
First report of Leptonectes (Ichthyosauria: Leptonectidae) from the Lower Jurassic (Pliensbachian) of Asturias, northern Spain
Article number: 21.2.29A
https://doi.org/10.26879/802
Copyright Paleontological Society, August 2018
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 7 July 2017. Acceptance: 2 July 2018
{flike id=2275}
ABSTRACT
Early Jurassic ichthyosaurs are well-known based on the abundant and exquisitely preserved European records of the Western Tethys. For example, faunas from Southern England and the Toarcian Posidonia shales of Holzmaden and surrounding areas (Germany) are known worldwide. However, between these areas there are still gaps and/or under-sampled localities from the northern coast of Europe. In recent years as a result of systematic palaeontological and geological explorations of Pliensbachian fossiliferous localities along the coast of Asturias (northern Spain), ichthyosaur specimens have been collected. One of them can be identified at lower taxonomic levels and it is referred to Leptonectes sp. These new findings highlight the richness of the fossil record of the northern coast of Spain and suggest that the abundance of Pliensbachian ichthyosaurs worldwide may have been underestimated.
Marta S. Fernández. División Paleontología Vertebrados, Museo de La Plata, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, CONICET, 1900 La Plata, Argentina. martafer@fcnym.unlp.edu.ar
Laura Piñuela. Museo del Jurásico de Asturias (Jurassic Museum of Asturias), Colunga 33328, Spain. lpinuela.muja@gmail.com
José Carlos García-Ramos. Museo del Jurásico de Asturias (Jurassic Museum of Asturias), Colunga 33328, Spain. jcgramos.muja@gmail.com
Keywords: ichthyosaurs; Pliensbachian; Lower Jurassic; Asturias; Spain
Final citation: Fernández, Marta S., Piñuela, Laura, and García-Ramos, José Carlos. 2018. First report of Leptonectes (Ichthyosauria: Leptonectidae) from the Lower Jurassic (Pliensbachian) of Asturias, northern Spain. Palaeontologia Electronica 21.2.29A 1-15. https://doi.org/10.26879/802 palaeo-electronica.org/content/2018/2275-spanish-ichthyosaurs
Copyright: August 2018 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
Ichthyopterygians were successful marine reptiles during most of the Mesozoic. Their fossil record spans from the Olenekian (Early Triassic) up to the Cenomanian (middle Late Cretaceous) (Fischer et al., 2014). Throughout this long history, Early Jurassic forms are well known based on the abundant and exquisitely preserved European records of the Western Tethys. However, there are still some gaps in the record and/or under-sampled geographic areas. Thus, Pliensbachian ichthyosaurs are comparatively scarce and/or not as well known as Hettangian-Sinemurian or Toarcian forms. However, the abundance of Pliensbachian occurrences will probably increase significantly as a new late Pliensbachian-early Toarcian lagerstätte, including ichthyosaurs, has been reported in Alberta (Canada) (Martindale et al., 2017).
Presently, Pliensbachian taxa are represented by a few species referred to Temnodontosaurus nuertengensis (Huene, 1931) species inquirenda sensu McGowan and Motani (2003), from the early Pliensbachian of Southern Germany; Ichthyosaurus anningae Lomax and Massare, 2015; Ichthyosaurus conybeari Lydekker, 1888 (Massare and Lomax, 2016a); Leptonectes moorei McGowan and Milner, 1999 from the Lower Pliensbachian of Dorset (England); and Leptonectes tenuirostris (Conybeare, 1822). This latter species is commonly found in the Blue Lias of Lyme Regis, however, its stratigraphic range was extended up to the Pliensbachian based on a fragment of a forefin from Belgium (Godefroit, 1992) and a skull and some postcranial remains from Switzerland (Maisch and Reisdorf, 2006). Also, a fragment of a rostrum from the Lower Pliensbachian of Bielefeld (Germany) has been referred as Temnodontosaurus sp. (Hungerbühler and Sachs, 1996).
Several Early Jurassic fossiliferous localities, particularly those of Pliensbachian age, are well known along the Atlantic coasts of central region of Portugal and Spain (centro-oriental Asturian coast) and marine reptiles have been mentioned in the literature as early as the nineteenth century (Pereda Suberbiola et al., 2010). However, marine reptile records are still comparatively scarce and poorly known not only from Spain (Ruiz-Omeñaca et al., 2006), but also from Portugal (Zbyszeski and Moitinho de Almeida, 1952; Ferreira Da Veiga, 1958; Crespo, 2002). Among Jurassic marine reptiles from the Iberian Peninsula, ichthyosaurs are the poorest represented, and they were tentatively identified as Ichthyosaurus sp. and Stenopterygius sp. Bardet et al. (2008b). Among the Portuguese material there is a complete and articulated forefin (Zbyszewski and Moitinho de Almeida, 1952, plate II, figure 4).
During the last decade, palaeontological and geological explorations of the Asturian coastal cliffs, carried out by scientific staff of the Museo del Jurásico de Asturias (MUJA), resulted in the recovery of several ichthyosaur remains from the Rodiles Formation (Pliensbachian, Early Jurassic). These materials are held at the MUJA, where they were mechanically prepared. The new findings, plus isolated remains displayed to local people at this institution, document an almost unknown Pliensbachian fauna from Spain. These findings are significant as they represent the most complete ichthyosaurs from Spain and one of the few from the Iberian Peninsula.
GEOLOGICAL CONTEXT
The most spectacular outcrops of the Jurassic successions in Asturias (northern Spain) extend, almost continuously, along the coast between the Gijón and Ribadesella localities (Figure 1). This section of littoral is known as “The Dinosaur Coast” because of the abundance of bones and footprints of these reptiles (García-Ramos et al., 2006).
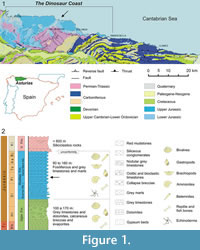 The Jurassic series are grouped into two high-order lithostratigraphic units (Valenzuela et al., 1986). The lower unit (Villaviciosa Group) is dominated by carbonate rocks of littoral (Gijón Formation) and open marine (Rodiles Formation) origin. The upper unit (Ribadesella Group) consists of siliciclastic rocks of continental origin (alluvial fan and fluvial) included within the La Ñora and Vega formations, respectively, and shelf lagoon and fluvial-dominated deltas, represented by the Tereñes and Lastres formations, respectively. The boundary between Rodiles and Vega formations is a regional unconformity including a stratigraphic gap of variable range related to an extensional phase which took place during the Late Jurassic-Early Cretaceous.
The Jurassic series are grouped into two high-order lithostratigraphic units (Valenzuela et al., 1986). The lower unit (Villaviciosa Group) is dominated by carbonate rocks of littoral (Gijón Formation) and open marine (Rodiles Formation) origin. The upper unit (Ribadesella Group) consists of siliciclastic rocks of continental origin (alluvial fan and fluvial) included within the La Ñora and Vega formations, respectively, and shelf lagoon and fluvial-dominated deltas, represented by the Tereñes and Lastres formations, respectively. The boundary between Rodiles and Vega formations is a regional unconformity including a stratigraphic gap of variable range related to an extensional phase which took place during the Late Jurassic-Early Cretaceous.
The marine reptile specimens of Asturias are recovered from the Rodiles Formation, which is Late Sinemurian-Early Bajocian in age (Suárez Vega, 1974; Gómez and Goy, 2000; Comas-Rengifo and Goy, 2010), and is dominated by limestone-marl alternations (90-170 m thick). This formation has been subdivided into the Buerres and Santa Mera members. The Buerres Member, reaching a maximum thickness of 34 m is Late Sinemurian (Obtusum Zone) in age, and characterised by nodular limestones with some thin marl layers, representing the more proximal inner part of a carbonate ramp (Valenzuela et al.,1985, 1986). The Santa Mera Member, up to 168 m thick, is Late Sinemurian (Oxynotum Zone) to Early Bajocian in age, and consists predominantly of tabular layers of limestones and marls adopting a rhythmic character that represents the middle and outer part of the ramp (Valenzuela et al., 1985, 1986; García-Ramos and Piñuela, 2010).
The present study focuses on the ichthyosaur skeletons found in the Santa Mera Member, specifically in the Jamesoni Zone (Pliensbachian) (Comas-Rengifo and Goy, 2010), composed of a rhythmic alternation of limestones, dark grey marls, calcareous mudstones and black shales that also contain abundant invertebrate macrofossils such as ammonites, bivalves, belemnites, brachiopods and echinoderms, as well as diverse trace fossils including: Asterosoma, Chondrites, Diplocraterion, Kulindrichnus, Phymatoderma, Planolites, Rhizocorallium, Thalassinoides and others (Suárez Vega,1974; Valenzuela et al., 1986, 1989; García-Ramos et al., 2011).
SYSTEMATIC PALAEONTOLOGY
Order ICHTHYOSAURIA de Blainville, 1835
Unranked PARVIPELVIA Motani, 1999
Family LEPTONECTIDAE Maisch, 1998
Genus LEPTONECTES McGowan, 1996
Leptonectes sp.
Figure 2, Figure 3, Figure 4, Figure 5, Table 1
Referred specimen. MUJA 3687, incomplete skeleton consisting of skull and jaw elements, vertebrae including atlas-axis and associated ribs, partially articulated proximal portion of the forefin and both coracoids.
 Horizon and locality. Ensenada de La Conejera (43º 32´ 6.18´´N; 5º 22´3.02´´ W), Selorio, Villaviciosa, Asturias, Spain. Rodiles Formation, Jamesoni Zone (Lower Jurassic, Pliensbachian). Ensenada de La Conejera (Selorio, Villaviciosa), Asturias, northern Spain.
Horizon and locality. Ensenada de La Conejera (43º 32´ 6.18´´N; 5º 22´3.02´´ W), Selorio, Villaviciosa, Asturias, Spain. Rodiles Formation, Jamesoni Zone (Lower Jurassic, Pliensbachian). Ensenada de La Conejera (Selorio, Villaviciosa), Asturias, northern Spain.
Description. The MUJA 3687 has been found in an area of approximately 4 m2. During extraction it was separated in several blocks to be transported. The skeleton is partially disarticulated, and bones are distributed in three main blocks (Figure 2). Block 1 contains elements of the posterior portion of a mandible, a quadrate, a partial jugal and part of the pterygoid, and numerous teeth. Block 2 comprises most of the vertebral column and ribs, pectoral girdle and forefin elements including the humerus. Block 3 bears caudal vertebrae including the posteriormost postflexural centra. The final burial has been followed by extensive pyritization, and most of the bones are strongly pyritized. This fact, and the disposition of the ribs overlying most of the skeleton, obscures the identification of most mandible and skull bones. General measurements of the MUJA 3687 are given on Table 1.
 Skull and mandible: The skull and mandibles are poorly preserved. Most elements of the mandible are present but cannot be precisely identified. Identifications are shown in Figure 3. As in all leptonectids, the jaw is slender and relatively long. An approximately 33 cm long fragment with a shallow groove that runs along the element is identified as part of a dentary (Figure 3.2). Scattered around this bone are several teeth. There is a single, almost complete quadrate. The stout articular portion of the bone, bearing the articular condyle, is separated by a short neck from the pterygoid and occipital lamellae. This lamella is broad with a conspicuous anterior-posterior expansion (Figure 4). Close to the quadrate, the posterior portion of a jugal, tentatively identified as the right one, is exposed. Its dorsal ramus is well-developed and strongly curved dorsally. The suborbitalbar of the jugal is robust and rod-like. The height of the preserved suborbitalbar is 18 mm. Close to the jugal are two sclerotic plates.
Skull and mandible: The skull and mandibles are poorly preserved. Most elements of the mandible are present but cannot be precisely identified. Identifications are shown in Figure 3. As in all leptonectids, the jaw is slender and relatively long. An approximately 33 cm long fragment with a shallow groove that runs along the element is identified as part of a dentary (Figure 3.2). Scattered around this bone are several teeth. There is a single, almost complete quadrate. The stout articular portion of the bone, bearing the articular condyle, is separated by a short neck from the pterygoid and occipital lamellae. This lamella is broad with a conspicuous anterior-posterior expansion (Figure 4). Close to the quadrate, the posterior portion of a jugal, tentatively identified as the right one, is exposed. Its dorsal ramus is well-developed and strongly curved dorsally. The suborbitalbar of the jugal is robust and rod-like. The height of the preserved suborbitalbar is 18 mm. Close to the jugal are two sclerotic plates.
 Dentition: The teeth are long, slender, straight and/or only slightly curved. The mean length of teeth is approximately 3.2 cm, and the tooth crown width/length ratio is 0.2. The enamel is almost smooth with very subtle striations, and the base of the enamel layer is poorly defined. In those teeth with crown and root completely preserved, crowns are relatively long (tooth length/crown length ratio = 2.0-2.5), and roots are straight without evident grooves (Figure 3.1).
Dentition: The teeth are long, slender, straight and/or only slightly curved. The mean length of teeth is approximately 3.2 cm, and the tooth crown width/length ratio is 0.2. The enamel is almost smooth with very subtle striations, and the base of the enamel layer is poorly defined. In those teeth with crown and root completely preserved, crowns are relatively long (tooth length/crown length ratio = 2.0-2.5), and roots are straight without evident grooves (Figure 3.1).
Axial Skeleton: Specimen MUJA 3687 preserves several vertebrae corresponding to different section of the vertebral column including the atlas-axis complex, cervicals, dorsals, and pre- and post-flexural caudals. Except for two short sections (of approximately five to six caudal centra), the vertebrae are disarticulated, and most are exposed in anterior or posterior views, making it difficult to determine their exact location within the column. The atlas and axis are fused with suture between them visible in lateral view. The atlas-axis complex is almost complete and exposed in ventrolateral view showing a prominent concave and ovoid facet for the axial intercentrum (Figure 5.1). In ventral view, a triangular facet for the reception of the atlantal intercentrum is also clearly defined. In lateral view, both atlas and axis centra are very short in comparison with the remaining cervical centra, and the axis notably shorter than the atlas (L=18 mm and 24 mm, respectively). Diapophyses and parapophyses of the axis are better demarcated than those apophyses of the atlas. The edge of the posterior articular surface of the axis is surrounded by a well-defined crest. In the same slab, and close to the atlas-axis, there is a cervical centrum, probably corresponding to the third cervical, which is 51.2 mm wide, 21.3 mm long and 47.3 mm high. The remaining cervical and/or dorsal centra are also strongly amphicoelous, none of them are articulated with the neural spines, and are similar in proportion being wider than high. There are few neural arches scattered among the cervical centra with pre- and postzygapophyses separated, so they could correspond to cervical and/or anterior dorsal vertebrae. Six pre-flexural caudal centra are partially articulated. Many ribs are preserved without a predominant orientation. The best preserved are bicipitals and depict a shallow groove along their proximal portion.
Pectoral girdle and forefin: The partial pectoral girdle (Figure 5.3) comprises the articulated coracoids. One of them is complete, but partially overlapped by vertebral centra and rib fragments, and it is interpreted as the left coracoid exposed in ventral view. It is massive and slightly anteroposteriorly longer than mediolaterally wide (length = 134 mm, width = 121 mm). On the lateral edge there is a short and poorly demarcated facet for the articulation with the scapula. The glenoid contribution of the coracoid is approximately twice the length of the facet for the scapula, and subequal to the intercoracoid facet. The posterior edge is slightly damaged, however, and it appears to not bear a posterior notch.
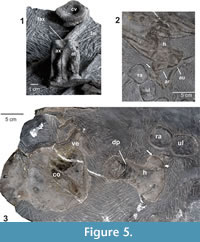 One humerus, interpreted here as the left, is preserved in three dimensions. Although it is still included in the matrix, its dorsal and ventral surfaces have been prepared and provide enough anatomical information (Figure 5.2-3). The humerus is characterized by a strong anteroposterior expansion of its distal end with an anterior facet on its leading edge. The maximum length of the humerus is 133 mm and its maximum distal width is 108 mm. The head of the humerus and the deltopectoral crest are covered with roughened bone, suggesting the persistence of an extensive cartilaginous cup covering this area. The humerus surface, here identified as the dorsal one (Figure 5.2), is partially hidden by four ribs; however, a relatively well-developed dorsal process can be distinguished. In ventral view (Figure 5.3) there is also a relatively well-developed deltopectoral crest. The distal extension of the deltopectoral crest on the ventral side of the humerus shaft resembles that of Wahlisaurus massarae (Lomax, 2016, figure 6). The shaft is constricted and stout. Both the leading and posterior edges of the bone are curved, but the leading edge is strongly concave. Distally the humerus is expanded and its leading edge bears an anterior facet. In dorsal and ventral views, the facets for the radius and ulna are relatively straight. The ulnar facet is longer than the radial one.
One humerus, interpreted here as the left, is preserved in three dimensions. Although it is still included in the matrix, its dorsal and ventral surfaces have been prepared and provide enough anatomical information (Figure 5.2-3). The humerus is characterized by a strong anteroposterior expansion of its distal end with an anterior facet on its leading edge. The maximum length of the humerus is 133 mm and its maximum distal width is 108 mm. The head of the humerus and the deltopectoral crest are covered with roughened bone, suggesting the persistence of an extensive cartilaginous cup covering this area. The humerus surface, here identified as the dorsal one (Figure 5.2), is partially hidden by four ribs; however, a relatively well-developed dorsal process can be distinguished. In ventral view (Figure 5.3) there is also a relatively well-developed deltopectoral crest. The distal extension of the deltopectoral crest on the ventral side of the humerus shaft resembles that of Wahlisaurus massarae (Lomax, 2016, figure 6). The shaft is constricted and stout. Both the leading and posterior edges of the bone are curved, but the leading edge is strongly concave. Distally the humerus is expanded and its leading edge bears an anterior facet. In dorsal and ventral views, the facets for the radius and ulna are relatively straight. The ulnar facet is longer than the radial one.
Distal to the humerus, rotated anteriorly 80º from their anatomical location, are two articulated forefin elements. The length of their proximal articular facets is approximately equal to the length of the radial and ulnar facets of the humerus. These elements are interpreted as the radius and ulna. The one located more anteriorly, interpreted as the radius, is roughly as long as wide (length = 47.3 mm; width = 55 mm) and, as in Leptonectes solei and L. moorei, is not notched. Contrary to the condition present in most of the specimens of L. tenuirostris (McGowan, 1993), the foramen between the occlusal edges of the radius and ulna is absent. The ulna is wider than long (antero-posterior width = 62 mm; proximo-distal length = 48 mm), and the proximal and distal articular facets are parallel and almost straight. On its anterior edge it bears an almost straight facet for the articulation with the radius, and its posterior edge is slightly convex.
Comments and remarks. Johnson (1977) proposed several size-independent criteria for estimating relative age in ichthyosaurs including humerus details. Thus, a smooth texture of the cortical bone of the shaft, and well-ossified epiphyses of the humerus (morphologically expressed as a caput humerus convex), indicates an adult state. In MUJA 3687 the texture of the shaft is smooth, and the epiphyses of the humerus, as well as the texture of the external bone layer of the radius and ulna, suggest that the specimen was osteologically mature. Unfortunately, the lack of a cranium prevents testing other auxiliary criteria such as the degree of fusion on the dorsal skull roof or the relative size of the sclerotic ring (Fernández et al., 2005; Maxwell et al., 2012).
The Leptonectidae (Maisch, 1998) is characterized by the relatively long snout, with a variable degree of overbite, and delicate, straight or slightly curved, teeth with smooth enamel. Although incomplete and not well exposed, bones of the mandible (particularly the dentary) of MUJA 3687 suggest a slender and relatively long mandible and so, a relatively long snout. The tooth morphology, as well as coracoids and humerus features, clearly differ from those of the leptonectid genus Wahlisaurus. On the other hand, MUJA 3687 can be confidently referred to Leptonectes based on a combination of forefin features including the humerus with a constricted shaft, the wide distal expansion and the presence of a prominent anterior facet on its leading edge (Godefroit, 1992; McGowan, 1996; McGowan and Motani, 2003).
Within the genus Leptonectes, besides the type species L. tenuirostris Conybeare, 1822, there are two other valid species: L. solei McGowan, 1993 and L. moorei McGowan and Milner, 1999 (McGowan and Motani, 2003; Lomax, 2016). Leptonectes tenuirostris is well known by several specimens (McGowan, 1989), and it has the longest stratigraphic range (probably from the Rhaetian up to the late Pliensbachian, Maisch and Reisdorf, 2006), and the widest geographic occurrence among leptonectids, including records from England (McGowan, 1996), Belgium (Godefroit, 1992), Germany (Maisch, 1999) and Switzerland (Maisch and Reisdorf, 2006). In contrast, L. solei and L. moorei are geographically restricted to Dorset (England), and stratigraphically to the Sinemurian (Obtusum Zone) (McGowan, 1993), and early Pliensbachian (McGowan and Milner, 1999), respectively.
Well-ossified epiphyses, and the smooth texture of the humerus shaft suggest that MUJA 3687 was an adult or a subadult; and the length of preserved elements indicates that this specimen is within the size range of Leptonectes tenuirostris (McGowan,1993), considerably smaller than the holotype of Leptonectes solei and slightly larger than the holotype and only known specimen of L. moorei.
Leptonectidae indet.
Figure 6.1-6.2
Referred specimen. MUJA 4095. Partial left dentary, anterior tip of the snout including both premaxillae and the anterior most portion of the right dentary with teeth. In the same block of the left dentary there are several scattered teeth.
 Horizon and locality. Rodiles Formation, Jamesoni Zone (Lower Jurassic, Pliensbachian). Ensenada de La Conejera (Selorio, Villaviciosa), Asturias, northern Spain.
Horizon and locality. Rodiles Formation, Jamesoni Zone (Lower Jurassic, Pliensbachian). Ensenada de La Conejera (Selorio, Villaviciosa), Asturias, northern Spain.
Description. MUJA 4095 preserves a fragment of the right dentary exposed in alveolar view. Most of the bone has been preserved as an impression and/or embedded on the matrix. The total preserved length of the dentary is 220 mm. The alveolar groove is deep and two tooth roots remain in their anatomical positions. The dorsal edges of the labial and lingual surfaces of the grooves are slightly waved, marking the locations of the alveoli. The impression of dentary depicts a slender lower jaw. Close to the dentary are two fin elements. One is roughly rectangular and the other is polygonal. Both are very small (proximal-distal length 8.9 mm and 9.9 mm, respectively), and the roughness of their texture suggest poor ossification. The anterior most ends of both premaxillae are preserved articulated and only slightly displaced from their anatomical location. There is a shallow groove on the lateral wall of the left premaxilla. Close to the ventral border of the left premaxilla, the anterior portion of the right dentary is exposed in alveolar view. Scattered in the same block, and close to the dentary and tip of the mandible, there are numerous teeth. Most of them are completely exposed and, on several crowns, the enamel can be clearly discerned. The height of the largest tooth is 17.6 mm (total height) and 10 mm (crown height). All teeth are almost straight, slender, with pointed apices. The crowns are elongated and the enamel layer is thin without conspicuous ornamentations; there are only very subtle ridges apicobasally oriented. The roots depict no evident grooves.
Comments and remarks. The texture of the preserved bones suggests that MUJA 4095 was osteologically immature, and its size is compatible with that expected in a perinatal individual.
The taxonomic value of tooth morphology (size and shape) of Early Jurassic ichthyosaurs has been questioned. Recent contributions have shown that size and shape of teeth change during ontogeny, at least in Stenopterygius (Dick and Maxwell, 2015; Dick et al., 2016). In the case of Ichthyosaurus communis and I. intermedius, historically differentiated on the basis of tooth morphology, Massare and Lomax (2017) recognized different morphologies, although they pointed out that tooth features were not reliable to distinguish these two species. Nevertheless, the dentition pattern of MUJA 4095 is consistent with that of Leptonectidae, particularly with MUJA 3687 and differs from that of Wahlisaurus massarae characterized by recurved crowns and large, bulbous infolded roots (Lomax, 2016). On the other hand, teeth of MUJA 4095 are inconsistent with tooth morphology of the other Lower Jurassic ichthyosaurs such as Temnodontosaurus, Ichthyosaurus and Stenopterygius, even considering ontogenetic change (Dick and Maxwell, 2015).
Ichthyosauria indet.
Figure 6.3-6
Referred specimens. MUJA 1020 (partial mandible symphysis and teeth), 4154 (fin elements), 4069 (four articulated vertebrae), 4227 (rostral fragment with teeth).
Horizon and locality. Rodiles Formation, Jamesoni Zone (Lower Jurassic, Pliensbachian). Santa Mera (Selorio, Villaviciosa) (MUJA 1020, 4154, and 4227) and Huerres (San Juan de Duz, Colunga) (MUJA 4069), Asturias, northern Spain.
Description. MUJA 1020 (Figure 6.3) consists of both mandibular rami articulated through the symphysis, however, the left ramus is slightly dislocated in such a way that the lateral wall of the left dentary is exposed and hides much of the left splenial. The preserved portion of the mandible is 190 mm long. The roots have not been completely preserved, but in two teeth the crowns seem to be relatively long. Both teeth end in pointed apices and the enamel layer lacks conspicuous ornamentations. The enamel is best preserved in the largest tooth and its base is not well defined.
MUJA 4154 (Figure 6.4) comprises three small phalanges. The anteroposterior length of the best exposed is 30 mm, and its proximodistal length is 23 mm. Most of its articular facets are straight. The polygonal outline of the phalanges suggests that they are not the most distal phalanges and their size indicates that they correspond to a juvenile specimen.
MUJA 4069 (Figure 6.5) comprises four vertebral centra that are partially articulated. An isolated neural arch, in ventral view, is associated with the vertebrae, and overlaps one of them. The anteroposterior length ranges from 24.5 mm to 17.7 mm, with a height/length ratio of 2. Unfortunately, since the lateral surfaces of the centra are not well preserved enough to distinguish the presence and/or location of the apophyses, it is difficult to establish their location in the vertebral series.
MUJA 4227 (Figure 6.6) consists of a posterior fragment of a premaxilla bearing a deep alveolar groove, overlapped by a fragment of the maxilla and a tooth. The tooth crown is relatively long and straight, and the base of the enamel is well defined. Aligned striation runs along most of the crown but not reaching the apical zone. The tooth morphology of this specimen clearly differs from those of the MUJA 3687, 4095 and 1020, suggesting that there are at least two different species in the same area.
DISCUSSION
In recent years, knowledge of Lower Jurassic ichthyosaurs has increased significantly. This progress is mainly triggered by re-examinations of British collections (e.g., Lomax and Massare, 2012, 2015; Lomax and Gibson, 2015; Martin et al., 2012; Massare and Lomax, 2016a, 2016b, 2017), and demonstrated that alpha taxonomy of iconic species represented by complete and, in most of the cases, well-preserved, specimens were in need of revision. It also highlights the need of increasing character sampling, but also the importance of systematic revisions of historical collections. The key practice of alpha taxonomy is the first, and usually underestimated, step upon which hypotheses of higher levels (from phylogeny up to macroevolutionary scenarios) rest. Contributions, as the mentioned above, showed that Early Jurassic diversity was underestimated, and that Leptonectes diagnosis, and species delimitations, are not very clear.
Northern Spain Ichthyosaurs
Although Lower Jurassic outcrops, particularly Pliensbachian ones, in the Iberian Peninsula (Spain and Portugal) are abundant and marine reptiles have been recovered from them as early as the nineteenth century (Schulz, 1858, Sauvage, 1897-1898; Zbyszeski and Moitinho de Almeida, 1952; Ferreira da Veiga, 1958; Crespo, 2002; Pereda Suberbiola et al., 2010), Iberian marine faunas are still poorly known (Bardet et al., 2008b). In the case of ichthyosaurs, fossils described herein represent, at least to our knowledge, the first ichthyosaurs to be described in detail and figured from the Jurassic of Spain, and one of the few from the Iberian Peninsula. Also, the tooth morphology of one of the specimen (MUJA 4227) clearly differs from the other specimens suggesting that at least two different species lived in the same area.
Most of the marine vertebrate remains recovered from the Pliensbachian of northern Spain cliffs correspond to ichthyosaurs, indicating that these reptiles were a major component of Iberian faunas during this age. To the six specimens, recovered with a strict stratigraphic control by the scientific staff of the MUJA, an additional four specimens were collected from the same area by local people (Table 2). Interestingly, in this Pliensbachian Asturian assemblage, three of the six specimens collected by the MUJA, are consistent in their size with juvenile individuals. Apart from ichthyosaurs, the only known marine reptile from the Rodiles Formation is a juvenile plesiosaur (Bardet et al., 2008a). Whether the coasts of northern Spain could act as a protected environment used for reproduction by these viviparous pelagic marine reptiles (Hogler, 1992; Deeming et al., 1993; Stinnesbeck et al., 2014) requires further scrutiny, but as a working hypothesis must not be dismissed. These findings highlight the fossiliferous potential of the localities of northern Spain, and that the relative low abundance of Pliensbachian ichthyosaurs worldwide may be due to incomplete and/or uneven sampling of Lower Jurassic localities. New discoveries as those reported here, as well as the re-examination of historical collections (e.g., Massare and Lomax, 2016b), and from other Lagerstätten such as the one recently described in detail from the Toarcian of Somerset, UK (Williams et al., 2015), will help to provide accurate estimations of palaeodiversity and will also allow to test evolutionary scenarios built on the classical European Lagerstättten.
ACKNOWLEDGEMENTS
The authors would like to thank D. Lomax and two anonymous reviewers whose comments and suggestions have greatly improved this article; and O. Fernández and B. Santaeugenia for the materials bestowed to the MUJA. This work is a contribution to the grants PICT 1039, UNLP N749-853 (MSF), MINECO/FEDER project CGL2015-66835-P (LP) and was funded by Sociedad Pública de Gestión y Promoción Turística del Principado de Asturias (LP and JCG-R) and Ayuntamiento de Villaviciosa (Asturias).
REFERENCES
Bardet, N., Fernández, M., García-Ramos, J.C., Pereda Suberbiola, X., Piñuela, L., Ruiz-Omeñaca, J.I., and Vincent, P. 2008a. A juvenile plesiosaur from the Pliensbachian (Lower Jurassic) of Asturias, Spain. Journal of Vertebrate Paleontology, 28:258-263.https://doi.org/10.1671/0272-4634(2008)28[258:AJPFTP]2.0.CO;2
Bardet, N., Pereda-Suberbiola, X., and Ruiz-Omeñaca, J.I. 2008b. Mesozoic marine reptiles from the Iberian Peninsula. Geo-Temas, 10:1245-1248.
Comas-Rengifo, M.J. and Goy, A. 2010.Caracterización biocronoestratigráfica del Sinemuriense Superior y el Pliensbachiense entre los afloramientos de la Playa Vegay Lastres (Asturias), p. 10-18. In García-Ramos, J.C. (ed.), Las sucesiones margo-calcáreas marinas del Jurásico Inferior y las series fluviales del Jurásico Superior. Acantilados de la playa de Vega (Ribadesella). Guía de la excursión A del V Congreso del Jurásico de España. Museo del Jurásico de Asturias, Colunga, Spain.
Conybeare, W.D. 1822. Additional notices on the fossil genera Ichthyosaurus and Plesiosaurus. Transactions of the Geological Society of London, 1:103-123. https://doi.org/10.1144/transgslb.1.1.103
Crespo, E.G. 2002. Paleoherpetofauna portuguesa. Revista Española de Herpetología, volumen especial:17-35.
de Blainville, H.D. 1835. Description de quelques espèces de reptile de la Californie, précédée del’analysed’un système general d’Erpétologie et d’Amphibiologie. Nouvelles Annales du Muséum d’Histoire Naturelle, Paris, 4:233-295.
Deeming,D.C., Halstead, L.B., Manabe, M., and Unwin, D.M. 1993. An ichthyosaur embryo from the Lower Lias (Jurassic: Hettangian) of Somerset, England, with comments on the reproductive biology of ichthyosaurs. Modern Geology, 18:423-442.
Dick, D.G. and Maxwell, E.E. 2015. Ontogenetic tooth reduction in Stenopterygius quadriscissus (Reptilia: Ichthyosauria): Negative allometry, changes in growth rate, and early senescence of the dental lamina. PLoS ONE, 10:e0141904. https://doi.org/10.1371/journal.pone.0141904
Dick, D.G., Schweigert, G., and Maxwell, E.E. 2016. Trophic niche ontogeny and palaeoecology of early Toarcian Stenopterygius (Reptilia:Ichthyosauria). Palaeontology, 59:423-431. https://doi.org/10.1111/pala.12232
Fernández, M.S., Archuby, F., Talevi, M., and Ebner, R. 2005. Ichthyosaurian eyes: paleobiological information content in the sclerotic ring of Caypullisaurus (Ichthyosauria, Ophthalmosauria). Journal of Vertebrate Paleontology, 25:330-337. https://doi.org/10.1671/0272-4634(2005)025[0330:IEPICI]2.0.CO;2
Ferreira Da Veiga, O. 1958. Novos restos de “Ictyosauridae” e “Stenopterygidae” encontrados no Lias de Portugal. Serviços Geológicos de Portugal, 62:175-181.
Fischer,V., Bardet, N., Guiomar, M., and Godefroit, P. 2014. High diversity in Cretaceous ichthyosaurs from Europe prior to their extinction. PloS ONE 9:e84709. https://doi.org/10.1371/journal.pone.0084709
García-Ramos, J.C. and Piñuela, L. 2010. La ritmita de calizas y margas del Pliensbachiense, p. 21-40. In García-Ramos, J.C. (ed.), Las sucesiones margocalcáreas marinas del Jurásico Inferior y las series fluviales del Jurásico Superior. Acantilados de la playa de Vega (Ribadesella). Guía de la excursión A del V Congreso del Jurásico de España. Museo del Jurásico de Asturias, Colunga, Spain.
García-Ramos, J.C., Piñuela, L., and Lires, J. 2006. Atlas del Jurásico de Asturias, Ediciones Nóbel, Oviedo.
García-Ramos, J.C., Piñuela, L., and Rodríguez-Tovar, F.J. 2011. Post-workshop Fieldtrip Guide of the XI International Ichnofabric Workshop. Museo del Jurásico de Asturias, Colunga.
Godefroit, P. 1992. Présence de Leptopterygius tenuirostris (Reptilia, Ichthyosauria) dans le Lias moyen de Lorraine Belge. Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, 62:163-170.
Gómez, J.J. and Goy, A. 2000. Definition and organization of limestone-marl cycles in the Toarcian of the northern and east-central part of the Iberian Subplate (Spain), p. 301-310. In Hall, R.L. and Smith, P.L. (eds.), Advances in Jurassic Research 2000: GeoResearch Forum 6. Tech Publications, Zürich.
Hogler, J.A. 1992, Taphonomy and paleoecology of Shonisaurus popularis (Reptilia: Ichthyosauria). Palaios, 7:108-117. https://doi.org/10.2307/3514800
Huene, F. 1931. Neue Ichthyosaurier aus Württemberg. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, B 65:305-320.
Hungerbühler, A. and Sachs, S. 1996. Ein großer Ichthyosaurier aus dem Pliensbachium von Bielefeld: neue Einblicke in die Ichthyosaurier des mittleren Lias und das Gebiss von Temnodontosaurus . Bericht Naturwissenschaftlicher Verein für Bielefeld und Umgegend, 37:15-52.
Johnson, R. 1977. Size independent criteria for estimating relative age and the relationships among growth parameters in a group of fossil reptiles (Reptilia: Ichthyosauria). Canadian Journal of Earth Science, 14:1916-1924. https://doi.org/10.1139/e77-162
Lomax, D.R. 2016. A new leptonectid ichthyosaur from the LowerJurassic (Hettangian) of Nottinghamshire, England, UK, and the taxonomic usefulness of the ichthyosaurian coracoid. Journal of Systematic Palaeontology, 15:1-16. https://doi.org/10.1080/14772019.2016.1183149
Lomax, D.R. and Gibson, B.J.A. 2015. The first definitive occurrence of Ichthyosaurus and Temnodontosaurus (Reptilia: Ichthyosauria) in Nottinghamshire, England and a review of ichthyosaur specimens from the county. Proceedings of the Geologists’ Association, 126:554-563. https://doi.org/10.1016/j.pgeola.2015.05.006
Lomax, D.R. and Massare, J.A. 2012. The first Leptonectes (Reptilia: Ichthyosauria) with associated embryos, from Somerset, England. Paludicola, 8:263-276.
Lomax, D.R. and Massare, J.A. 2015. A new species of Ichthyosaurus from the Lower Jurassic of West Dorset, England. Journal of Vertebrate Paleontology, 35:e903260. https://doi.org/10.1080/02724634.2014.903260
Lydekker, R. 1888. Note on the classification of the Ichthyopterygia with a notice of two new species. Geological Magazine, 5:309-314. https://doi.org/10.1017/s0016756800181968
Maisch, M.W. 1998. A new ichthyosaur genus from the Posidonia Shale (Lower Toarcian, Jurassic) of Holzmaden, SW Germany with comments on the phylogeny of post-Triassic ichthyosaurs. Neues Jahrbuch für Geologie und Paleaontologie, Abhandlungen, 209:47-78.
Maisch, M.W. 1999. Leptonectiden und Temnodontosauriden aus dem Alpha- Ölschiefer (Sinemurium) von Baden-Württemberg (SW-Deutschland). Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, 8:490-512.
Maisch, M.W. and Reisdorf, A.G. 2006. Evidence for the longest stratigraphic range of a post-Triassic ichthyosaur: a Leptonectes tenuirostris from the Pliensbachian (Lower Jurassic) of Switzerland. Geobios, 39:491-505. https://doi.org/10.1016/j.geobios.2005.04.005
Martin, J.E., Fischer, V., Vincent, P., and Suan, G. 2012. A longirostrine Temnodontosaurus (Ichthyosauria) with comments on Early Jurassic ichthyosaur niche partitioning and disparity. Palaeontology, 5:995-1005. https://doi.org/10.1111/j.1475-4983.2012.01159.x
Martindale, R.C., Them, T.R., Gill, B.C., Marroquín, S.M., and Knoll, A.H. 2017. A new Early Jurassic (ca. 183 Ma) fossil Lagerstätte from Ya Ha Tinda, Alberta, Canada. Geology, 45:255-258. https://doi.org/10.1130/G38808.1
Massare, J.A. and Lomax, D.R. 2016a. A new specimen of Ichthyosaurus conybeari (Reptilia: Ichthyosauria) from Watchet, Somerset, U.K. and a re-examination of the species. Journal of Vertebrate Paleontology, 36:e1163264. https://doi.org/10.1080/02724634.2016.1163264
Massare, J.A. and Lomax, D.R. 2016b. Composite skeletons of Ichthyosaurus in historic collections. Paludicola, 10:207-250.
Massare, J.A. and Lomax. D.R. 2017. A taxonomic reassessment of Ichthyosaurus communis and I. intermedius and a revised diagnosis for the genus. Journal of Systematic Palaeontology, 16:263-277. https://doi.org/10.1080/14772019.2017.1291116
Maxwell, E., Fernández, M.S., and Schoch, R.R. 2012. First diagnostic marine reptile remains from the Aalenian (Middle Jurassic): a new ichthyosaur from southwestern Germany. PLoS ONE, 7(8):e41692. https://doi.org/10.1371/journal.pone.0041692
McGowan, C. 1989. Leptopterygius tenuirostris and other long-snouted ichthyosaurs from the English Lower Lias. Palaeontology, 32:409-427.
McGowan, C. 1993. A new species of large, long-snouted ichthyosaur from the English lower Lias. Canadian Journal of Earth Sciences, 30:1197-1204. https://doi.org/10.1139/e93-101
McGowan, C. 1996. The taxonomic status of Leptopterygius Huene, 1922 (Reptilia: Ichthyosauria). Canadian Journal of Earth Sciences, 33:439-443. https://doi.org/10.1139/e96-033
McGowan, C. and Milner, A.R. 1999. A new Pliensbachian ichthyosaur from Dorset, England. Palaeontology, 42:761-768. https://doi.org/10.1111/1475-4983.00096
McGowan, C. and Motani, R. 2003. Handbook of Paleoherpetology, Part 8 Ichthyopterygia. Verlag Dr. Friedrich Pfeil, München.
Merino Tomé, O., Suárez Rodríguez, A., Alonso Alonso, J.L., González Menéndez, L., Heredia Carballo, N., and Vallaure, M. 2011. Mapa Geológico Digital Continuo a Escala 1:50.000 (PLANGEODE). Instituto Geológico y Minero de España, Madrid.
Motani, R. 1999. Phylogeny of the Ichthyopterygia. Journal of Vertebrate Paleontology, 19:473-496. https://doi.org/10.1080/02724634.1999.10011160
Pereda Suberbiola, X., Ruiz-Omeñaca, J.I., Bardet, N., Piñuela, L., and García-Ramos, J.C. 2010. Wilhelm (Guillermo) Schulz and the earliest discoveries of dinosaurs and marine reptiles in Spain. Geological Society, London, Special Publications, 343:155-160. https://doi.org/10.1144/SP343.8
Ruiz-Omeñaca, J.I., García-Ramos, J.C., Piñuela, L., Bardet, N., Bermúdez- Rochas, D.D., Canudo, J.I., and Pereda Suberbiola, X. 2006. Restos directos de vertebrados del Jurásico de Asturias, p. 171-173. In Fernández-Martínez, E. (ed.), XXII Jornadas de la Sociedad Española de Paleontología y simposios de los proyectos PICG, 493, 503, 499, y467. Universidad de León. León.
Sauvage, H.E. 1897-1898. Vertébrés fossiles du Portugal. Contribution à l´étude des poissons et des reptiles du Jurassique et du Crétacé. Commissão do Serviço Geologico, Lisbon.
Schulz, G. 1858. Descripción geológica de la Provincia de Oviedo. Imprenta de D. José González, Madrid.
Stinnesbeck, W., Frey, E., Rivas, L., Pérez, J.P., Cartes, M.L., Soto, C.S., and Lobos, P.Z. 2014. A Lower Cretaceous ichthyosaur graveyard in deep marine slope channel deposits at Torres del Paine National Park, southern Chile. Geological Society of America Bulletin, 126:1317-1339. https://doi.org/10.1130/B30964.1
Suárez Vega, L.C. 1974. Estratigrafía del Jurásico en Asturias. Cuadernos de Geología Ibérica, 3:1-368.
Valenzuela, M., García-Ramos, J.C., González-Lastra, J., and Suárez de Centi, C. 1985. Sedimentación cíclica margo-calcárea de plataforma en el Lias de Asturias. Trabajos de Geología, Universidad de Oviedo, 15:45-52.
Valenzuela, M., García-Ramos, J.C., and Suárez de Centi, C. 1986. The Jurassic sedimentation in Asturias (N Spain). Trabajos de Geología, Universidad de Oviedo, 16:121-132.
Valenzuela, M., García-Ramos, J.C., and Suárez de Centi, C. 1989. La sedimentación en una rampa carbonatada dominada por tempestades, ensayos de correlación de ciclos y eventos en la ritmita margo-calcárea del Jurásico de Asturias. Cuadernos de Geología Ibérica, 13:217-235.
Williams, M., Benton, M.J., and Ross, A. 2015. The Strawberry Bank Lagerstätte reveals insights into Early Jurassic life. Journal of the Geological Society, 172:683-692. https://doi.org/10.1144/jgs2014-144
Zbyszewski, G. and Moitinho de Almeida, F. 1952. Restes d’Ichtyosauriens dans le Lias de S. Pedro de Muel. Serviços Geológicos de Portugal, 33:5-10.
