 New fossil cylindrical bark beetle (Zopheridae: Colydiinae: Gempylodini) from Eocene Baltic amber: An abnormal or intermediate form within Tenebrionoidea
New fossil cylindrical bark beetle (Zopheridae: Colydiinae: Gempylodini) from Eocene Baltic amber: An abnormal or intermediate form within Tenebrionoidea
Article number: 22.2.43
https://doi.org/10.26879/963
Copyright Paleontological Society, July 2019
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 29 January 2019. Acceptance: 13 June 2019
{flike id=2625}
ABSTRACT
Damzenia groehni gen. et sp. nov., preserved in Baltic amber is described as the first fossil representative of the tribe Gempylodini (Zopheridae: Colydiinae). In addition to the typical general characters of zopherids and colydiines, the newly described beetle possesses the unique tarsal formula 4-5-4 and distinct subcoxal lines on the metaventrite and abdominal ventrite 1. The tarsal formula of the Eocene fossil can be considered an intermediate stage between tenebrionoids with 5-5-4 and tenebrionoids with 4-4-4 tarsal formula. Computer reconstructions of the habitat for the newly described beetle created with the use of the Terragen4 program and the Xfrog software are provided.
Vitalii I. Alekseev. Shirshov Institute of Oceanology, Russian Academy of Sciences, Nahimovskiy prospekt 36, 117997. Moscow, Russia. alekseew0802@yahoo.com and Kaliningrad Regional Amber Museum, Marshal Vasilevskii square 1, 236016, Kaliningrad, Russia
Pavel I. Alekseev, Komarov Botanical Institute of the Russian Academy of Sciences, Prof. Popov str. 2, 197376. St. Petersburg, Russia. PAlekseev@binran.ru
Key words: Coleoptera; Paleogene; habitat reconstruction; fossil resin; new genus; new species
Alekseev, Vitalii I. and Alekseev, Pavel I. 2019. New fossil cylindrical bark beetle (Zopheridae: Colydiinae: Gempylodini) from Eocene Baltic amber: An abnormal or intermediate form within Tenebrionoidea. Palaeontologia Electronica 22.2.43A 1-10. https://doi.org/10.26879/963
palaeo-electronica.org/content/2019/2625-new-fossil-cylindrical-beetle
Copyright: July 2019 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/DF1997F0-AEEF-4945-9B53-F5EC8ED1DFB9
INTRODUCTION
Before the current study, eight fossilized Baltic amber species from five genera of zopherids belonging to Synchitini Erichson, 1845 (six species), and Pycnomerini Erichson, 1845 (two species), were described (Alekseev, 2017; Bukejs et al., 2019). All of these taxa were comparatively easily assigned to Recent genera and do not display any especially peculiar characters. A very interesting “intermediate” between the subfamilies Zopherinae and Colydiinae, Cretomysteria burmanica Deng, Ślipiński, Ren and Pang, 2017, and also belonging to the tribe Synchitini Erichson, 1845, new taxon Paleoendeitoma antennata Deng, Ślipiński, Ren and Pang, 2017, were described (Deng et al., 2017) from older, Mesozoic (Upper Cretaceous) Myanmar amber.
Colydium or Colydium-like specimens preserved in Baltic amber have been previously mentioned in the literature. For example, Berendt (1845: p. 56) listed “Colydium” under “Xylophaga”, the latter an informal name without nomenclatural standing. Giebel (1856: p. 125) recorded amber inclusions of the genus “Colydium Fabr.” within “55. Familie. Colydini”. Larsson (1978: p. 160) cited Berendt’s report of Colydium. Hieke and Pietrzeniuk (1984: p. 307) mentioned the presence of Colydium-like specimens, as well as the specimens labeled as “Colydium” in the Berendt collection of Baltic amber deposited in the Museum of Natural History (Germany, Berlin). All of the abovementioned determinations have to be considered provisional; the cylindrical-shaped representatives of different genera and families (e.g., Bothrideridae, Teredidae, Tenebrionidae, Zopheridae) may be inadvertently included in these reports. The most credible identification of the genus in Baltic amber is a single specimen of Colydium Fabr., Colydiidae (Kubisz, 2000: p. 227), which is deposited in the Baltic amber collection of the Museum of Natural History of the Institute of Systematics and Evolution of Animals in Kraków (Poland). The examination of recently collected material yielded a new “Colydium-like beetle” which is described in the present paper and tentatively placed in Gempylodini.
The tribe Gempylodini Sharp, 1893, includes extremely elongate and subcylindrical colydiids often found in the tunnels of wood-boring insects. The group is usually characterized by the combination of concealed antennal and mandibular insertions, broadly closed procoxal cavities, a non-articulated external spine at the apex of all tibiae, narrowly separated metacoxae (Lawrence, 1980), prothorax impressed laterally for reception of legs and antenna gradually clavate in some genera (Ivie et al., 2016). Two similarly elongate and cylindrical tribes of colydiine beetles differ from Gempylodini in having sublateral pronotal carinae and a 3-segmented, distinct antennal club (four genera of tribe Colydiini Erichson, 1842); or in having dorsally visible mandibular bases and a 2-segmented antennal club (the monogeneric tribe Nematidiini Sharp, 1894). Six known Recent genera of the quite diverse Gempylodini are distributed throughout the warmer parts of the Old World and occur also in the Neotropical Region (Lawrence, 1980; Ivie and Ślipiński, 1990; Ślipiński and Pal, 1997; Aoki et al., 2008; Makita, 2017): Aprostoma Guerin-Meneville, 1840, is known from Central and West tropical Africa and Madagascar; Gempylodes Pascoe, 1863, is restricted to Eastern and South-Eastern Asia; Mecedanum Erichson, 1845, is reported from Africa, Madagascar and Seychelles; Munaria Reitter, 1882, is known from East India and Australia; Pseudendestes Lawrence, 1980, is distributed in Japan, Taiwan, India, New Guinea and Australia; and Endestes Pascoe, 1863, occurs in Central and South America.
MATERIAL AND METHODS
The amber piece “GPIH 4946” containing the holotype is deposited in the museum of the Geological-Palaeontological Institute [Das Geologisch-Paläontologische Museum] of Hamburg University, now: CeNak, Centrum of Natural History [Centrum für Naturkunde], Germany and can be found in the separate collection of Carsten Gröhn under number “8586”.
Preparation of the amber specimen for examination and photography was performed by Jonas Damzen (Vilnius, Lithuania). The amber was cut and polished in order to better view the dorso-lateral and ventral sides of the specimen. Photographs were taken using a Canon 70D camera with a macro lens (Canon MPE-65mm). Extended depth of field at high magnifications was achieved by combining multiple images from a range of focal planes using Helicon Focus software. Measurements were taken using an ocular micrometer. The habitus reconstruction was made based on freehand drawing during examination of the original specimens. Pictures with the habitat reconstructions were rendered with the Terragen4 scenery generator program, while the models of Damzenia groehni gen. et sp. nov. and typical Eocene plants were created with the Xfrog Plants3.5 software library. All images were edited and assembled into figures using Adobe Photoshop CS8.
SYSTEMATIC PALAEONTOLOGY
Family ZOPHERIDAE Solier, 1834
Subfamily COLYDIINAE Billberg, 1820
Tribe GEMPYLODINI Sharp, 1893
Remarks. The beetle specimen from amber piece No. 4946 can be classified into Gempylodini due to the following characteristics: (1) antennal and mandibular insertions concealed; (2) pro-and mesocoxal cavities closed; (3) metacoxae narrowly separated by the acute abdominal intercoxal process; (4) 11-segmented antennae with weak 3-segmented club; (5) all tibiae toothed at outer apical angle; (6) frons above antenna forming fine ridge; (7) body strongly elongate, subcylindrical, glabrous; (8) intercoxal process of prosternum narrower than coxal cavity; (9) subantennal grooves absent; (10) scutellary striole absent; (11) ventrites 1-2 connate; (12) prothorax long, impressed laterally for reception of legs; (13) prothoracic hypomera without antennal cavities.
The specimen possesses doubled punctation on the elytra, long discrimen, unique tarsal formula 4-5-4 (all Colydiinae have tarsal formula 4-4-4, discussed below) and distinct subcoxal (femoral) lines (not known in other Colydinae, but present in different cucujoids, e.g., Languriinae, Silvanidae, Endomychidae, Coccinellidae, Bothrideridae, etc.). In spite of the last two contradictory characters, the new genus is with reservation placed in the extant tribe Gempylodini of Colydiinae. These characters (unusual tarsal formula, subcoxal lines) were treated as putatively primitive within Colydiinae.
Genus Damzenia V. Alekseev gen. nov.
zoobank.org/AE94A212-05EC-4268-A17F-097374E4B797
Type species. Damzenia groehni sp. nov.
Diagnosis. Damzenia gen. nov. can be differentiated from all other representatives of the tribe Gempylodini by the following combination of characters: habitus moderately elongate (approximately 4× as long as wide); antennal grooves on head absent; antenna with weakly distinct 3-segmented antennal club; pronotum without basal pits or carinae; prosternal process moderately broad, not convex, with raised lateral borders; procoxae separated; discrimen long; mesometaventral articulation dicondylic; elytral apices rounded; ventrite 5 simple. This genus differs from other gempylodines in having femoral (postcoxal) lines on metaventrite and abdominal ventrite 1 and unique tarsal formula 4-5-4.
There are no similarly elongate, cylindrical-shaped zopherids described from Baltic amber presently. The Baltic amber species Bitoma glaesisepulta Alekseev, 2015 (Zopheridae: Colydiinae: Synchitini) clearly differs from the new genus in having the antennal club distinctly 2-segmented, tarsi tetramerous, body length 2.75 mm, procoxal cavities open, head and anterior pronotal margin longitudinally rugose.
Within Gempylodini, the habitus of Damzenia gen. nov. resembles the New World Endestes Pascoe, 1863, and the Australasian Pseudendestes Lawrence, 1980. The new genus can be easily distinguished from both abovementioned genera by its non-carinate pronotum, as well as distinct elytral punctation and doubled teeth on the tibiae.
Remarks. The new genus resembles in some respects the tenebrionid species Yantaroxenos colydioides Nabozhenko, Kirejtshuk and Merkl, 2016 (Nabozhenko et al., 2016) of Tertiary age, but differs in glabrous body (squamiform pubescence on dorsum in Yantaroxenos), tarsal formula 4-5-4 (5-5-4 in Yantaroxenos), distinctly clubbed antennae (moniliform in Yantaroxenos), body length 4 mm (8.2 mm in Yantaroxenos), etc.
Etymology. The new genus is named after Mr. Jonas Damzen (Vilnius, Lithuania), an enthusiast and specialist in Baltic amber and the collector of the amber piece. The gender is feminine.
Composition. The new genus is monotypic, represented by the type species only. Therefore, the generic description considerably overlaps that of the species.
Damzenia groehni V. Alekseev sp. nov.
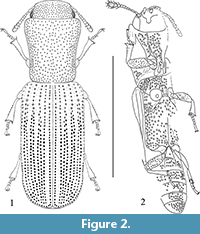
(Figure 1.1-5, Figure 2.1-2)
zoobank.org/96B999B1-3B69-4503-ACF5-AA2B2C7AAF3C
Type material. Holotype GPIH No. 4946 [coll. Gröhn No. 8586], adult, sex unknown, almost complete (right middle leg lacking), included in flattened amber piece, yellowish in colour, with approximate dimensions: 20×18×4 mm, and preserved without supplementary fixation. The ventral head area is partially obscured by milky foam in the surrounding amber. Syninclusions are represented by numerous stellate fagacean hairs, two complete specimens of Diptera (Nematocera: Sciaridae) and by four phoretic mites (Acari: Sarcoptiformes?) attached to the surface of the beetle, with the position including: to the base of left elytron, near the right mesocoxa, to abdominal ventrite 3 and to the left half of the metaventrite.
 Type strata. Baltic amber, Middle or Upper Eocene.
Type strata. Baltic amber, Middle or Upper Eocene.
Type locality. Yantarny settlement (formerly Palmnicken), Sambian (Samland) Peninsula, Kaliningrad Region, Russia.
Diagnosis. As stated for the new genus.
Description. Body elongate (3.8× as long as wide), subcylindrical, black, glabrous, shiny, punctured (Figure 1.1, 1.2, Figure 2.1). Total body length 4.03 mm; maximal body width 1.05 mm; pronotal length 1.25 mm; maximal pronotal width 1.0 mm; elytral length 2.4 mm.
Head: Strongly transverse, 0.5× as long as wide (measured ventrally including eyes), partially inserted into prothorax. Eyes lateral, flattened, flush with head, finely faceted, almost straight anteriorly, without intrafacetal setae. Fronto-clypeal suture not distinctly impressed. Anterior edge of clypeus convex. Frons weakly convex, forming fine ridge above antennal insertions and eyes, with moderately dense punctures. Distance between punctures 0.5 to 2.0× diameter of single puncture. Subantennal grooves not visible. Terminal labial and maxillary palpomeres fusiform, truncate. Antennae 11-segmented, stout, short, extending to apical third of pronotum, gradually widened, with 3-segmented antennal club. Antennomere 1 rounded, with at least its basal half concealed and not visible from above. Antennomeres 2 and 3 conical, approximately 2.5× as long as wide. Antennomere 3 narrower than antennomere 2. Antennomeres 4 to 7 subquadrate, subequal in shape; antennomeres 8 to 10 conical; antennomere 11 ovate, widely rounded. Antennomeres 9 to 11 (forming weak antennal club) bearing long, erect, fine setae.
 Thorax: Pronotum oblong (1.25× as long as wide), widest within anterior fourth of its length, finely bordered basally and laterally, punctate. Anterior and posterior angles rounded, not prominent. Anterior and posterior margins almost straight, lateral margins subparallel basally and distinctly widened in anterior half. Pronotal punctation moderate on the disc (distance between punctures about 1.0 to 3.0× diameter of one puncture), coarser and denser laterally, denser and smaller basally, smaller anteriorly. Prothorax long, widely impressed laterally for reception of legs. Prothoracic hypomera without antennal cavities. Prosternal intercoxal process moderately broad, with pair of symmetrical oblong grooves and with raised lateral borders, distinctly expanded apically, truncate. Maximum (apical) width of prosternal process equal to diameter of procoxal cavity. Procoxal cavities moderately wide separated (distance between coxae about 0.6× diameter of one coxa), closed externally. Pro-, meso-and metasternum punctate. Mesepisternum densely and coarsely punctured, triangular. Mesepimeron not visible from ventral side. Metepisternum narrow and almost completely concealed by elytral epipleuron. Mesocoxae separated (distance between coxae about 0.4× diameter of one coxa). Mesocoxal cavities closed. Mesometaventral articulation dicondylic. Metaventrite with weakly arcuate submesocoxal (femoral) lines (almost as long as mesocoxal diameter) and with long discrimen (about 0.6 × length of metaventrite). The ratio of lengths of prothorax to mesoventrite to metaventrite to abdomen (measured medially) is 6.0:2.5:6.5:8.7. Metacoxae slightly separated by very narrowly rounded (almost acute) intercoxal process of abdominal ventrite 1.
Thorax: Pronotum oblong (1.25× as long as wide), widest within anterior fourth of its length, finely bordered basally and laterally, punctate. Anterior and posterior angles rounded, not prominent. Anterior and posterior margins almost straight, lateral margins subparallel basally and distinctly widened in anterior half. Pronotal punctation moderate on the disc (distance between punctures about 1.0 to 3.0× diameter of one puncture), coarser and denser laterally, denser and smaller basally, smaller anteriorly. Prothorax long, widely impressed laterally for reception of legs. Prothoracic hypomera without antennal cavities. Prosternal intercoxal process moderately broad, with pair of symmetrical oblong grooves and with raised lateral borders, distinctly expanded apically, truncate. Maximum (apical) width of prosternal process equal to diameter of procoxal cavity. Procoxal cavities moderately wide separated (distance between coxae about 0.6× diameter of one coxa), closed externally. Pro-, meso-and metasternum punctate. Mesepisternum densely and coarsely punctured, triangular. Mesepimeron not visible from ventral side. Metepisternum narrow and almost completely concealed by elytral epipleuron. Mesocoxae separated (distance between coxae about 0.4× diameter of one coxa). Mesocoxal cavities closed. Mesometaventral articulation dicondylic. Metaventrite with weakly arcuate submesocoxal (femoral) lines (almost as long as mesocoxal diameter) and with long discrimen (about 0.6 × length of metaventrite). The ratio of lengths of prothorax to mesoventrite to metaventrite to abdomen (measured medially) is 6.0:2.5:6.5:8.7. Metacoxae slightly separated by very narrowly rounded (almost acute) intercoxal process of abdominal ventrite 1.
Elytra elongate (about 2.3× as long as combined widht), parallel-sided, with distinct humeri, jointly rounded apically, without costae. Scutellary striole absent. Each elytron with six puncture rows basally and nine puncture rows in the middle (striae 3, 4 and 5 duplicate within distal two-thirds of elytral length). Two interstrial intervals (between stria 2 and 3 basally; between stria 3 and 4 basally) convex, other intervals almost flat. All interstrial intervals with sparse, minute punctures. Elytral epipleura short, not reaching ventrite 1. Hind wings not apparent.
Legs: Short, robust. Femora sparsely and finely punctate. Meso-and metatibiae with groove along inner margin. Trochanter visible. Tibiae flattened and expanded apically; with 4-5 short spines along outer margin; with two narrow triangular teeth at outer apical angle; with two long, curved spurs at inner apical angle. Tarsal formula 4-5-4 (Figure 1.3-5). Relative length ratios of protarsomeres 1-4 equal to 5:3:3:9; relative length ratios of mesotarsomeres 1-5 equal to 5:3:3:3:9; relative length ratios of metatarsomeres 1-4 equal to 5:3:3:9. Claws long, falcate, simple, symmetrical, equal in size, acute.
Abdomen: With five visible ventrites, abdominal sutures distinct throughout length. Abdominal ventrites 1-2 more solidly fused together than ventrites 3-5. Ventrite length ratios (measured medially) are 30:15:12:10:20. Ventrite 1 with long, almost straight submetacoxal lines (Figure 1.5, Figure 2.2). Intercoxal process of ventrite 1 acute, triangular. Ventrites 1 to 4 moderately punctate, punctation of ventrite 5 denser (distance between punctures 0.3 to 1.0× diameter of puncture). Ventrite 5 simple, without preapical groove, convex.
Remarks. The scutellar shield is not visible due to flow lines and expansion cracks in the surrounding amber. The details of the mouthparts are hidden by milky, opaque amber around the oral cavity and not sufficiently visible for description. The medially doubled elytral punctation and two convex interstrial intervals are considered as putatively reduced elytral carination: more developed elytral carinae in other representatives of the new genus could be expected in future studies.
Etymology. The specific epithet is a patronym formed from the surname of Mr. Carsten Gröhn (Glinde, Germany), an enthusiast and specialist in Baltic amber, who donated the holotype to the museum of GPIH in CeNak.
DISCUSSION
Tarsal formula of the beetle. The most unusual and enigmatic feature of the beetle under study is its tarsal formula. Damzenia groehni gen. et sp. nov. has pentamerous mesotarsi and tetramerous pro-and metatarsi (i.e., tarsal formula 4-5-4). Members of the superfamily Tenebrionoidea Latreille, 1802, are generally characterized by tarsal formula of 5-5-4 (an autapomorphic character). Smaller taxa from this superfamily (belonging to distinct, independent groups) possess tarsal formula 4-4-4 with one tarsomere reduced in all tarsi (e.g., subfamily Colydiinae and several Zopherinae within Zopheridae; the entire families Cisidae and Prostomidae, several groups in Salpingidae, Ripiphoridae, Ulodidae, and Tenebrionidae). Only representatives of the family Mycetophagidae and some Phrenapatinae (Tenebrionidae) possess sexually dimorphic tarsal formula: the male 3-4-4 and the female 4-4-4 in Mycetophagidae; the male 3-3-3 and the female 4-4-4 in Phrenapatinae. Given the known tarsal counts and their patterns, the tarsal formula of the new fossil species is noteworthy in the context of the superfamily Tenebrionoidea.
On the one hand, the tarsal formula 4-5-4 has not been reported for the extant or fossil representatives of the superfamily Tenebrionoidea. The tarsal formula 4-5-4 is known for several representatives of the genus Augasmus Motschulsky, 1858 (superfamily Cucujoidea: family Phalacridae) only (Gimmel 2013).
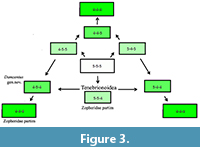 On the other hand, the tarsal formula 4-4-4 is derived from the ancestral tarsal formula 5-5-5. The most primitive, archaic and basic character state of the tarsi of Coleoptera is its pentamerous condition. During the evolutionary course of tarsal oligomerization (gradual process of tarsomere number reduction), each tarsus can lose tarsomeres and become pseudotetramerous, tetramerous, pseudotrimerous, trimerous etc. Tarsomeres can be reduced separately on one leg-pair, usually tarsomere-by-tarsomere. The primitive condition of 5-5-5 tarsi and the relatively derived condition of 4-4-4 tarsi allow six mathematically possible conditions [23=8]: 5-5-4, 5-4-4, 4-5-4, 4-4-5, 4-5-5 and 4-5-4. There are three principally possible intermediate character states in two stages and six ways of forming the tarsal formula 4-4-4 from 5-5-5 (Figure 3.1). The tarsal formula 4-4-4 can be obtained from the basic for Tenebrionoidea tarsal formula 5-5-4 by two putative ways: by the way of 5-4-4 stage or by the way 4-5-4 stage. Both intermediate stages unknown in extant Tenebrionoidea. The Eocene fossil Damzenia groehni gen. et sp. nov. shows the intermediate tarsal 4-5-4 formula and can be considered an intermediate stage between tenebrionoids with 5-5-4 and tenebrionoids with 4-4-4 tarsal formula.
On the other hand, the tarsal formula 4-4-4 is derived from the ancestral tarsal formula 5-5-5. The most primitive, archaic and basic character state of the tarsi of Coleoptera is its pentamerous condition. During the evolutionary course of tarsal oligomerization (gradual process of tarsomere number reduction), each tarsus can lose tarsomeres and become pseudotetramerous, tetramerous, pseudotrimerous, trimerous etc. Tarsomeres can be reduced separately on one leg-pair, usually tarsomere-by-tarsomere. The primitive condition of 5-5-5 tarsi and the relatively derived condition of 4-4-4 tarsi allow six mathematically possible conditions [23=8]: 5-5-4, 5-4-4, 4-5-4, 4-4-5, 4-5-5 and 4-5-4. There are three principally possible intermediate character states in two stages and six ways of forming the tarsal formula 4-4-4 from 5-5-5 (Figure 3.1). The tarsal formula 4-4-4 can be obtained from the basic for Tenebrionoidea tarsal formula 5-5-4 by two putative ways: by the way of 5-4-4 stage or by the way 4-5-4 stage. Both intermediate stages unknown in extant Tenebrionoidea. The Eocene fossil Damzenia groehni gen. et sp. nov. shows the intermediate tarsal 4-5-4 formula and can be considered an intermediate stage between tenebrionoids with 5-5-4 and tenebrionoids with 4-4-4 tarsal formula.
Unfortunately, only one specimen of this new taxon is available for study, and the second middle leg on the unique exemplar is lost. For objectivity, three possible explanations must be considered:
1. The exemplar is abnormal with one extra mesotarsomere (teratological deformity), and the normal tarsal formula of the species should be 4-4-4.
2. One (first?) protarsomere is extremely small and hidden (not visible at the maximum available magnification of 87.5×; the protarsi are pseudotetramerous, and the real tarsal formula should be 5-5-4.
3. The fossil record preserves an extinct transitional morphological character. The real tarsal formula is 4-5-4 (as it is seen); this is an evolutionary intermediate condition of the gradual reduction in number of tarsomeres in the zopherid lineage from the ancestral state “tarsal formula 5-5-4” (most members of the subfamily Zopherinae: tribes Usechini, Monommatini, Phellopsini, Zopherini) to the modern state “tarsal formula 4-4-4” (the subfamily Colydiinae and several genera from Pycnomerini Erichson, 1845, and Latometini Ślipiński and Lawrence, 1999).
Additional specimens of the species (which may be found in future) will further clarify the situation of this unique tarsomere number. Careful examination of the head external characters, possibly extruded genitalia or wings on other specimens can confirm or refute the seemingly archaic and somewhat intermediate position of the taxon and determine its real taxonomic status.
 Habitat in the “amber forest”. Baltic amber generally occurs in the Upper Eocene (Priabonian) marine sediments (Aleksandrova and Zaporozhets, 2008). Huge volumes of conifer resin were primarily deposited in the forest soil and later accumulated in mud layers after transgression of the sea. The Eocene “amber forest” was represented by a species-rich, humid, thermophilic mixed forest, where Fagaceae, Pinaceae and Cupressaceae plants dominated (Alekseev and Alekseev, 2016). Also the Lauraceae, Palmaceae and Magnoliaceae were usual elements in this forest. Similar to extant beetles of the tribe Gempylodini, Damzenia gen. nov. was possibly connected with the dried-out, standing and/or partially destroyed, lying trunks of old trees. The presumable climax character of the “amber forest” community is further supported by the diverse assemblage of xylophagous, xylomycetophagous beetles and their predators (e.g., the newly described beetle, and cerambycids, melandryids, ptinids, salpingids, zopherids, etc., also known from Baltic amber), whose habitat requires various and numerous trees reaching their maximum size and age. When the known flora from previous publications is combined with the community stage inferred from beetles and other fossil insects in the amber assemblage, it leads to a relatively comprehensive model of the ancient forest (Figure 4.1-2).
Habitat in the “amber forest”. Baltic amber generally occurs in the Upper Eocene (Priabonian) marine sediments (Aleksandrova and Zaporozhets, 2008). Huge volumes of conifer resin were primarily deposited in the forest soil and later accumulated in mud layers after transgression of the sea. The Eocene “amber forest” was represented by a species-rich, humid, thermophilic mixed forest, where Fagaceae, Pinaceae and Cupressaceae plants dominated (Alekseev and Alekseev, 2016). Also the Lauraceae, Palmaceae and Magnoliaceae were usual elements in this forest. Similar to extant beetles of the tribe Gempylodini, Damzenia gen. nov. was possibly connected with the dried-out, standing and/or partially destroyed, lying trunks of old trees. The presumable climax character of the “amber forest” community is further supported by the diverse assemblage of xylophagous, xylomycetophagous beetles and their predators (e.g., the newly described beetle, and cerambycids, melandryids, ptinids, salpingids, zopherids, etc., also known from Baltic amber), whose habitat requires various and numerous trees reaching their maximum size and age. When the known flora from previous publications is combined with the community stage inferred from beetles and other fossil insects in the amber assemblage, it leads to a relatively comprehensive model of the ancient forest (Figure 4.1-2).
ACKNOWLEDGEMENTS
The authors are sincerely grateful to Mr. J. Damzen (Vilnius, Lithuania) for permission to use his excellent photographs, to Dr. E.A. Sidorchuk (Borissiak Paleontological Institute of the Russian Academy of Sciences, Moscow, Russia) for help in identification of acari specimen, and to Dr. D.A. Pollock (Eastern New Mexico University, Portales, USA) for a native English speaker proofread of our manuscript. Mr. C. Gröhn (Glinde, Germany) is acknowledged for donating the specimens to GPIH in CeNak (Hamburg, Germany). We thank two anonymous reviewers for valuable suggestions that improved the manuscript.
The study of the first author was done with support of the state assignment of IO RAS (Theme No. 0149-2019-0013). The study of the second author was carried out within the framework of the institutional research project (No. AAAA-A19-119021190031-8) of the Komarov Botanical Institute of the Russian Academy of Sciences.
REFERENCES
Aleksandrova, G.N. and Zaporozhets, N.I. 2008. Palynological characteristics of the Upper Cretaceous and Paleogene deposits on the west of the Sambian Peninsula (Kaliningrad Region), part 1. Stratigraphy and Geological Correlation, 16(3):75-96 (In Russian). https://doi.org/10.1134/s0869593808030052
Alekseev, V.I. 2017. Coleoptera from the Middle-Upper Eocene European ambers: Generic composition, zoogeography and climatic implications. Zootaxa, 4290(3):401-443. https://doi.org/10.11646/zootaxa.4290.3.1
Alekseev, V.I. and Alekseev, P.I. 2016. New approaches for reconstruction of the ecosystem of an Eocene amber forest. Biology Bulletin, 43(1):75-86. https://doi.org/10.1134/s1062359016010027
Aoki, J., Narukawa, N., and Tanaka, I. 2008. On Pseudendestes andrewesi (Grouvelle) found from Japan (Zopheridae). Coleopterists’ News, 164:1-3 (In Japanese)
Berendt, G. 1845. Die im Bernstein befindlichen organischen Reste der Vorwelt gesammelt in Verbindung mit mehreren bearbeitet. Erster Band. Abtheilung I. Der Bernstein und die in ihm befindlichen Pflanzenreste der Vorwelt. Nikolaische Buchhandlung, Danzig. https://doi.org/10.5962/bhl.title.66910
Bukejs, A., Alekseev, V.I., Cooper, D.M.L., King, G.A., and McKellar, R.C. 2019. A new fossil species of Pycnomerus Erichson (Coleoptera: Zopheridae) from Baltic amber, and a replacement name for a Recent North American congener. Zootaxa, 4550(4):565-572. https://doi.org/10.11646/zootaxa.4550.4.6
Deng, C., Ślipiński, A., Ren, D., and Pang, H. 2017. The first Mesozoic colydiid beetles (Coleoptera: Zopheridae: Colydiinae) from the Upper Cretaceous amber of Myanmar. Cretaceous Research, 78:71-77. https://doi.org/10.1016/j.cretres.2017.04.010
Giebel, C.G.A. 1856. Geologische Übersicht der vorweltlichen Insecten. Zeitschrift für die Gesammten Naturwissenschaften, 8:174-188.
Gimmel, M. L. 2013. Genus-level revision of the family Phalacridae (Coleoptera: Cucujoidea). Zootaxa, 3605(1):1-147. https://doi.org/10.11646/zootaxa.3605.1.1
Hieke, F. and Pietrzeniuk, E. 1984. Die Bernstein-Käfer des Museums für Naturkunde, Berlin (Insecta, Coleoptera). Mitteilungen aus dem Zoologischen Museum Berlin, 60:297-326.
Ivie, M.A., Lord, N.P., Foley, I.A., and Ślipiński, S.A. 2016. Colydiine genera (Coleoptera: Zopheridae: Colydiinae) of the New World: A key and nomenclatural acts 30 years in the making. The Coleopterists Bulletin, 70(4):755-788. https://doi.org/10.1649/0010-065X-70.4.755
Ivie, M.A. and Ślipiński, S.A. 1990. Catalog of the genera of world Colydiidae (Coleoptera). Annales Zoologici (Warszawa), 43(suppl. 1):1-32.
Kubisz, D. 2000. Fossil beetles (Coleoptera) from Baltic amber in the collection of the Museum of Natural History of ISEA in Krakow. Polish Journal of Entomology, 69(2):225-230.
Larsson, S.G. 1978. Baltic Amber-A Palaeobiological Study. Volume 1. Scandinavian Science Press Ltd., Klampenborg, Denmark.
Lawrence, J.F. 1980. A new genus of Indo-Australian Gempylodini with notes on the constitution of the Colydiidae (Coleoptera). Journal of the Australian Entomological Society, 19:293-310. https://doi.org/10.1111/j.1440-6055.1980.tb00989.x
Makita, Sh. 2017. New distributional records of Pseudendestes andrewesi (Grouvelle, 1908) (Coleoptera: Zopheridae) from Taiwan. Taiwanese Journal of Entomological Studies, 2(3):8-12.
Nabozhenko, M., Kirejtshuk, A.G., and Merkl, O. 2016. Yantaroxenos colydioides gen. et sp. nov. (Tenebrionidae: Lagriinae) from Baltic amber. Annales Zoologici (Warszawa), 66(4):563-566. https://doi.org/10.3161/00034541ANZ2016.66.4.008
Ślipiński, S.A. and Lawrence, J.F. 1999. Phylogeny and classification of Zopheridae sensu novo (Coleoptera: Tenebrionoidea) with a review of the genera of Zopherinae (excluding Monommatini). Annales Zoologici (Warszawa), 49:1-53.
Ślipiński, S.A. and Pal, T.K. 1997. A review of the genus Aprostoma Guerin-Meneville (Coleoptera: Colydiidae). Annales Zoologici (Warszawa), 47(1-2):93-105.
