 Holotype: Left M1 (V 14177.1)
Holotype: Left M1 (V 14177.1)
Pararhizomys qinensis new species
(Fig. 1.2,
1.5, Fig. 2;
Table 1)
Holotype: Left M1 (V 14177.1)
Paratypes: Right M2 (V 14177.2) and a partial skull (IVPP V 14176).
Type locality: Lantian, Shaanxi Province, Loc. 13, lower part of the Bahe Formation.
Diagnosis: Snout robust, deeper than in Pararhizomys hipparionum, but with a shorter diastema. Tooth dimensions less than 90% of Pararhizomys hipparionum. M1 and M2 with less undulating enamel base on both labial and lingual sides, and shorter striae.
Etymology: “qin,” Chinese Mandarin spelling for central Shaanxi Province and the Ancient dynasty.
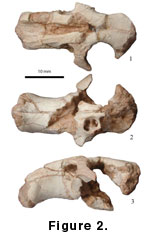
Description: The partial skull from Loc. 13 has only the premaxillae, the nasals, the maxillae with the anterior part of the left zygomatic arch and the alveolar surface for right M1 preserved, the frontals and portions of parietals. The snout is short and strongly built, with a short diastema (16.5 mm from incisors to M1 alveolus). The origin of the anterior part of the lateral masseter is on the maxillary portion of the zygomatic arch, under the large infraorbital foramen, and just posterior to the premaxilla-maxilla suture. The zygoma shows modified myomorphy, as do fossil rhizomyids (Flynn 1982) in that the ventral slit of the keyhole infraorbital foramen is constricted, nearly closed, with the larger part of the foramen shifted dorsally. The anterior margin of the incisive foramen is at the middle point of the diastema.
M1 (V 14177.1) is high crowned, cylindrical, and slightly recurved anteriorly in lateral view. On the occlusal surface there are one lingual and two labial reentrants that yield a sigmoid outline, which recalls the M2 of siphneines. The lingual reentrant extends anteriorly to the central longitudinal axis. The anterior labial reentrant bends slightly anteriorly and extends to just in front of the deepest part of the lingual reentrant. The posterior labial reentrant is deepest and extends posterolingually beyond the midline of M1. The base of the enamel approaches closest to the roots anteriorly. On the lingual side, the reentrant terminates well short of the base of the enamel, not quite as far on the labial side. Two roots can be seen at the base of the tooth, the anterior one much larger.
M2 is smaller than M1. There are only one labial and one lingual reentrant, which yield a general “S” shape. The lingual reentrant is shorter and bends gently anteriorly; the labial one extends posterolingually beyond the central longitudinal axis. Anteriorly, the lingual corner of the tooth is sharply triangular. Anterior and posterior walls of M2 are flattened; the anterior width is larger. Two roots are observed.
Comparison of Species:The single
specimen of Pararhizomys hipparionum from its type locality has
only lower molars, plus the incisor. Fortunately, the complete skull with lower
jaws from Altan Teli allows comparison among all fossils attributed to the
genus. The Altan Teli lower dentition is very similar to that of the holotype,
differences (Table 1) due to more advanced wear of the Altan Teli individual.
 The upper dentition of the cranium from Altan Teli resembles that of the Fugu
maxilla, but dimensions are greater, again reflecting its advanced wear stage
(occlusal size of M1 is considerably greater, but the less worn M3 is nearly the
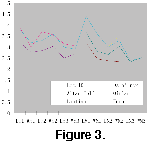
same in the two specimens). Figure 3 shows these size differences in M1, the
less contrasting M2, and the similar M3.
The upper dentition of the cranium from Altan Teli resembles that of the Fugu
maxilla, but dimensions are greater, again reflecting its advanced wear stage
(occlusal size of M1 is considerably greater, but the less worn M3 is nearly the
same in the two specimens). Figure 3 shows these size differences in M1, the
less contrasting M2, and the similar M3.
The importance of considering wear stage cannot be overemphasized. Molar size is, of course, constant during an individual’s life, but measurements, usually made with respect to the occlusal surface, are affected by wear. In Pararhizomys, like many high crowned rodents, some occlusal dimensions increase greatly with wear. The wear surface of upper molars increases dramatically in width during life; lower molars, especially m3, show increase in length. Some researchers measure only occlusal surfaces, as did, apparently, Teilhard and Young (1931). We recorded maximum crown dimensions and remeasured a cast of the holotype of P. hipparionum (Table 1). Note that measurements in late wear could exceed our estimates, due to ontogenetic shift of the angle of the wear surface with respect to the base of the enamel; holotype m1 length could approach 4.0 mm, and m3 length could exceed 3.5 mm. Given this consideration, the worn Altan Teli specimen is quite consistent with the Fugu specimens.
However, this logic suggests that the lower molars from Qin’an, which appear to be smaller than those of the holotype, have achieved nearly the same wear stage and are perhaps too small for this species allocation (Table 1, Figure 1.3). Apparent size of the molars in the Qin’an jaw would increase in more advanced wear, but it is at almost the same wear stage as the holotype. This specimen appears small for Pararhizomys hipparionum, but being lower dentition, it is difficult to compare with the Lantian fossils.
The upper molars from Lantian show significantly smaller size of Pararhizomys qinensis than Pararhizomys hipparionum. Dimensions of the specimens from Lantian are less than 90% those of the upper dentition from Fugu, including molar length, which would not change much with wear (possibly decreases). Width of M2 in V 14177.2 is less than 80% that of the Fugu maxilla. These specimens, approximately at the same wear stage, show different vertical penetration of reentrants in M2; the striae (lateral views of reentrants) do not approach the base of the enamel as deeply as in the Fugu specimen.
The Lantian skull (Figure 2) has the same modified myomorphous zygomasseteric structure as that of the Altan Teli specimen. The origin of the anterior part of the lateral deep masseter is on the maxillary portion of the zygomatic arch, which expands anteriorly, and the infraorbital foramen is in a high position with abbreviated ventral slit. The Altan Teli skull is larger; its diastema between the upper incisor and M1 is 24.5 mm (16.5 in V 14176). The snout is relatively longer and less deep dorsoventrally in the Altan Teli specimen, lending a robust appearance for the Lantian skull. Despite the difference in size, the Altan Teli upper molars have almost exactly the same structures as those from Lantian, although the deeply worn lingual and anterior labial reentrants are enclosed as enamel lakes. The M2/M1 length ratio for Lantian is 0.74, in contrast to 0.68 for the Altan Teli individual.
The two species of Pararhizomys are similar in molar and skull structure. Differences include smaller size of Pararhizomys qinensis (usually presumed as a primitive condition), less undulating base of the enamel (primitive), and shorter striae (considered primitive in other high crowned rodents, such as siphneines). M2 is relatively shorter, slightly more reduced (derived), in Pararhizomys hipparionum, based on the few specimens available.
Summary: With the Fugu maxilla V 14178 as a standard for Pararhizomys hipparionum, the Lantian specimens are proportionally smaller in size (Figure 3) and with a less undulating base of the enamel on both labial and lingual sides. The striae are longer in P. hipparionum. Considering these dental features and the more robust snout with shorter diastema, new species status is proposed for the Lantian specimens. The species Pararhizomys qinensis is considered more primitive and likely older than P. hipparionum. Both species show the derived zygomatic condition of modified myomorphy.
The large body size, molar lophodonty, high crowns, and simple tooth structures suggest comparison of Pararhizomys with the muroid rodent groups Tachyoryctoididae, Siphneinae, Spalacidae, and Rhizomyidae. Members of all of these (with the possible exception of the extinct Tachyoryctoididae) show fossorial adaptations in the humerus, incisor morphology, and occipital and cervical structures. Such adaptations are not definitively expressed in known specimens of Pararhizomys.
Tachyoryctoididae: Tachyoryctoides occurs widely across China and Kazakhstan, for example in the Oligocene deposits of Shargaltein-Tal and Taben-Buluk, Gansu Province (Bohlin 1937, 1946), and the early Miocene of Xining and Lanzhou basins (Li and Qiu 1980). The mandibles are strongly built, with prominent masseteric crests under m1/m2 as in Pararhizomys. A cranium of Tachyoryctoides (V 5999) from Xining Basin, has the anterior part of the lateral masseter extending below the infraorbital foramen. The lower cheek teeth all have two labial reentrants, reminiscent of m1 and m2 of Pararhizomys. The posterior reentrants are much deeper and more strongly posteriorly oriented, unlike the shallower and more transverse reentrants in Pararhizomys. The lingual reentrants on upper cheek teeth are strongly anteriorly oriented and opposite to the anterior labial reentrants in Tachyoryctoides. We see no special features shared to support phylogenetic relationship with Pararhizomys.
Siphneinae: The hypsodonty and simple tooth morphology of Pararhizomys are superficially consistent with siphneines, the living fossorial zokors of China. Recent study shows that the group is probably derived from middle Miocene Plesiodipus (Zheng et al. 2004). The molar pattern differs significantly in some respects, for example the simple, small third molars of Pararhizomys, in contrast to the large, complex posterior teeth of the zokors. Similarities are likely shared primitive conditions (two labial reentrants on m2) and convergences.
Spalacidae: Extinct spalacids are grouped in four genera: Debruijnia, Heramys, Sinapospalax, and Pliospalax (Ünay 1999; Sarica and Sen 2003). The fossil and extant spalacids are restricted to western Asia, southeastern Europe, and northern Africa. Mesoloph(id)s are present in primitive genera and reduced in later genera. There is no lingual reentrant on M3, except for Debruijnia having a shallow and narrow reentrant. The labial anterior reentrant in Spalacidae diminishes from very shallow in early taxa to being absent in extant species. The labial posterior reentrants are shallow and strongly posteriorly orientated. These features argue that Pararhizomys is related to neither primitive nor derived genera. Flynn (1982) felt that the rounded infraorbital foramen of Spalax was more consistent with hystricomorphy than myomorphy and that the Altan Teli skull appeared to be hystricomorphous. Re-examination of the Altan Teli specimen and observations on the new Lantian skull affirm that Pararhizomys exhibits modified myomorphy.
Rhizomyidae: Echoing Teilhard and Young (1931), McKenna and Bell (1997) grouped Pararhizomys with the living Rhizomys and Cannomys, plus extinct Anepsirhizomys and Brachyrhizomys, in the tribe Rhizomyini. There are indeed some characters shared with these taxa, such as incisors deeply rooted with well-developed capsules, lophodont cheek teeth, and modified myomorphy with anterodorsally shifted origin of the lateral masseter, which restricts and partially closes the ventral slit of the infraorbital foramen. However, Pararhizomys differs from most rhizomyines in its reduced third molars, extra anterolabial reentrant on m2, and otherwise simplified tooth morphology. Flynn (1990) excluded Pararhizomys from Rhizomyidae based on several features and thought the most important character to be the lower second molar with two labial reentrants. The extra anterior labial reentrant in m2 of Pararhizomys could result from a hypertrophied labial arm of the muroid anterolophid, that is, a condition derived from a primitive muroid feature, present in early rhizomyids, and therefore not a falsification of classification in Rhizomyidae. Also, the new Lantian skull confirms that the zygomatic structure of Pararhizomys shows a condition of modified myomorphy, consistent with fossil rhizomyids. Further, the retained ridge on the lower incisor of Pararhizomys is a feature seen in early rhizomyids (and retained in tachyoryctines). Other common features could represent shared ancestry and parallel evolution.
While there is no compelling case for assignment of this genus to any living family, we consider two likely scenarios. Pararhizomys may be a rhizomyid, as Teilhard and Young (1931) postulated, but it would not be closely related to modern rhizomyines such as Brachyrhizomys shansius. Instead, Pararhizomys may be derived from earlier forms retaining a distinct labial arm of the anterolophid on m2 and the ridge on the lower incisor. The second possibility is that Pararhizomys is a late-surviving member of an independent muroid lineage. It remains for more fossils to be discovered in China to clarify the affinity of Pararhizomys and to determine whether this taxon represents a separate muroid lineage, heretofore unrecognized.
Lantian Loc. 13, which is located about 20 m above the rich small mammal site Loc. 19, produced Progonomys sinensis (Zhang et al. 2002; Qiu et al. 2004a) and Abudhabia baheensis (Qiu et al. 2004b). Loc. 19 yielded additional taxa (Qiu et al. 2003). Together with Loc. 12, these three sites document the earliest fauna in the Bahe Formation. Preliminary paleomagnetic data (Kaakinen and Lunkka 2003) and faunal comparisons (Zhang et al. 2002; Qiu et al. 2003; Qiu et al. 2004a) argue that the fossil mammals from the lower part of Bahe Formation can be compared with European Vallesian faunas. Thus the type locality for Pararhizomys qinensis is early late Miocene, perhaps around 10 Ma.
Pararhizomys hipparionum is advanced with respect to Pararhizomys qinensis in greater size, longer striae, and more undulating base of the enamel. Consistent with the derived nature of this species, Pararhizomys hipparionum evidently is found in younger localities. The Fugu maxilla V 14178 is from Lamagou, which produced abundant mammalian fossils (Xue et al. 1995) and has a paleomagnetic age of about 8 Ma. The Fugu type locality “Loc. 10,” as previously discussed, is of uncertain age, but seems to be later based on greater size of the holotype of Pararhizomys. The fossil level of the Qin’an material at about the 9 m level in the first section of Guo et al. (2002) has a projected paleomagnetic date of ca. 7 Ma. Altan Teli and Dalai Nor both might be latest Miocene. The more advanced Pararhizomys hipparionum is apparently from younger deposits than Pararhizomys qinensis.
![]()