 Holotype. Target Pan 3a'93 housed at the Geological Survey of Namibia, Windhoek.
Holotype. Target Pan 3a'93 housed at the Geological Survey of Namibia, Windhoek.
AUH: Abu Dhabi, specimens curated with the Abu Dhabi Islands Archaeological Survey, Abu Dhabi, United Arab Emirates. YPM: Peabody Museum of Natural History, Yale University, New Haven, Connecticut, USA.
Type oospecies. Diamantornis wardi Pickford and Dauphin 1993
Oogenus Diagnosis. Large eggs (16 x 13.3 cm), the external surface of which is patterned by large (2 to 8 mm) circular (in plan view), funnel-shaped (in section view) pore complexes (or 'megapores') (Pickford and Dauphin 1993). Regions of eggshell between the pore complexes are smooth and devoid of small pores; pore complex density ranges from zero to nine pore complexes per every 4 cm2; and eggshell thickness between the different oospecies of Diamantornis ranges approximately from 2.1 to 4.3 mm (Dauphin et al. 1996). Microstructural analyses confirm assignment of Diamantornis to the ratites (Dauphin et al. 1996).
Known distribution. Neogene Tsondab Sandstone aeolianites of the Namib Sand Sea, Namibia (Pickford and Dauphin 1993; Pickford et al. 1995). Lower Member of the Nawata Formation, Lothagam, Kenya (Harris and Leakey 2003; Harrison and Msuya 2005).
Holotype. Target Pan 3a'93 housed at the Geological Survey of Namibia, Windhoek.
Oospecies Diagnosis. Pore complexes circular in shape, having a diameter of about 5 mm (see below). These complexes tend to be well separated from each other by areas of smooth eggshell. Pore complexes are funnel shaped in section. Pore complex density is zero to four pore complexes per 4 cm2. Eggshell thicknesses range from 2.1 to 2.9 mm, mode = 2.4 mm (n=22) (Pickford et al. 1995, shell thickness means are unpublished).
The original diagnosis (Pickford et al. 1995) for this oospecies describes the pore complexes as having an approximate diameter of about 9.5 mm, a value inconsistent with the illustrations in the paper in which the pore complexes of D. laini eggshells appear to be closer to 5 mm in diameter. Later publications (Dauphin et al. 1996; Senut 2000) more accurately referred to D. laini pore complexes as being about 5 mm in diameter.
Known Distribution. As for Diamantornis above with the inclusion of D. cf. laini from Lothagam (Harrison and Msuya 2005).
New Material. Eighty-four eggshell fragments from the Late Miocene Baynunah Formation, Abu Dhabi Emirate, United Arab Emirates (AUH 922, 923, 926, 929–931, 934, 940–943, 947–952, 954–993, 995–1010, YPM 56709–56718).
Localities Present. Hamra (H3-1, H5-0), Baynunah Municipality Site (MU1-2), Ras al-Qal'a (Q1-1), Shuwaihat (S2-0, S2-1, S2-5, S3-0, S4-0, S8-1).
Description. The eggshell fragments vary in preservation. In general, the AUH specimens vary from greatly eroded to very well preserved. The 10 YPM specimens are greatly eroded. We consider highly weathered specimens to be those that exhibit rounding of broken edges, smoothing of pore complex rims, texturing of the surface between pore complexes, and exposure of the pore complexes on the internal surface of the eggshell. Fresh cross-sectional breaks clearly exhibit the abrupt transition between the mammillary and squamatic layers diagnostic of Palaeognathae (Grellet-Tinner 2000). Maximal fragment length ranges between about 1 cm and 6 cm. The eggshell fragments likely derive from multiple eggs and clutches, although it is a possibility that certain specimens may be from a single egg or a single clutch. The thickness of the eggshells ranges from 2.11 to 3.46 mm with a mean thickness of 2.77 mm (standard deviation 0.28). Pore complex diameter ranges from 1.43 to 6.17 mm with a mean diameter of 4.1 mm (66 measurements from 36 specimens; standard deviation 1.05). Mean pore complex density is 0.26 pore complexes/cm2 (73 specimens measured; standard deviation 0.20).
Fossil eggshell fragments of Diamantornis laini occur at almost all exposures of the Baynunah Formation in the UAE and in abundance relative to other faunal elements. These ratite eggshells from the Baynunah differ markedly in pore morphology and eggshell thickness from those of all modern ratites, including the genus Struthio. In terms of eggshell thickness, pore complex size and density, and overall morphology, the eggshell specimens conform well to the description of Diamantornis laini from Namibia. The Baynunah specimens exhibit an upper range of thickness exceeding that noted in the Namibian sample. Pore complex diameters range to just greater than the approximate measure in the Namibian sample. In all, the Baynunah specimens are metrically indistinguishable from specimens assigned to Diamantornis laini. Although the microstructure of D. laini has been described (Dauphin et al. 1996), no microstructural comparisons were made given the extremely characteristic macromorphology of these eggshells (cf. Stidham 2004; Harrison and Msuya 2005).
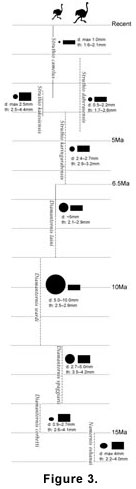 The occurrence of Diamantornis in southwestern Africa (Namibia), probably East Africa (Kenya), and now Arabia (UAE) is evidence for the existence in the latter half of the Late Miocene of at least a single form of large ratite bird distributed across this vast region. If D. laini dispersed to occupy its total geographic range soon after its origination and then only existed for a relatively short temporal span, then its presence would be of biostratigraphic significance. Pickford, Dauphin, and Senut described a sequence of ratite eggshells from the Neogene fossil record of Namibia that they interpreted as an anagenetic lineage spanning almost 20 million years (e.g.,
Senut et al. 1998).
Beginning with the ootaxon Namornis oshanai, originally described as
Struthio oshanai, and attributed a Pliocene age (Sauer
1966), their eggshell sequence progressed through four oospecies of
Diamantornis and two of Struthio to culminate in Struthio camelus,
the modern ostrich (Figure 3). " In total, two genera and seven species were named, each characterized primarily by discrete differences in eggshell thicknesses and pore complex widths and densities, and with no two forms displaying any overlap in temporal range. These workers proposed that ratite eggshell may be of relatively precise biostratigraphic utility (Senut and Pickford 1995;
Senut 2000).
Harrison and Msuya (2005) reported the temporal overlap of the oospecies Struthio cf. karingarabensis and Struthio kaesiensis at Laetoli, both of which would overlap with the suggested age range of the oospecies Struthio daberasensis (Senut 2000;
Stidham 2004). Further discoveries of eggshell from Miocene sites may determine that the different oospecies of Diamantornis also exhibited some temporal overlaps. In general, however, all the recent discoveries of fossil ratite eggshell have shown that the proposed biostratigraphic sequence of ootaxa is at least relatively accurate, even if the biochronology is imprecisely resolved.
The occurrence of Diamantornis in southwestern Africa (Namibia), probably East Africa (Kenya), and now Arabia (UAE) is evidence for the existence in the latter half of the Late Miocene of at least a single form of large ratite bird distributed across this vast region. If D. laini dispersed to occupy its total geographic range soon after its origination and then only existed for a relatively short temporal span, then its presence would be of biostratigraphic significance. Pickford, Dauphin, and Senut described a sequence of ratite eggshells from the Neogene fossil record of Namibia that they interpreted as an anagenetic lineage spanning almost 20 million years (e.g.,
Senut et al. 1998).
Beginning with the ootaxon Namornis oshanai, originally described as
Struthio oshanai, and attributed a Pliocene age (Sauer
1966), their eggshell sequence progressed through four oospecies of
Diamantornis and two of Struthio to culminate in Struthio camelus,
the modern ostrich (Figure 3). " In total, two genera and seven species were named, each characterized primarily by discrete differences in eggshell thicknesses and pore complex widths and densities, and with no two forms displaying any overlap in temporal range. These workers proposed that ratite eggshell may be of relatively precise biostratigraphic utility (Senut and Pickford 1995;
Senut 2000).
Harrison and Msuya (2005) reported the temporal overlap of the oospecies Struthio cf. karingarabensis and Struthio kaesiensis at Laetoli, both of which would overlap with the suggested age range of the oospecies Struthio daberasensis (Senut 2000;
Stidham 2004). Further discoveries of eggshell from Miocene sites may determine that the different oospecies of Diamantornis also exhibited some temporal overlaps. In general, however, all the recent discoveries of fossil ratite eggshell have shown that the proposed biostratigraphic sequence of ootaxa is at least relatively accurate, even if the biochronology is imprecisely resolved.
The age range of D. laini from Namibia has been determined as "late middle Miocene" (Senut et al. 1994), from 11 to 8 Ma (Senut and Pickford 1995), and from 10 to 8 Ma (Pickford and Senut 2000; Senut 2000). These estimates were based primarily on the stratigraphic occurrence of the D. laini material and its associated fauna. Pickford and Dauphin (1993) and Senut et al. (1994) listed Parapedetes namaquensis (Rodentia) and Giraffidae as the mammalian material associated with D. laini specimens. However, P. namaquensis appears to be present throughout the Namibian Miocene sequence, associated also with other, older Diamantornis oospecies (Senut et al. 1994), and the family Giraffidae is well known to have representatives extending as far back as the early Miocene (Churcher 1978). Thus, the biostratigraphic precision of the Namibian eggshell fossils, including D. laini, leaves much to be desired. Specimens of D. cf. laini from the Lower Nawata Member of Lothagam are dated to between 7.4 and 6.5 Ma. (Harrison and Msuya 2005). Whereas no eggshell is reported from below these levels, D. cf. laini is replaced above the Marker Tuff by a more derived form of Diamantornis (Harrison and Msuya 2005). Metrically, the Lothagam D. cf. laini is almost indistinguishable from Namibian D. laini and from the Baynunah sample. The Baynunah fauna has been biostratigraphically constrained to somewhere between 8 and 6 Ma (Whybrow and Clements 1999a). The presence of D. laini may suggest that the Baynunah fauna is no younger than 6.5 Ma. In a similar case, the Baynunah hippopotamid Archaeopotamus cf. lothagamensis (Gentry 1999; Boisserie 2005) bears affinities with the Lower Nawata A. lothagamensis, which occurs between 8 and 6.5 Ma (Weston 2000).
The presence of D. laini in Arabia serves to strengthen the biogeographical affinity of this region to Africa, to the exclusion of Eurasia. Specifically, it is of interest that fossil eggshell of the Namibian Diamantornis–Struthio type has been reported from sites in Kenya, Malawi, Namibia, and Tanzania but not from other well-known sequences to the north of the continent such as from the Middle Awash in Ethiopia, the Tugen Hills, Kenya, the deposits of Chad, or from Sahabi, Libya. Boisserie (2005) has found endemic interconnections between the fossil hippopotamids at Lothagam and those from the Baynunah fauna. With the continued lack of reports of fossil eggshell of Diamantornis from Central and North African fossil sites, it is possible to propose a hypothesis of a strengthened faunal relationship between the Baynunah fauna and specifically those of southern East Africa and western southern Africa. Models depicting the different stages of the Red Sea's formation and the proximity of the Arabian and African portions of the Straits of Mendeb allow for a dry route between these two regions across which terrestrial forms could have migrated throughout the Late Miocene, until at least about 6.2 Ma (Redfield et al. 2003).
D. laini is reported from arid and fluvial environments in Namibia, and fluvial sediments representing variations of wooded to open-grassy and dry habitats from Lothagam, and the UAE. As Stidham (2004) noted for Struthio daberasensis from Malawi, the wide range of paleohabitats for these extinct ratite forms is well within what is observed for the modern ostrich, Struthio camelus.
 Diagnosis. A form of fossil ratite eggshell characterized by pores shaped as: 1) linearly aligned long, bent, and forked grooves; 2) shorter groove or comma-shaped pores ('dagger-point'); and 3) tiny, circular to ovoid 'needle-point' or 'sting' pores (Sauer 1972). Under magnification, the long groove pore type typically appears comprised of two ovate pores connected by a superficial groove. Reported eggshell thicknesses range between 1.4 and 3.05 mm (Sauer 1972;
1976;
1978;
Sauer and Sauer 1978) with individual samples displaying more conservative thickness ranges. In gross morphology, aepyornithid-type eggshell is extremely similar to eggshell of Aepyornis, the extinct Elephant Bird of Madagascar. Aepyornithid-type eggshell differs from that of Aepyornis in having generally thinner eggshell [based on measurements of Aepyornis of 2.88–4.30 mm given in
Sauer (1972),
Silyn-Roberts and Sharp (1986), and
Hirsch et al. (1997), and our own observations]. Comparison with eggshell of Mullerornis and Paraphysornis was not possible. Aepyornithid-type eggshell differs from Diamantornis in the absence of the large (>2 mm) circular pore complexes characteristic of Diamantornis. Aepyornithid-type eggshells are on average slightly thinner (compared to D. laini) to significantly thinner (compared to D. wardi) than eggshells of Diamantornis. Aepyornithid-type eggshell differs from those of Struthio in the absence of pore complexes in the aepyornithid-type. It differs from Struthio, Namornis, and Struthiolithus (Psammornis) in the presence of long groove pores in aepyornithid-type eggshells, and in the characteristically linear and sub-parallel arrangement of the pores, as opposed to the more random scattering of pores and pore complexes in Struthio and Namornis. Aepyornithid-type eggshell differs from Incognitoolithus known from the Eocene of North America (Hirsch et al. 1997) by the former's greater eggshell thickness and its stronger resemblance to the Aepyornis pore pattern (dominance of the subparallel long groove pores). Aepyornithid-type eggshells differ from those of Rhea primarily in their significantly greater thickness. It's worth noting that the pore morphologies of Aepyornis, Rhea, and aepyornithid-type eggshell are very similar, at least macroscopically. The similarities between eggshells of Aepyornis, Rhea, Incognitoolithus, and aepyornithid-type fossils that are widely dispersed in time and space suggest that the defining characteristics of these eggshells' morphology may be plesiomorphic for the Ratitae as a whole.
Diagnosis. A form of fossil ratite eggshell characterized by pores shaped as: 1) linearly aligned long, bent, and forked grooves; 2) shorter groove or comma-shaped pores ('dagger-point'); and 3) tiny, circular to ovoid 'needle-point' or 'sting' pores (Sauer 1972). Under magnification, the long groove pore type typically appears comprised of two ovate pores connected by a superficial groove. Reported eggshell thicknesses range between 1.4 and 3.05 mm (Sauer 1972;
1976;
1978;
Sauer and Sauer 1978) with individual samples displaying more conservative thickness ranges. In gross morphology, aepyornithid-type eggshell is extremely similar to eggshell of Aepyornis, the extinct Elephant Bird of Madagascar. Aepyornithid-type eggshell differs from that of Aepyornis in having generally thinner eggshell [based on measurements of Aepyornis of 2.88–4.30 mm given in
Sauer (1972),
Silyn-Roberts and Sharp (1986), and
Hirsch et al. (1997), and our own observations]. Comparison with eggshell of Mullerornis and Paraphysornis was not possible. Aepyornithid-type eggshell differs from Diamantornis in the absence of the large (>2 mm) circular pore complexes characteristic of Diamantornis. Aepyornithid-type eggshells are on average slightly thinner (compared to D. laini) to significantly thinner (compared to D. wardi) than eggshells of Diamantornis. Aepyornithid-type eggshell differs from those of Struthio in the absence of pore complexes in the aepyornithid-type. It differs from Struthio, Namornis, and Struthiolithus (Psammornis) in the presence of long groove pores in aepyornithid-type eggshells, and in the characteristically linear and sub-parallel arrangement of the pores, as opposed to the more random scattering of pores and pore complexes in Struthio and Namornis. Aepyornithid-type eggshell differs from Incognitoolithus known from the Eocene of North America (Hirsch et al. 1997) by the former's greater eggshell thickness and its stronger resemblance to the Aepyornis pore pattern (dominance of the subparallel long groove pores). Aepyornithid-type eggshells differ from those of Rhea primarily in their significantly greater thickness. It's worth noting that the pore morphologies of Aepyornis, Rhea, and aepyornithid-type eggshell are very similar, at least macroscopically. The similarities between eggshells of Aepyornis, Rhea, Incognitoolithus, and aepyornithid-type fossils that are widely dispersed in time and space suggest that the defining characteristics of these eggshells' morphology may be plesiomorphic for the Ratitae as a whole.
Known Distribution. Africa: Mio-Pliocene of Lanzarotte, Canary Islands (Sauer 1972); Mio-Pliocene of Ouarzazate, Morocco (Sauer and Sauer 1978); Early Miocene localities of the Tsondab Sandstone Formation, Namibia (Pickford et al. 1995); Late Miocene locality of Lothagam, Kenya (Harris and Leakey 2003); Asia: Late Miocene and Pliocene of Turkey (Sauer 1976); Late Miocene localities of the Baynunah Formation, United Arab Emirates (this paper); and ?Pliocene Hasnot Locality, Dhok Pathan Horizon, Siwaliks, Punjab, India (Sauer 1972). We also include a ?Pliocene locality east of Bayan Zag (Shabrakh Usu), Mongolia, from which exists a collection that Sauer (1972) felt probably represents the 'Aepyornithoid' type.
New Material. Twenty-five eggshell fragments from the Late Miocene Baynunah Formation, Abu Dhabi Emirate, United Arab Emirates (AUH 924, 925, 927, 932, 935, 936, 938, 939, 945, 994, 1012–1026).
Localities Present: Jebel Barakah (B5–0), Hamra (H3–1 H5–0, H5–2), Shuwaihat (S2–5), Talahafa (TL1–0).
Description. Most of the specimens derive from a single site (TL1–0) and are relatively well preserved, exhibiting only moderate weathering. Maximal fragment length ranges between about 2 and 4.5 cm. The thickness of the 25 eggshells ranges from 1.65 to 2.29 mm with a mean thickness of 1.93 mm (standard deviation 0.16). Pore morphologies are dominated by long groove-pores in sub-parallel arrangement, along with shorter groove pores, and small ovate to round pores. Some specimens exhibit a predominance of small ovate to round pores with areas of eggshell devoid of the long groove pores.
Here we present a new fossil sample for a type of ratite eggshell that had previously been referred to as 'Type A' or 'Aepyornithoid' type (for example, Sauer 1972). We include in our assessment of the aepyornithid-type all forms classified by Sauer and other workers as Aepyornithoid or Type A eggshell, marked particularly by its relative thickness and the presence of subparallel groove pores, and we include eggshell fragments from these same assemblages that display what Sauer termed to be Type A–Type S transitional morphologies (explained below).
Two of us (F.B. and B.P.K.) found and collected eggshell fragments from the Oligocene (Early Oligocene?) locality of Tashgain Bel, Hsanda Gol Formation and the Late Miocene locality of BUK–A, Loh Formation, Mongolia. These fragments appear to be aepyornithid-type, exhibiting an eggshell thickness of about 2.5 mm and variations of groove and circular pore morphologies. In addition, Sauer (1972) described eggshells supposedly derived from the Eocene Irdin Manha Formation, Camp Margetts near Iren Dabasu, Inner Mongolia, China. If the taxonomic attribution and the stratigraphic provenance, respectively, of these two records can be confirmed, the age of the earliest aepyornithid-type eggshell would extend into the Oligocene and Eocene. Nevertheless, Aepyornithid-type eggshell has been well documented from deposits extending from the Early Miocene to the Pliocene and is known to have occurrences from areas as far apart as Central Asia and southwestern Africa. Thus, aepyornithid-type eggshell, known cross-continentally and throughout most of the Neogene, does not currently seem to be of any precise biogeographic or biostratigraphic utility. Further work on the different samples, particularly microstructural comparisons (e.g., Grellet-Tinner 2000; Zelenitsky and Modesto 2003), may help establish a taxonomic and phylogenetic framework against which a biostratigraphy may be constructed. At the moment, it is likely that more than one species of extinct ratite produced the egg type that we and other workers include within the aepyornithid-type. It is a point of great interest that aepyornithid-type eggshell is found with very little variation across such vast temporal and spatial distributions. This is in contrast to the Namibian Namornis–Diamantornis–Struthio eggshell oospecies that appear to have had relatively restricted geographical and temporal spans.
![]()