

Article Search
Volume 27.1
January–April 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
See all articles in 26.2 May-August 2023
See all articles in 26.1 January-April 2023
 Floréal Solé. D.O. Earth and history of Life, Department of Paleontology, Royal Belgian Institute of Natural Sciences, Rue Vautier 29, B-1000 Brussels, Belgium. fsole@naturalsciences.be
Floréal Solé. D.O. Earth and history of Life, Department of Paleontology, Royal Belgian Institute of Natural Sciences, Rue Vautier 29, B-1000 Brussels, Belgium. fsole@naturalsciences.be
He is researcher in Palaeontology at the Royal Belgian Institute of Natural Sciences (Brussels, Belgian). He has a PhD in Vertebrate Paleontology from Muséum National d'Histoire Naturelle (Paris, France). He is interested in the evolution, phylogeny, and biogeography of the earliest carnivorous mammals (Hyaenodonta, Carnivoramorpha, Mesonychidae). His research also focuses on the impacts of the climatic changes of the Paleogene on the emergence and evolution of these particular mammals.

 El Mabrouk Essid. Office National des Mines (ONM), 24 rue 8601, 2035 La Charguia, Tunis BP: 215 - 1080 Tunis, Tunisia. mab_04@yahoo.fr
El Mabrouk Essid. Office National des Mines (ONM), 24 rue 8601, 2035 La Charguia, Tunis BP: 215 - 1080 Tunis, Tunisia. mab_04@yahoo.fr
He is engineer in Geology and cartographer. He is head of the Documentation Service and Data bases. This service is in charge of the management and monitoring of activities of the documentation center of the National Office of Mines (Tunisia).
 Wissem Marzougui. Office National des Mines (ONM), 24 rue 8601, 2035 La Charguia, Tunis BP: 215 - 1080 Tunis, Tunisia. wi_marzougui@yahoo.fr
Wissem Marzougui. Office National des Mines (ONM), 24 rue 8601, 2035 La Charguia, Tunis BP: 215 - 1080 Tunis, Tunisia. wi_marzougui@yahoo.fr
He is an engineer in geological sciences specialized in structural geology. He realized numerous geological and thematic maps at different scales. He also provides geological expertise.
 Rim Temani. Office National des Mines (ONM), 24 rue 8601, 2035 La Charguia, Tunis BP: 215 - 1080 Tunis, Tunisia. rim.temani@onm.nat.tn
Rim Temani. Office National des Mines (ONM), 24 rue 8601, 2035 La Charguia, Tunis BP: 215 - 1080 Tunis, Tunisia. rim.temani@onm.nat.tn
She is a biostratigrapher (Foraminifera and Ostracoda), in the geological department at the National Office of Mines in Tunisia. She got her MSc [earth sciences] degrees in the Faculty of Mathematical, Physical and Natural Sciences of Tunis. Her PhD Thesis focused on Post-Tortonian ostracoda in Eastern Tunisia. She is mainly working on Upper Cretaceous and Tertiary sediments. Her research focuses on Messinian, Pliocene and Quaternary with a special emphasis on, quantitative analysis, paleoenvironmental reconstruction and climatic interpretation, in Tunisia and in the Mediterranean Sea.
 Hayet Khayati Ammar. Office National des Mines (ONM), 24 rue 8601, 2035 La Charguia, Tunis BP: 215 - 1080 Tunis, Tunisia. khayatihayet@yahoo.fr
Hayet Khayati Ammar. Office National des Mines (ONM), 24 rue 8601, 2035 La Charguia, Tunis BP: 215 - 1080 Tunis, Tunisia. khayatihayet@yahoo.fr
She is engineer in Micropalaeontology. She is assistant director of Geological Studies in the Department of Geology of the National Office of Mines (Tunisia). This service manages the activities of the paleontology department such as the sedimentological studies, and those of the thematic studies service (production of geological maps of Tunisia, notably indicating the natural hazards ).
 Mhammed Mahboubi. Laboratoire de Paléontologie stratigraphique et Paléoenvironnement, Université d’Oran, B.P. 1524 El M’naouer, Oran 31000, Algeria. mahboubi.med@gmail.com
Mhammed Mahboubi. Laboratoire de Paléontologie stratigraphique et Paléoenvironnement, Université d’Oran, B.P. 1524 El M’naouer, Oran 31000, Algeria. mahboubi.med@gmail.com
He is Professor of Geology in the Department of Earth Sciences, University of Oran 2 (Algeria). His research axis involves the stratigraphic study of the Paleocene and Eocene sediments recorded in the continental basins of southwestern Algeria.
 Laurent Marivaux. Laboratoire de Paléontologie, Institut des Sciences de l’Évolution de Montpellier (ISE-M, UMR 5554, UM, CNRS, IRD, EPHE), c.c. 064, Université de Montpellier, Place Eugène Bataillon, F-34095 Montpellier Cedex 05, France. laurent.marivaux@umontpellier.fr
Laurent Marivaux. Laboratoire de Paléontologie, Institut des Sciences de l’Évolution de Montpellier (ISE-M, UMR 5554, UM, CNRS, IRD, EPHE), c.c. 064, Université de Montpellier, Place Eugène Bataillon, F-34095 Montpellier Cedex 05, France. laurent.marivaux@umontpellier.fr
He is CNRS researcher in paleontology and head of the Paleontology Department at the "Institut des Sciences de l'Evolution de Montpellier". He has a PhD in Vertebrate Paleontology from the University of Montpellier. His research focuses on macroevolutionary, phylogenetic, historical biogeography and paleobiological aspects of the early Tertiary adaptive radiations of some mammal groups (especially Rodentia, Primates, Dermoptera and Chiroptera, and others in collaboration), with a special emphasis on the role of the geodynamic and paleoenvironment changes on their emergence, dispersals and evolution. His is primarily a field paleontologist working in Paleo- and Neotropics.
 Monique Vianey-Liaud. Laboratoire de Paléontologie, Institut des Sciences de l’Évolution de Montpellier (ISE-M, UMR 5554, UM, CNRS, IRD, EPHE), c.c. 064, Université de Montpellier, Place Eugène Bataillon, F-34095 Montpellier Cedex 05, France. monique.vianey-liaud@umontpellier.fr
Monique Vianey-Liaud. Laboratoire de Paléontologie, Institut des Sciences de l’Évolution de Montpellier (ISE-M, UMR 5554, UM, CNRS, IRD, EPHE), c.c. 064, Université de Montpellier, Place Eugène Bataillon, F-34095 Montpellier Cedex 05, France. monique.vianey-liaud@umontpellier.fr
She works in the "Département Forme, Institut des Sciences de l'Evolution » of the University of Montpellier, as an emeritous professor of Paleontology. Her researches concern early adaptive radiations of mammals, mainly European Paleogene rodents (theridomorphs for the most) and in a lesser extent, lagomorphs, North African Paleogene rodents, Paleogene multituberculates and Dinosaur eggshells. She actively participates in field work in Morocco and in the South of France (Quercy, Corbières, Provence) (and in Tunisia until 2011).
 Rodolphe Tabuce. Laboratoire de Paléontologie, Institut des Sciences de l’Évolution de Montpellier (ISE-M, UMR 5554, UM, CNRS, IRD, EPHE), c.c. 064, Université de Montpellier, Place Eugène Bataillon, F-34095 Montpellier Cedex 05, France. rodolphe.tabuce@umontpellier.fr
Rodolphe Tabuce. Laboratoire de Paléontologie, Institut des Sciences de l’Évolution de Montpellier (ISE-M, UMR 5554, UM, CNRS, IRD, EPHE), c.c. 064, Université de Montpellier, Place Eugène Bataillon, F-34095 Montpellier Cedex 05, France. rodolphe.tabuce@umontpellier.fr
He is CNRS researcher in paleontology. He has a PhD in vertebrate paleontology from the University of Montpellier. His research focuses on early Tertiary emergence, diversification, and paleobiogeographic history of Afro-Arabian mammals (notably afrotherians). He is also working on European late Cretaceous eutherians and intra-European faunal turnover during the early Eocene hyperthermals.
FIGURE 1. 1, localisation of the African fossiliferous localities that yielded Paleocene and Eocene hyaenodonts (see Table 1); 1: Ouled Abdoun Basin (Morocco); 2: Ouarzazate Basin (Morocco); 3: El Kohol (Algeria); 4: Gour Lazib (Algeria); 5: Chambi (Tunisia); 6: Black Crow locality from Sperrgebiet (Namibia); 7: Bir el Ater (Algeria); 8: Dur at-Talah (Libya); 9: Fayum Province = Birket Qarun Locality 2, Jebel Qatrani, and Qasr el-Sagha Formation (Egypt); 10: Taqah (Oman). 2, correlation of Eocene localities including hyaenodonts (see Table 1) [adapted from Coster et al. (2012, figure 1) and Seiffert (2010, figure 2.1)]. The black circles ( 1) and the names in Bold Italic ( 2) identify the localities under study. Abbreviations: Barton.= Bartonian; Priabon.= Priabonian; Seland.= Selandian; Thanet.= Thanetian.
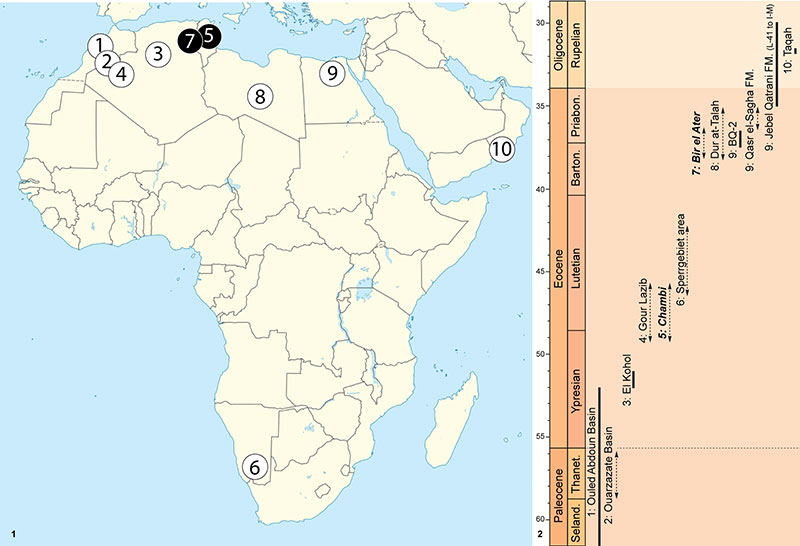
FIGURE 2. Furodon crocheti Solé et al., 2014b from Chambi CBI-1 (Tunisia). 1-2, CBI-1-550, right P2 in lingual ( 1) and occlusal ( 2) views. 3-4, CBI-1-551, right maxillary fragment bearing a P2 in labial ( 3) and occlusal ( 4 ) views. 5-6, CBI-1-552, right M1 in labial ( 5) and occlusal ( 6) views. 7-9, CBI-1-612, right dp3 in lingual ( 7), labial ( 8) and occlusal ( 9) views. 10-12, CBI-1-554, left m3 in lingual ( 10), labial ( 11) and occlusal ( 12) views. 13-15, CBI-1-553, right m1 in lingual ( 13), labial ( 14) and occlusal ( 15) views. Abbreviations: Hd, Hypoconulid; Par., Paraconid; Pro., Protoconid; Psd, Protostylid. Scale bar equals 5 mm.
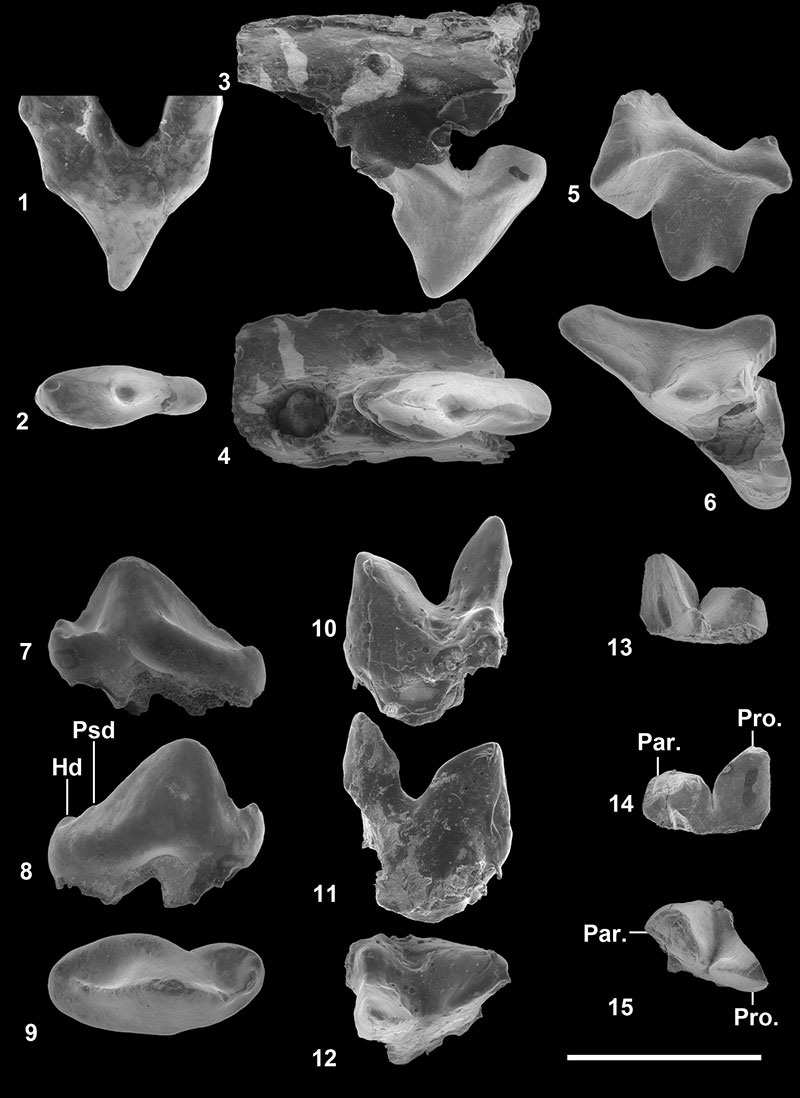
FIGURE 3. Parvavorodon gheerbranti Solé et al., 2014b from Chambi CBI-1 (Tunisia). 1-2, CBI-1-555, left P3 in labial ( 1) and occlusal ( 2) views. 3-4, CBI-1-556, left M1or2 in labial ( 3) and occlusal ( 4) views. 5-7, CBI-1-557, left p2 in lingual ( 5), labial ( 6) and occlusal ( 7) views. 8-10, CBI-1-558, right m1 in lingual ( 8), labial ( 9) and occlusal ( 10) views. 11-13, CBI-1-559, left m3 in lingual ( 11), labial ( 12) and occlusal ( 13) views. Scale bar equals 5 mm.
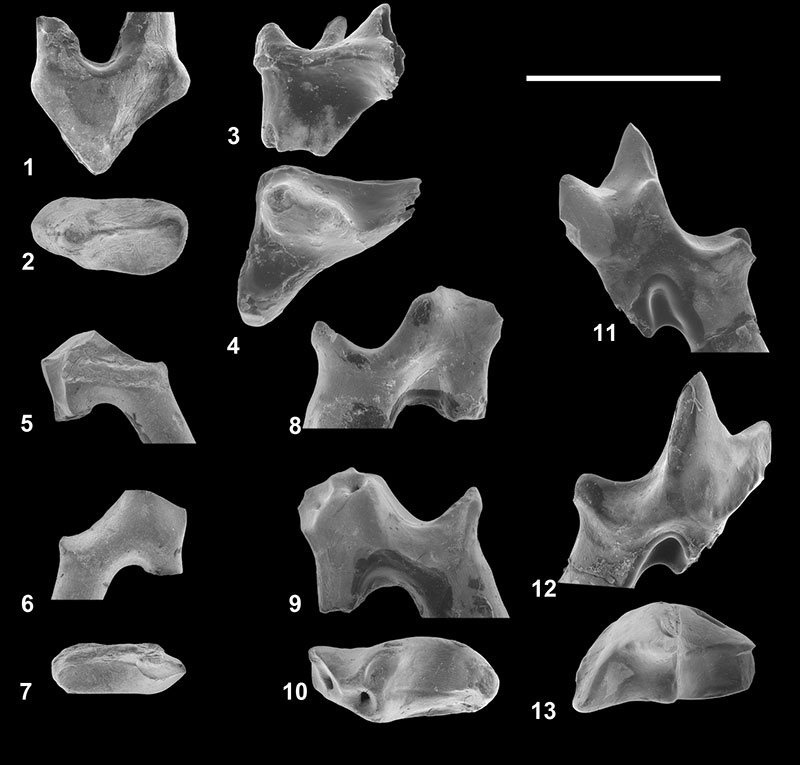
FIGURE 4. Parvavorodon gheerbranti Solé et al., 2014b from Chambi CBI-1 (Tunisia). 1-6, CBI-1-613, left dentary bearing dp4, m1, alveoli of dp3 and m2, and distal alveolus of dp2? in labial ( 1) occlusal ( 2), occlusal based on a CT-scan reconstruction ( 3), occlusal close-up ( 4), labial close-up ( 5) and lingual close-up ( 6) views. The black arrows on 2 indicate the two foramina related to the eruption of the permanent teeth (see text). Scale bar equals 2 mm.
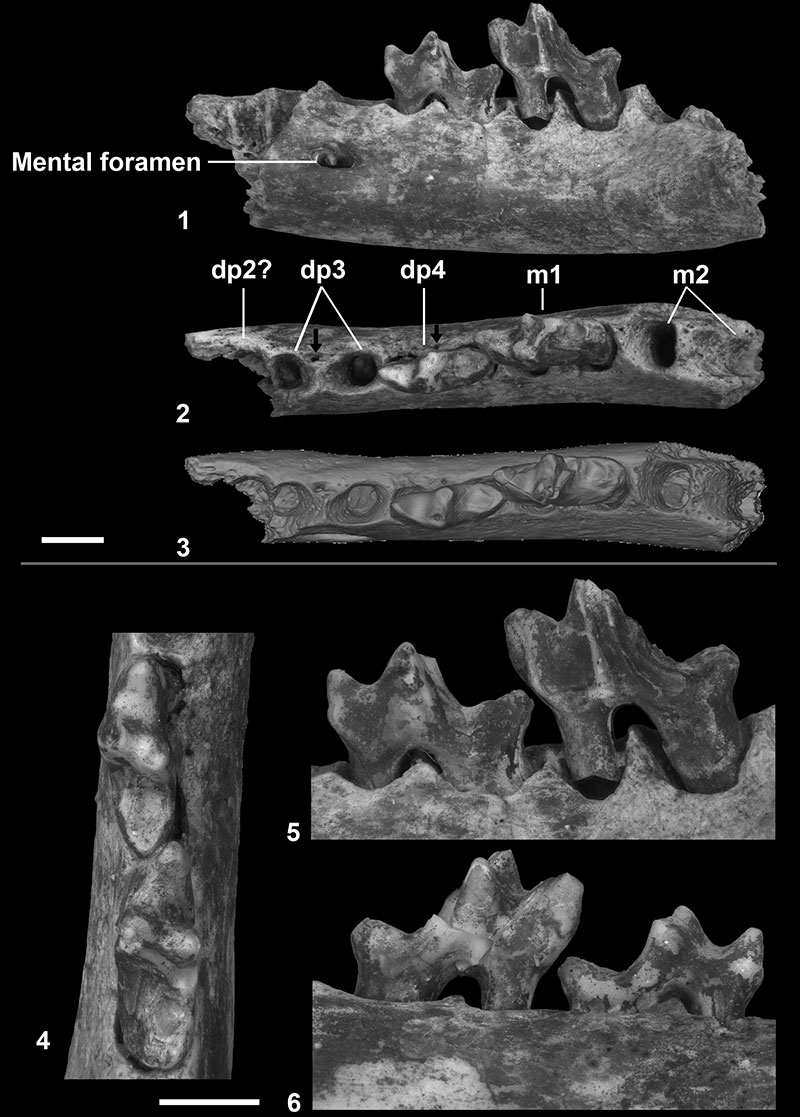
FIGURE 5. CT-scan reconstruction of Parvavorodon gheerbranti Solé et al., 2014b from Chambi CBI-1 (Tunisia); CBI-1-613, left dentary bearing dp4, m1, alveoli of dp3 and m2, and distal alveolus of dp2?. 1, occlusal view; 2, parasagittal slice showing the roots of the dp4 and m1. Note the absence of germ below dp3 and dp4. Scale bar equal 2 mm.
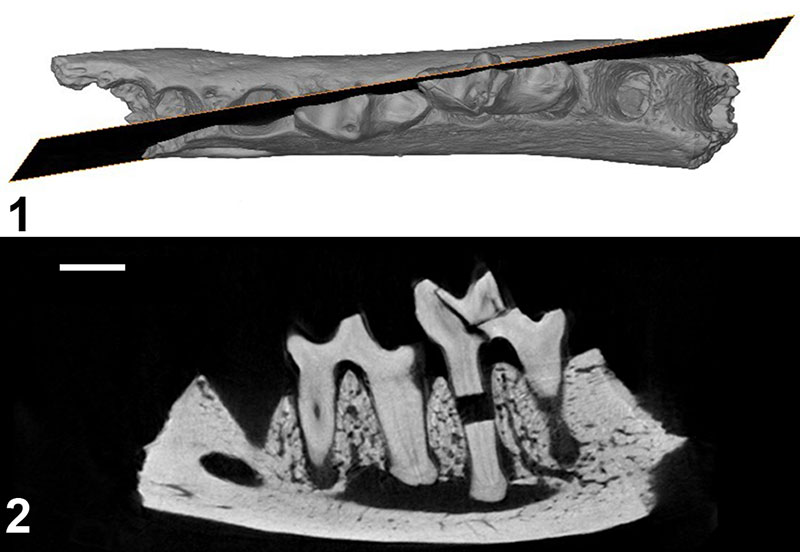
FIGURE 6. Hyainailourinae indet. from Bir el Ater (Algeria). 1-2, UON 84-359, left P3 in occlusal ( 1) and labial ( 2) views. Scale bar equals 5 mm.
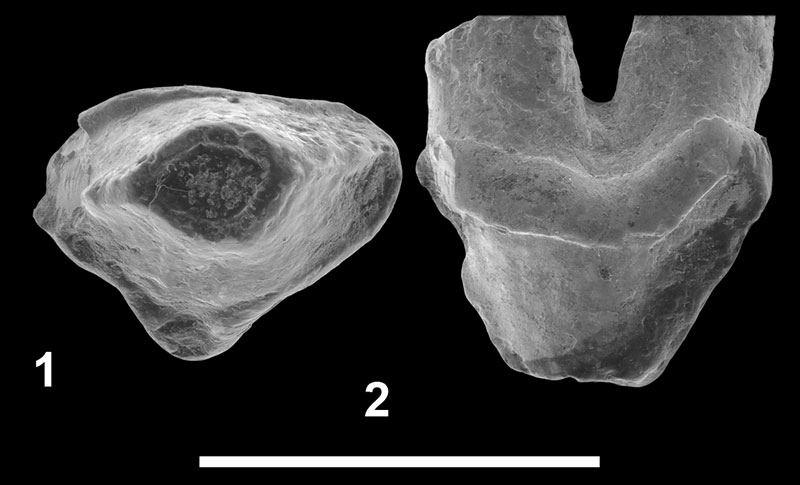
FIGURE 7. Masrasector cf. ligabuei from Bir el Ater (Algeria). 1-2, UON 84-360, right M1or2 in occlusal (1) and labial ( 2) views. 3-4, UON 84-361, right M3 in lingual ( 3) and occlusal ( 4) views. 5-7, UON 84-397, right p3 in occlusal ( 5), lingual ( 6) and labial ( 7) views. Scale bar equals 5 mm.
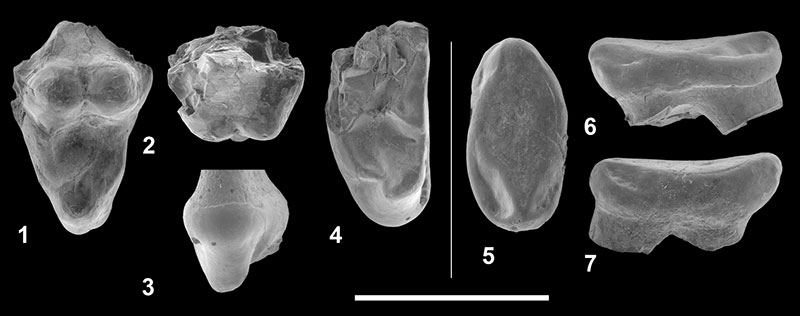
FIGURE 8. Teratodontinae indet. 1 from Chambi CBI-1 (Tunisia). 1-2, CBI-1-614, right P4 in occlusal ( 1) and labial ( 2) views. Scale bar equals 5 mm.
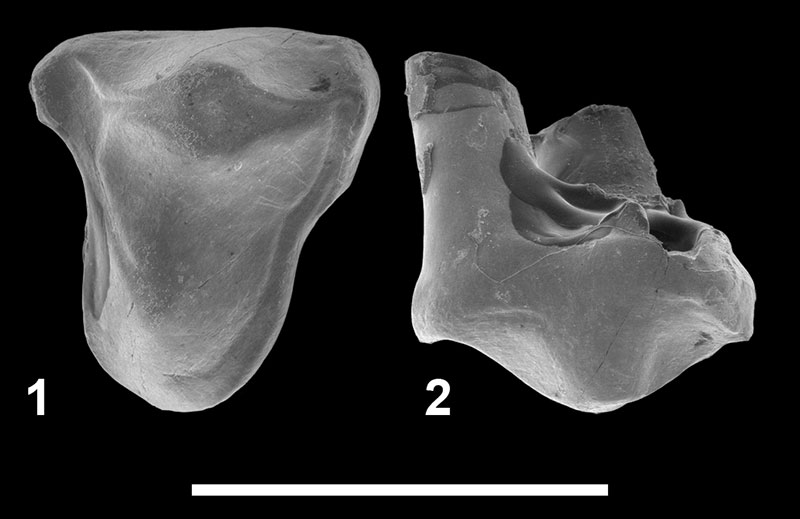
FIGURE 9. 1, geographic distribution of the Faunal Assemblages (see text). 2, stratigraphic distribution of Tinerhodon and hyaenodont subfamilies in Africa during Paleocene and Eocene, and of the faunal assemblages regarding the main climatic events and mammal appearances. Note the gap in the fossil record of the Koholiinae. Abbreviations: Barton.= Bartonian; EECO: Early Eocene Climatic Optimum; MECO: Middle Eocene Climatic Optimum; PETM: Paleocene-Eocene Thermal Maximum; Priabon.= Priabonian; Seland.= Selandian; Thanet.= Thanetian.
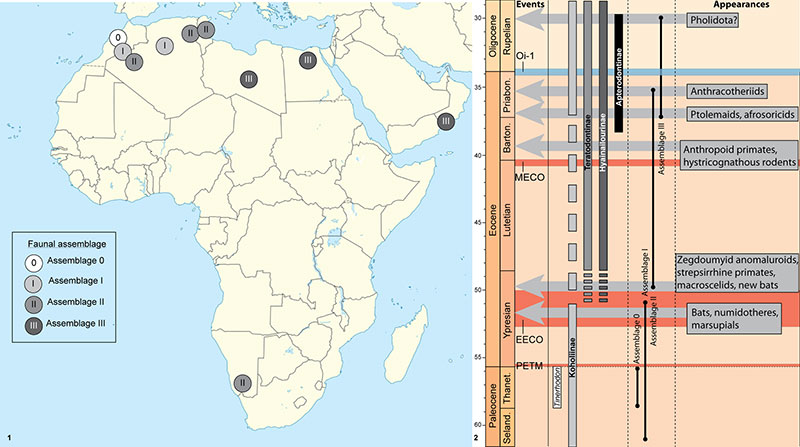
TABLE 1. Distribution of the hyaenodonts presently known in the Paleocene and Eocene of Africa. Ouled Abdoun Basin: Gheerbrant et al. (2006) and Solé et al. (2009); Ouarzazate Basin: Gheerbrant (1995; et al., 2006); El Kohol: Crochet (1988); Gour Lazib area: Solé et al. (2014b); Black Crow locality from Sperrgebiet: Pickford et al. (2008); Dur at-Talah: Grohé et al. (2012); Birket Qarun Locality 2: Borths et al. (2010); Jebel Qatrani and Qasr el-Sagha Formations: Simons and Gingerich (1974, 1976), Holroyd (1994, 1999), Lewis and Morlo (2010); Taqah: Crochet et al. (1990); Chambi and Bir el Ater based on the present paper. *: Taxa described in Holroyd (1994) but not formally published. First column refers to Figure 1 localities.
| Faunal Assemblage | Localities | Incertae sedis | Teratodontinae | Koholiinae | Hyainailourinae | Apterodontinae | |
| 10 | III | Taqah | - | Masrasector ligabuei | - | - | - |
| 9 | III | I and M
(Jebel Qatrani Formation) |
- | Masrasector pithecodacos* | - | Pterodon syrtos
Pterodontine A |
Apterodon macrognathus |
| III | G and V
(Jebel Qatrani Formation) |
- | Masrasector aegypticum
Metasinopa osborni |
Metapterodon schlosseri | Pterodontine C | - | |
| III | E
(Jebel Qatrani Formation) |
- | Masrasector ligabuei | - | Pterodon sp. | Apterodon macrognathus
Apterodon altidens |
|
| III | A and B
(Jebel Qatrani Formation) |
Brychotherium ephalmos*
Masrasector ligabuei |
Metapterodon schlosseri | Pterodon africanus
Pterodon phiomensis Akhnatenavus leptognathus |
Apterodon macrognathus
Apterodon altidens |
||
| III | L-41
(Jebel Qatrani Formation) |
- | Brychotherium ephalmos*
Masrasector ligabuei |
- | Akhnatenavus aff. leptognathus
Pterodontine D |
- | |
| III | Qasr el-Sagha Formation | - | - | Metapterodon sp. | - | Apterodon saghensis
Apterodon sp. |
|
| III | Birket Qarun Locality 2 | - | ?Masrasector | - | - | - | |
| 8 | III | Dur at-Talah | - | - | - | - | Apterodon langebadreae
Apterodon indet. |
| 7 | II | Bir el Ater | - | Masrasector cf. ligabuei | - | Hyainailourinae indet. | - |
| 6 | II | Sperrgebiet area | - | Teratodontinae indet. | - | Pterodon sp. | - |
| 5 | II | Chambi | - | Teratodontinae indet. | - | Furodon crocheti
Parvavorodon gheerbranti |
- |
| 4 | II | Gour Lazib area | - | Glibzegdouia tabelbalaensis | - | Furodon crocheti
Parvavorodon gheerbranti |
- |
| 3 | I | El Kohol | - | - | Koholia | - | - |
| 2 | 0 | Ouarzazate Basin | Tinerhodon disputatus
?Proviverrinae |
- | - | - | - |
| 1 | I | Ouled Abdoun Basin | - | - | Ypresian: Boualitomus | - | - |
| Selandian:
Lahimia |
TABLE 2. Measurements (in mm) of the specimens of Furodon crocheti Solé et al., 2014b from Chambi. *: fragmentary specimen.
| Locus | n | OR | |
| P2 | L | 2 | 4.5-4.82 |
| W | 2 | 1.64-1.8 | |
| M1 | L | 1 | 5.74 |
| W | 1 | 5.22 | |
| dp3 | L | 1 | 5.84 |
| W | 1 | 2.56 | |
| m1 | L | 1 | 2.86* (trigonid) |
| W | 1 | 2.32* | |
| m3 | L | 1 | 4.34* (trigonid) |
| W | 1 | 2.98* |
TABLE 3. Measurements (in mm) of the specimens of Parvavorodon gheerbranti Solé et al., 2014b from Chambi. *: fragmentary specimen; **: based on alveoli.
| Locus | n | OR | |
| P3 | L | 1 | 3.48 |
| W | 1 | 1.65 | |
| M1or2 | L | 1 | 3.55* |
| W | 1 | 3.56 | |
| dp3 | L | 1 | 3.78** |
| W | 1 | 1.4** | |
| dp4 | L | 1 | 3.8 |
| W | 1 | 1.44 | |
| p2 | L | 1 | 2.76 |
| W | 1 | 1.14 | |
| m1 | L | 1 | 4.48 |
| W | 1 | 1.86 | |
| m3 | L | 2 | 4.11-4.21 |
| W | 2 | 1.71-2.45 | |
| DD | 1 | 5.42 |
New fossils of Hyaenodonta (Mammalia) from the Eocene localities of Chambi (Tunisia) and Bir el Ater (Algeria), and the evolution of the earliest African hyaenodonts
Plain Language Abstract
For many years, palaeontologists from the Institut des Sciences de l'Evolution de Montpellier (France) focused on the origin and evolution of the African mammals during the Paleogene. In this framework, they work with geologists and palaeontologists from Tunisia (Office National des Mines) and Algeria (Université d'Oran and Université Abou Bekr Belkaid Tlemcen). Their field and collaborative works led to the discovery of numerous localities in North Africa that provided crucial data, notably for understanding the emergence and evolution of primates, rodents, bats, and afrotherians. A collaboration with F. Solé (Royal Belgian Institut of Natural Sciences; Belgium), who is interested in the evolution of the earliest carnivorous mammals, resulted in the publication of the hyaenodonts from the Gour Lazib area, an Algerian locality near the Ypresian/Lutetian Eocene boundary, in 2014.
The present study deals with the hyaenodonts from Chambi (late Ypresian or early Lutetian; Tunisia) and Bir el Ater (latest Bartonian or earliest Priabonian; Algeria). The fossil material provides new elements for poorly known Eocene hyaenodonts, which are the oldest ones for two African subfamilies: Hyainailourinae and Teratodontinae. The new fossils described also allow discussing the age of the two localities and understanding the evolution of these extinct predators in Africa.
This knowledge is important because (1) the hyaenodonts have been the only carnivorous mammals in Africa until the arrival of Carnivora (during the late Oligocene, 23 m.y.) and (2) because, above all, the geographic origin of this group is still debated. There is a global increase in the subfamilial richness amongst hyaenodonts during the Eocene. We, however, identified a modification of the hyaenodont fauna in the Maghreb after the "Early Eocene Climatic Optimum" (EECO), 53–50 m.y. This faunal change, which seems contemporaneous of a general drastic replacement in the composition of the African mammal faunas, might be related to the decrease of the global temperature after the EECO event, an important increase of the greenhouse. This faunal turnover could result either from endemic evolution or dispersals. Conversely, the African carnivorous fauna did not suffer from the abrupt cooling (Oi-1) that occurred around the Eocene-Oligocene boundary.
Resumen en Español
Nuevos fósiles de Hyaenodonta (Mammalia) de las localidades eocenas de Chambi (Túnez) y Bir el Ater (Algeria), y la evolución de los hienodóntidos africanos más tempranos
Presentamos y describimos nuevos fósiles del Eoceno de África del Norte de localidades de Chambi (Túnez, Ypresiano tardío o Lutetiano temprano) y Bir el Ater (Algeria; Bartoniano más tardío o Priaboniano más temprano). Los especímenes de Chambi permiten el reconocimiento de dos Hyainailourinae recientemente descriptos: Furodon crocheti y Parvavorodon gheerbranti; estos taxones se conocían previamente del área de Gour Lazib (Algeria; Ypresiano tardío tardío o Lutetiano temprano). El nuevo material de Túnez incluye un dentario fragmentario de Parvavorodon que apoya substancialmente el estatus de Hyainailourinae de este género y representa el fragmento de dentario más antiguo conocido para un juvenil de Hyaenodonta en África. La presencia de Furodon y Parvavorodon en Chambi fortalece el apoyo de la hipótesis de contemporaneidad de las localidades portadoras de mamíferos eocenas Gour Lazib y Chambi. Además, el hallazgo de un cuarto premolar teratodontino típico en Chambi testifica la presencia de un pequeño representante de este grupo. El registro fósil en Bir el Ater es más escaso que en Chambi. Sin embargo, reconocemos especímenes atribuíbles a un Hyainailourinae y un teratodontino. Este último es referido a Masrasector cf. ligabuei, y es el registro más antiguo de este género. Identificamos tentativamente una modificación en la fauna de hienodóntidos en el Maghreb luego del "Óptimo Climático del Eoceno Temprano" (OCET). Este cambio faunístico puede estar relacionado con el descenso de la temperatura global luego del evento OCET. Sería contemporáneo a un recambio drástico en la composición de las faunas de mamíferos en África. Finalmente, el final del Eoceno (Priaboniano) muestra un aumento en la riqueza a nivel de subfamilia dentro de los hienodóntidos.
Palabras clave: Teratodontinae; Hyainailourinae; Koholiinae; Apterodontinae; África del Norte; Paleógeno
Traducción: Diana Elizabeth Fernández
Résumé en Français
Nouveaux fossiles de Hyaenodonta (Mammalia) des localités éocènes de Chambi (Tunisie) et de Bir el Ater (Algérie), et l'évolution des plus anciens Hyaenodonta africains.
Nous présentons et décrivons de nouveaux fossiles provenant des localités éocènes nord-africaines de Chambi (Tunisie ; Yprésien récent ou Lutétien ancien) et de Bir el Ater (Algérie ; Bartonien terminal ou Priabonien basal). Les spécimens de Chambi permettent d'identifier deux Hyainailourinae récemment décrits : Furodon crocheti et Parvavorodon gheerbranti ; ces taxons étaient connus jusqu'à présent dans la zone du Gour Lazib (Algérie ; Yprésien récent ou Lutétien ancien). Le nouveau matériel de Tunisie inclut un dentaire fragmentaire de Parvavorodon qui soutient fortement l'appartenance de ce genre aux Hyainailourinae et représente le plus ancien fragment de dentaire connu jusqu'à présent pour un juvénile de Hyaenodonta en Afrique. La présence de Furodon et Parvavorodon à Chambi renforce l'hypothèse selon laquelle les localités à mammifères éocènes de Chambi et des Gour Lazib sont contemporaines. De plus, la découverte d'une quatrième prémolaire typique des Teratodontinae à Chambi témoigne de la présence d'un représentant (de petite taille) de ce groupe. Le registre fossile de Bir el Ater est moins riche que celui de Chambi. Nous reconnaissons toutefois des spécimens appartenant à un Hyainailourinae et à un Teratodontinae. Ce dernier est attribué à Masrasector cf. ligabuei, et représente la plus ancienne occurrence de ce genre. Nous identifions un possible changement dans la faune des Hyaenodonta au Maghreb après « l'Optimum Climatique de l'Éocène Ancien » (EECO, « Early Eocene Climatic Optimum »). Ce changement faunique pourrait être lié à la diminution des températures globales après l'évènement de l'EECO. Il apparaît comme étant contemporain d'un remplacement modifiant drastiquement la composition des faunes mammaliennes en Afrique. Enfin, la fin de l'Éocène (Priabonien) montre une augmentation de la richesse au niveau sous-familial chez les Hyaenodonta.
Mots-clés : Teratodontinae ; Hyainailourinae ; Koholiinae ; Apterodontinae ; Afrique du Nord ; Paléogène
Translator: Antoine Souron
Deutsche Zusammenfassung
In progress
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024