DISCUSSION
Referral of Fragmentary Specimens
The difficulty in systematizing many of the taxa described above reflects not only the fragmentary nature of the specimens but also the fact that published anatomical data on isolated bones of squamates is meager. Fossil iguanids are known from a few good Tertiary specimens (e.g., Parasauromalus olseni, Aciprion formosum), and morphological cladistic analyses of Iguania have focused on gross skeletal characteristics of extant species. Thus, there are no characters in the maxilla per se that have been argued to diagnose iguanid clades. I have attempted to redress the problem by comparison with disarticulated Recent and fossil specimens. Much work, however, remains to be accomplished.
Known Richness in Squamate Assemblages
The fossil record has yielded squamate assemblages of greatly differing species richness, in part because sample sizes differ. In the Medicine Pole Hills l.f., 17 lizard and 2 amphisbaenian species have been described. Additionally, four snake taxa have been identified on the basis of numerous vertebrae. Thus, when all squamate taxa are included—that is, when squamates are not split into paraphyletic, partially ecomorphic groups like "Lacertilia"—the Medicine Pole Hills l.f. has about 23 species and is one of the most diverse fossil squamate assemblages heretofore described. The second most diverse assemblage is the Ukhaa Tolgod assemblage from the Late Cretaceous of Mongolia, from which 21 taxa have been described. Notably, the Ukhaa Tolgod assemblage completely lacks snakes (Gao and Norell 2000).
Squamate Species Richness across the Eocene/Oligocene Boundary
Sampling and richness. Particularly relevant to this inquiry is the fact that there are no microvertebrate quarries from which squamate assemblages have been described in the lower Oligocene. The one well-studied quarry from the upper Eocene, which produced the Calf Creek l.f. (Holman 1972), was not very diverse. This fact has two effects on the study of richness. First, it greatly increases the number of known specimens for a few late Eocene taxa without greatly increasing the total number of species. Second, it might lead one to conclude that the late Eocene is in fact better studied than the early Oligocene, because microvertebrate quarries often produce much higher diversities of small taxa than surface collections (e.g.,
Silcox and Rose 2001). However, this conclusion is not necessarily warranted. Calf Creek is the most northerly locality of those compiled by
Sullivan and Holman (1996). Negative latitudinal gradients in species richness are a pervasive feature of nearly all higher taxa in the modern world (e.g.,
Pianka 1966;
Rosenzweig 1995), including reptiles (Kiester 1971;
Pianka 1977), and they have also been described from the fossil record (e.g.,
Crane and Lidgard 1989). One can reasonably assume that squamate richness was likewise deployed in the late Eocene and therefore conclude that the Calf Creek l.f. should be the least rich of all sampled faunas. In other words, it may contribute relatively little to total richness while superficially bettering sampling by inflating the number of specimens.
Sampling statistics for the late Eocene and early Oligocene clarify these points. Counting skull elements alone, there are 109 published squamate specimens from the Chadronian and 93 from the Orellan (compiled from
Estes 1983;
Holman 1972;
Sullivan and Holman 1996). In terms of specimen counts, the Chadronian appears to be better sampled. However, when large specimens—especially glyptosaurine anguids—are removed from the samples (they are much easier to pick up in surface collecting), then sampling counts decline to 66 specimens from the Chadronian and 78 from the Orellan. Even so, frequency distributions of the number of specimens per species (glyptosaurines removed) show that in the Chadronian, two (25%) of the species are represented by
≥10 specimens, both of them from the Calf Creek l.f. In the Orellen, two (only 11%) of the species are represented by
≥10 specimens.
 The situation appears to be quite different in Europe, where the major terrestrial faunal break is known as the Grande Coupure (Stehlin 1909).
Rage (1984;
1986) and
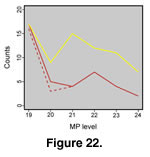
Rage and Augé (1993) have argued that squamate richness in Europe plummets across the Eocene/Oligocene boundary. Richness drops from 16–17 taxa in level MP19 (see
Schmidt-Kittler 1987), to 3–5 in MP20 and 4 in MP21 (Figure 22)
(Rage and Augé 1993). (These estimates do not include boid taxa A–M that Rage included in his 1984 tabulation and would increase the magnitude of the change.) One way to examine sampling issues is by comparing the number of well-correlated localities at each level to species richness. There are 17 such localities for MP19, 9 for MP20, and 15 for MP21 (Figure 22; compiled from
Schmidt-Kittler 1987). It is not necessarily the case that all of these localities have produced described squamate fossils, but the numbers do suggest a decrease in sampling from MP19 to MP20, which may account for the drop in richness at that time. However, richness remains low in MP21 despite many more localities for that level. A drop in richness near the Eocene/Oligocene boundary in Europe is probably robust to increased sampling.
The situation appears to be quite different in Europe, where the major terrestrial faunal break is known as the Grande Coupure (Stehlin 1909).
Rage (1984;
1986) and
Rage and Augé (1993) have argued that squamate richness in Europe plummets across the Eocene/Oligocene boundary. Richness drops from 16–17 taxa in level MP19 (see
Schmidt-Kittler 1987), to 3–5 in MP20 and 4 in MP21 (Figure 22)
(Rage and Augé 1993). (These estimates do not include boid taxa A–M that Rage included in his 1984 tabulation and would increase the magnitude of the change.) One way to examine sampling issues is by comparing the number of well-correlated localities at each level to species richness. There are 17 such localities for MP19, 9 for MP20, and 15 for MP21 (Figure 22; compiled from
Schmidt-Kittler 1987). It is not necessarily the case that all of these localities have produced described squamate fossils, but the numbers do suggest a decrease in sampling from MP19 to MP20, which may account for the drop in richness at that time. However, richness remains low in MP21 despite many more localities for that level. A drop in richness near the Eocene/Oligocene boundary in Europe is probably robust to increased sampling.
North American Record of Species Richness Revisited. With these new data, the current central North American record of squamates is at least qualitatively consonant with the European record and is furthermore consistent with expectation. There are now 25 known Chadronian squamates in comparison with 18 in the Orellan (Table 1). Of the Chadronian taxa, 11 were previously described, and 14 are new. Other Lazarus taxa remain unaccounted for (Table 1). Snake vertebrae from the Medicine Pole Hills l.f. have not yet been described, which will further increase known squamate richness in the Chadronian. However, sampling in the Chadronian is now also better than in the Orellan, so it is difficult to know whether these differences are real.
The apparent central North American drop in richness (~30%) is not as dramatic
as that seen in Europe (>50%;
Rage and Augé 1993). Climatic conditions across the Eocene/Oligocene boundary have not been as well studied in Europe, at least in part because of the discontinuous nature of the fossil record there.
Collinson (1992) noted a generally large decrease in the richness of tropical and subtropical plant species and a sudden increase in the proportion of temperate palynomorphs at the Grande Coupure in the Paris Basin. These floral changes probably reflect cooling, but it is unclear how cooling in Europe compares with that in continental North America.
Grimes et al. (2005) and
Mosbrugger et al. (2005) recently demonstrated that MAT decreased by a few degrees celcius
from the late Eocene to early Oligocene, primarily as a result of increased
MART. Other factors besides temperature are doubtlessly involved in producing
different results, such as the relatively large number of Asian immigrants in
the earliest Oligocene of Europe (Rage and Augé 1993) and the geography of late Eocene Europe.
Poor sampling in North America remains a great impediment to our understanding (see also
Estes 1970;
Hutchison 1992;
Tihen 1964). There is little locality or stratigraphic information for older collections, screenwashing has been sporadic at best, and samples are generally geographically disjunct (geography, independently of time, can now influence the pattern). In this situation techniques like rarefaction that merely address the number of specimens rather than the pattern of their occurrence in space are inapplicable. Thus, an understanding of the response of squamate richness to Eocene/Oligocene climate change will require sampling-standardized collections from stratigraphically controlled localities in geographically restricted areas.
Composition of Late Eocene Squamate Assemblages. The Medicine Pole Hills l.f. is over three times as diverse as the Calf Creek l.f. It is possible that the difference partially reflects the age difference (~1 m.y.) between the localities. However, the difference in local richness is not as great as it appears. Rarefaction is a statistical technique that normalizes richness for sample size (e.g.,
Hurlbert 1971;
Heck et al. 1975). It is appropriate here because we are comparing individual localities, and the results are directly interpretable because the deposits are roughly isotaphonomic. The two amphisbaenian taxa from the Calf Creek l.f. are only known from (a disproportionate number of) vertebrae and constitute outliers in the specimen frequency distribution. Because differing specimen separate rarefaction curves were calculated and were derived for cranial elements alone and for all specimens using the program Analytic Rarefaction 1.3 (Holland 2001) (Figure 23).
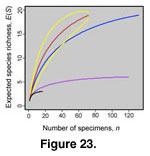 If the specimen sample size from the Medicine Pole Hills l.f. was the same as that from Calf Creek, the richness of the former would be reduced to 10.1 ± 2.8 species, for cranial specimens, versus 18.6 species when all specimens are considered; only three species are known from cranial remains at Calf Creek. The cranial comparison, because it does not rely on a frequency distribution with extreme outliers, is probably the more meaningful of the two analyses. Rarefaction reduces the apparent difference in species richness for cranial remains alone—16 species (19–3)—by more than half, to 7.1 species (10.1–3). The results are still consistent with a latitudinal gradient in local species richness.
If the specimen sample size from the Medicine Pole Hills l.f. was the same as that from Calf Creek, the richness of the former would be reduced to 10.1 ± 2.8 species, for cranial specimens, versus 18.6 species when all specimens are considered; only three species are known from cranial remains at Calf Creek. The cranial comparison, because it does not rely on a frequency distribution with extreme outliers, is probably the more meaningful of the two analyses. Rarefaction reduces the apparent difference in species richness for cranial remains alone—16 species (19–3)—by more than half, to 7.1 species (10.1–3). The results are still consistent with a latitudinal gradient in local species richness.
Depending on the specific status of the taxa present, the Calf Creek and Medicine Pole Hills local faunas have up to five taxa in common, viz., a small and a large rhineurid, Peltosaurus, "Palaeoxantusia" borealis, and Cypressaurus. Calf Creek is conspicuous in lacking a xenosaurid, an acrodontan, a varanid, and a scincoid. (The latter four are represented by few specimens in the Medicine Pole Hills l.f., so their absence at Calf Creek is not altogether surprising.) Additionally, the largest anguid (Helodermoides) is seemingly absent in the Calf Creek l.f.
Gilmore (1928:120) noted the presence of "Glyptosaurus sp." in the "Cypress Hills";
Holman (1976) also listed this taxon without providing information on new specimens. Since the Cypress Hills Formation spans both the Duchesnean and Chadronian land-mammal "ages," and Gilmore provided no locality data for the specimen (a single cephalic osteoderm), the definitive presence of "Glyptosaurus" or Helodermoides cannot be supported with present data. Finally, the Calf Creek l.f. lacks the great diversity of iguanids seen in North Dakota.
The Medicine Pole Hills squamate assemblage is in some respects intermediate between early Eocene and Miocene faunas. There are two holdovers of early Eocene immigrants, the presumed relatives of the varanid Saniwa and the acrodontan Tinosaurus. These taxa are thought to have entered North America during the wet, warm climate of the early Eocene (Estes 1983); they evidently persisted until near the end of the Eocene. Additionally, there are a number of more ancient North American lineages that remain, including a xantusiid and (probably) a xenosaurid. The former apparently disappears in central North America by the beginning of the Oligocene. Despite some similarities to early Eocene squamate assemblages, the Medicine Pole Hills l.f. is remarkable in its diversity of iguanids, in this respect more like the Miocene (Tihen 1964). Over 40% of the lizards are iguanids, as compared with an estimated 20% in the early Eocene Wasatch Formation of Wyoming (Gauthier 1982).
The transformation of central North American squamate assemblages over time has been described as showing a growing prominence of xeric-adapted lizards (especially iguanids).
Tihen (1964) argued that modernization began in the late Oligocene and was centered in the early Miocene (Arikareean). Regardless of the poverty of sampling in the medial and late Oligocene, the data presented here superficially contradict Tihen's thesis in the high proportion of iguanids present in the Medicine Pole Hills l.f., but only if we assume that by modernization is meant "phrynosomatization." In other words, there is contradiction only if we are wed to the idea that late Eocene iguanids are of the type now inhabiting the western interior (i.e., phrynosomatine and crotaphytine). Of those taxa that can possibly be related to extant clades, one is a phrynosomatine (this study) and one a crotaphytine (Holman 1972;
Estes 1983;
McGuire 1996), but one is a polychrotine. Thus, at least one iguanid clade may show the same pattern as many scincomorph and anguimorph lizards, in which Eocene taxa retreated or became restricted to the south to become members of the modern Sierra-Madrean and Austroriparian herpetofaunas. An increased proportion of iguanids does not by itself imply that the Medicine Pole Hills squamate fauna is any meaningful sense "modern." That Iguanidae may show such a historical biogeographical pattern should not be surprising in view of the great variability in autecology within the clade.
