUMP 1022 is absent, or is more medial in position and obscured on B9. Unlike Ugandamys, B9 has a gently concave distal end, and instead of a circular basin the talonid has a narrow valley, which opens lingually.
Lavocat (1973) described two taxa of phiomyids from the early Miocene of Kenya. Andrewsimys parvus was reported from only two incomplete maxillae from Songhor; the lower dentition is unknown. Phiomys andrewsi was described from a skull, upper jaw, and mandibles from Songhor, Rusinga, and Koru. Holroyd (1994) considered A. parvus to be a valid genus within the Phiomyidae. Although she accepted the Kenyan “Phiomys andrewsi” in the Phiomyidae, she thought it distinct at both the generic and specific levels.
Unfortunately, Lavocat (1973, figure 28.4) did not illustrate the Dp4 of the Kenyan “Phiomys andrewsi,” although the m1-m3 are shown. His measurements for the Dp4 of one specimen, SO 879, are 1.55 x 1.45 mm. BUMP 1022 is slightly shorter than the Dp4 of SO 879, but the Bukwa tooth is significantly (63%) narrower. It is noteworthy that there are strong mesolophs on the lower molars, although they extend only halfway across the m1. Posterolophids contact the entoconid, enclosing a mesiodistally compressed basin on m1 and a more transverse valley on m2 and m3.
Fejfar (1987) illustrated four Dp4s of Metaphiomys schaubi (Dp4s not reported for the other taxa) from Zallah, Libya (Fejfar 1987, figure 1.1-1.4). The Zallah Dp4s (smallest is about 2.3 x 1.6 mm; Fejfar 1987, figure 2) are all appreciably larger than BUMP 1022. Like BUMP 1022, the protoconid and metaconid are opposite, but these cusps are comparable in size on the Libyan specimens. Compared to Ugandamys, the Zallah specimens also differ in at least three of them having a distinct preprotocristid. The specimens also have a small to absent mesolophid, a more lingually placed ectolophid, and a proportionally wider trigonid. Three of four specimens from Zallah have a triangular talonid defining a circular basin, similar to Ugandamys.
The rodent fauna from Thaytiniti, Oman, includes two Dp4s referred to Phiomys cf. P. andrewsi, which are illustrated (one specimen) and briefly described by Thomas et al. (1989). Holroyd (1994, p. 85) did not consider this material referable to P. andrewsi. The one measurement given by Thomas et al. (1989), 1.45 x 0.94 mm, is comparable to that of Ugandamys. Ugandamys differs from the Oman tooth in that the later has the protoconid and metaconid of comparable size, the protoconid has a distinct mesio-internal projection, the metalophid is stronger, the ectolophid is more oblique and more lingual, the hypolophid is longer, and the hypoconulid larger. The mesolophids cannot be adequately compared with the available illustration of the Oman specimen.
BUMP 1022 is distinct from the Dp4 of Protophiomys algeriensis from Algeria (Jaeger et al. 1985). Compared to BUMP 1022, the Dp4 of Protophiomys algeriensis is proportionally wider (mean measurements 1.60 x 1.73 mm, N = 3), has a more anterior and proportionally smaller metaconid, lacks a metalophid connecting the metaconid and protoconid, lacks a mesolophid, has a more lingual ectolophid, lacks the triangular talonid, and has a stronger hypoconulid.
DP4 and M1 or M2s: Compared to the Fayum phiomyids (Wood 1968; Holroyd 1994), the DP4 and M1 or M2s of Ugandamys are most similar to Holroyd’s (1994, e.g., figure 4.7, DPC 8170) proposed new genus. The new genus includes three species; lengths of the BUMP specimens are within the range of the Fayum material (Holroyd 1994, table 4.6), but the BUMP specimens are proportionally slightly narrower. The mesoloph is usually present on the species represented by DPC 8170 but is variable in development. It may be as strong as the consistently long mesoloph in Ugandamys. The species represented by DPC 8170 does not have a distinct metaconule, but Ugandamys has a small to distinct (2 of 3 specimens) metaconule. An incomplete metaloph is also present on Holroyd’s (1994) new genus, but it may be stronger and join the area of origin of the mesoloph in addition to joining the posteroloph.
Lavocat (1973, plate 28.6, 28.8, 28.9) illustrated, but did not describe, two isolated small thryonomyoid upper molars from Bukwa. Plate 28.8 (B12) is a right upper molar referred to a thryonomyoid “a little smaller than Paraphiomys stromeri of small size and of a form a little different” (Lavocat 1973, p. 158; translated from French). Figure 9 (B11) is a “left upper molar of P. stromeri of small size with the mesoloph reaching the border of the crown” (Lavocat 1973, p. 158; translated from French). Measurements of these specimens are not given, but approximate measurements from the Plate are B12, 1.40 x 1.20 mm and B11, 1.60 x 1.70. It is probable that these teeth belong to the same species, with the differences ascribable to intraspecific variation. B12 is comparable in size to BUMP 1023 (1.44 x 1.16 mm) and BUMP 1025 (1.52 x 1.20 mm). B11 is comparable in length but wider. None of the BUMP specimens have the lingual side of the tooth as proportionally short compared to the labial side, as seen on B11. Overall morphology of B11 and B12 and Ugandamys is similar. Especially notable is the long mesoloph, which extends to the labial border of the crown (B11 also has a mesostyle) and a short metaloph. The anteroloph is better developed on B11 than B12. The anteroloph on B11 is comparable to that of BUMP 1025, the DP4. The labial part of both BUMP 1023 and 1024 is damaged, so detailed comparisons of the anteroloph are not possible. Definitive presence or absence of a metaconule cannot be determined from the photographs of B11 and B12. It is possible that Lavocat’s (1973) B11 and B12 are referable to Ugandamys, with the differences related to intraspecific variation.
Lavocat (1973, plate 41.6) described and illustrated the upper dentition of “Phiomys andrewsi” from Songhor, Kenya. He noted the strong development of the mesoloph, a distinct mesocone (= metaconule in the terminology used here), and a short metaloph. Lavocat (1973, table 14) provided measurements for one upper dentition (specimen number not given): DP4 (P4 of Lavocat) 1.6 x 1.7 mm, M1 1.6 x 1.75 mm, M2 1.5 x 1.7 mm. These teeth are proportionally wider than Ugandamys (acknowledging that it is unknown if the BUMP specimens are M1 or M2s). Length to width ratios for his specimens are DP4, 0.94; M1, 0.91; M2, 0.88. Length to width ratio for BUMP 1025 DP4 is 1.27, BUMP 1023 is 1.24. The posterior portion of Lavocat’s “P. andrewsi” differs significantly from Ugandamys in formation of a circular basin bounded mesially by a connection between the metaloph and the metaconule or the mesoloph near where it contacts the metaconule.
Lavocat (1973, plate 42.4) also described and illustrated two incomplete maxillae of Andrewsimys parvus from Songhor, Kenya. This taxon is diagnosed by its small size and distinct cusps. The lophs (e.g., the mesoloph) are reduced, although the mesoloph looks strong on the photograph of “P4” (Lavocat 1973, plate 42.4). A metaconule and short metaloph are present. Lavocat considered the specimens to have P4s, not DP4s, although he was uncertain. His “P4s” of A. parvus are 1.1 x 0.95 mm and 1.15 x 1.1 mm; the M1s are 1.2 x 1.1 mm and 1.15 x 1.25 mm (Lavocat 1973, table 14). If Lavocat’s specimens are DP4s, they are about 27% shorter and proportionally wider (length to width ratios 1.04 and 1.16) than BUMP 1025. The M1s of Andrewsimys parvus are about 18% shorter and proportionally wider (length to width ratios 1.09 and 0.92) than the M1 or M2 (BUMP 1023) of the BUMP taxon.
The sample of phiomyids from Zallah Oasis, Libya (Fejfar 1987), did not include any DP4s. Two M1 or M2s of Metaphiomys schaubi were illustrated (Fejfar 1987, figure 1.19, 1.20). Measurements for three specimens of this taxon are significantly larger (Fejfar 1987, figure 2; smallest specimen about 2.31 x 2.65 mm) than Ugandamys, and no further comparison is given here. An upper molar of Phiomys paraphiomyoides was also illustrated (Fejfar 1987, figure 1.17; no tooth position given, but likely M1 or M2). This specimen is closer in size (about 1.45 x 1.4 mm from Fejfar 1987, figure 2) to BUMP 1023. Phiomys paraphiomyoides was diagnosed (in part) by Wood (1968, page 41) by the metaloph connecting with both the posteroloph and mesoloph. This connection appears to be present in Fejfar’s specimen, which also has a very short mesoloph and is squarer in outline than Ugandamys.
Thomas et al. (1989) illustrated and briefly described the upper molars of some phiomyids from Thaytiniti and Taqah, Oman. Phiomys cf. P. lavocati was reported from both sites (total of three specimens). This material differs significantly in smaller size from Ugandamys (e.g., M1 from Taqah 0.97 x 1.02 mm) and squarer outline, shorter or interrupted mesoloph, and connection of the metaloph to the posteroloph and mesoloph/metaconule, helping form a circular basin in the Oman material. Thomas (1989, figure 11.a) also referred an M1 to cf. Metaphiomys sp. 1. Holroyd (1994, page 102) considered this specimen to belong to her proposed new genus of phiomyid. The M1 of cf. Metaphiomys sp. 1 (1.52 x 1.66 mm) is comparable in length but wider than BUMP 1023. Like Ugandamys, the Oman tooth has a metaloph, metacone (= metaconule), and a strong mesoloph reaching the labial wall. However, the M1 of cf. Metaphiomys sp. 1 has a more complicated occlusal pattern, including a short crest between the anteroloph and the protoloph, a distal elongation of the protocone, and a metaloph connecting to both the mesoloph and the posteroloph (in part forming a circular basin).
The description of Protophiomys algeriensis from Algeria (Jaeger et al. 1985) included three DP4s and six M1 or M2s. The morphological variability of the illustrated M1 or M2s (Jaeger et al. 1985, plate 1.6-1.8) may reflect presence of more than one taxon. Thus, comparisons of this taxon with the Bukwa upper cheek teeth would be misleading. However, it should be noted that the diagnosis of Protophiomys algeriensis includes upper molars lacking a mesoloph (strong on Bukwa specimens), and the metaloph never connected to the posteroloph (always connected on Bukwa specimens).
Family Thryonomyidae
Pocock,
1922
cf. Paraphiomys
sp.
Figure 5
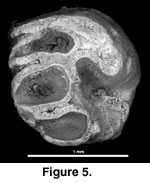 Referred material. BUMP 1026, right upper cheek
tooth, from Bukwa IIA.
Referred material. BUMP 1026, right upper cheek
tooth, from Bukwa IIA.
Description. BUMP 1026 is incomplete, especially along the labial side. Minimum measurements are 1.4 x 1.2 mm. This tooth is etched, suggesting digestion or the effect of postdepositional soil acids. Some smooth polishing of the sides of the tooth suggests postmortem transport. There are four major nearly transverse lophs: anteroloph, protoloph, metaloph, and posteroloph. The third loph is considered a metaloph, because it goes directly to the metacone and does not seem to originate from a central cusp (metaconule). BUMP 1026 lacks the strong mesoloph seen on the other upper molars described here from Bukwa. The protoloph and metaloph connect to a longitudinal crest located lingual to the midline of the tooth. All cusps are worn flat, and the hypocone is completely missing (broken). A strong root is present under the protocone. Another root is present under the hypocone and likely also under the distolabial aspect of the tooth. The rest of the underside of the tooth is damaged.
Discussion
The occlusal pattern of this tooth (e.g., four nearly transverse lophs) is comparable to a small species of Paraphiomys (i.e., P. stromeri size or smaller), but incompleteness of the specimen precludes specific assignment. Paraphiomys is a common component of early Miocene and, to a lesser extent, middle Miocene African faunas (e.g., Lavocat 1973; Winkler 1992; Mein et al. 2000, López Antoñanzas et al. 2004). Paraphiomys simonsi was reported from the Fayum of Egypt (Wood 1968). Holroyd (1994) proposed transferring this species to a new genus, but López Antoñanzas et al. (2004) retained it within Paraphiomys. Whatever the allocation of P. simonsi, the Bukwa tooth is smaller and has a strong metaloph (metaloph lacking in P. simonsi; Holroyd 1994 and López Antoñanzas et al. 2004).
![]()