DISCUSSION
The general history of the Orthida
in the GIOR is in accordance with the general tectonic and faunal history of the
region established by earlier authors (Cocks and Torsvik
2002). Even though
endemic rates and ordination analysis targets two different aspects of the
faunal data they can be compared and support each other or conversely
indicate that the results of the analyses are uncertain.
The faunal data can be divided
into three segments based on the overall association trends of the analyses: The
1st segment includes the Early-Middle Ordovician, where the old
Gondwana marginal terranes of Baltica and later Avalonia changed faunal content
to develop a more mixed composition between the equatorial terranes and the
polar to high latitude Gondwanan terranes. At the
same time there was a dramatic diversification and geographical expansion among
the orthides (Harper and Gallagher 2001;
Harper and Mac Niocaill
2002), which
was truncated by an Abereiddian-Llandeilian diversity reduction and an increase
in endemism. This diversity reduction may have been the result of a contraction
in available living space caused by a regression during the Arenig (Barnes et
al. 1996). From the Early Ordovician, the European massifs formed their own
faunal association, with Armorica and Iberia forming a tight link, which is in
accordance with the conclusions of Cocks and Torsvik (2002); these authors
included Iberia with Armorica, except the South Portuguese Zone where the oldest
rocks are of Middle Devonian age (Cocks
2000). Note that the faunal association
of the European massifs may not be restricted to these, as the analysis
presented here does not include all the plates in the neighborhood of the
European massifs.
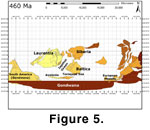 The 2nd segment
(Figure 5) was initiated during the Costonian with the singularly largest diversity
increase for most of the plate groups investigated. During the 2nd
segment, Baltica, Avalonia and Laurentia consolidated as a coherent faunal
association independent of the peri-Gondwanan European massifs and core
Gondwanan terranes, creating a gradient that correlates roughly with
palaeolatitude (Torsvik 1998;
Cocks and Torsvik
2002). Three distinct faunal
associations formed, however, note that neither analytical method presented here
provides a measure of the physical distance between the plates. The 3rd
segment (Figure 6) covers the Silurian, during which the overall faunal
association pattern changed rapidly as a result of the Late Ordovician
extinction event, forming a more or less homogenous faunal association during
the early parts of the Llandovery.
The 2nd segment
(Figure 5) was initiated during the Costonian with the singularly largest diversity
increase for most of the plate groups investigated. During the 2nd
segment, Baltica, Avalonia and Laurentia consolidated as a coherent faunal
association independent of the peri-Gondwanan European massifs and core
Gondwanan terranes, creating a gradient that correlates roughly with
palaeolatitude (Torsvik 1998;
Cocks and Torsvik
2002). Three distinct faunal
associations formed, however, note that neither analytical method presented here
provides a measure of the physical distance between the plates. The 3rd
segment (Figure 6) covers the Silurian, during which the overall faunal
association pattern changed rapidly as a result of the Late Ordovician
extinction event, forming a more or less homogenous faunal association during
the early parts of the Llandovery.
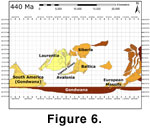 This
homogenous association gradually differentiated, however, perhaps as a result of
the core Iapetan plates moving towards Gondwana during the Silurian, the same
degree of faunal provincialism as evidenced during the Middle-Late Ordovician
was never re-established. At the Lower-Middle Silurian boundary, two distinct
faunal associations emerged from the supergroup, forming a (within the region)
cosmopolitan low-mid latitude fauna and a high latitude fauna (Clarkeia
fauna, see Fortey
and Cocks 2003). Furthermore, the apparent isolation of Siberia indicates a
separate faunal association of this plate.
This
homogenous association gradually differentiated, however, perhaps as a result of
the core Iapetan plates moving towards Gondwana during the Silurian, the same
degree of faunal provincialism as evidenced during the Middle-Late Ordovician
was never re-established. At the Lower-Middle Silurian boundary, two distinct
faunal associations emerged from the supergroup, forming a (within the region)
cosmopolitan low-mid latitude fauna and a high latitude fauna (Clarkeia
fauna, see Fortey
and Cocks 2003). Furthermore, the apparent isolation of Siberia indicates a
separate faunal association of this plate.
During this interval the European massifs retained
their faunal integrity, however, Bohemia changed from being associated with the
massifs to the larger Gondwanan-Iapetan association, indicating movement of this
terrane. These conclusions are tentative however, being hampered by very low
Silurian diversities, which add uncertainty to the results of the analyses.
Siberia
Siberia is highly mobile throughout the analyses,
which is partly the result of low diversities during the earlier and later
temporal intervals analyzed. The Siberian orthides appear to be of a mixed
association, including both Gondwanan and Laurentian elements, which causes the
spurious placement in the dendrograms (Figure
3). The relationship of Siberia with the other GIOR plates is clearer in the
NMDS diagrams (Figure 4), where it is
initially shown with a placement near Laurentia and South America, however,
during the Late Ordovician Siberia became progressively more affiliated with the
core Iapetan plates, retaining, however, a distinct faunal signal. The Late
Ordovician extinction event possibly affected the Siberian faunas differently
than the Laurentian as seen in the placement of Siberia in the Rhuddanian NMDS
diagram (Figure 4, bottom left). A
tentative link with the Silurian Gondwanan-Iapetan faunal association was,
however, retained during the entire Silurian. This pattern is consistent with
other faunal data (Nikiforova and
Andreeva 1961) and is repeated for groups like trilobites (Chugaeva
1973). These faunal data suggest that Siberia was at least in partial faunal
contact with Laurentia during the Silurian, this distance being smaller than the
distance to the North China Plate, whose influence was apparently less; during
the Early and Early-Middle Silurian the fauna remained a tropical pandemic (Fortey
and Cocks 2003).
The Precordillera
This terrane is also highly mobile in the cluster and
ordination diagrams presented here, owing both to changing compositions in the
faunal data and low diversities. Orthide data support a Laurentian origin for
the Precordillera. During the Arenig, 15 genera are reported from the
Precordillera. With the exception of Monorthis, the Precordilleran genera
were all found on Laurentia. Monorthis is reported from the Early
Ordovician only from Wales (Avalonia) and Argentina (the Precordillera). This is
in accordance with Fortey
and Cocks (2003), who noted an Avalonian influence in the faunas. Apart from
Argentina, orthides are reported from Bolivia, Brazil and Colombia within South
America. These records are unfortunately much more fragmented than those from
Argentina. There are five genera recorded from Arenig deposits of these
countries, and none of these is found on the Precordillera, however, for example
Acanthorthis and Orthidium are shared between the Precordillera
and Laurentia. This lends to support for Precordillera origin within Laurentia
as proposed by Thomas (1991); Astini
et al. (1995); Davis
et al. (1997) and Gerbi (1999).
The orthides of the Middle-Late Ordovician from
the Precordillera are commonly widespread to cosmopolitan. The orthide data,
therefore, cannot be safely used to delineate the position in time and space of
this terrane during the Late Ordovician faunal data from other groups needs
to be employed.
Bohemia (Perunica)
A greater amount of orthide data are known from
the Ordovician-Silurian of Bohemia, than from any other peri-Gondwanan terrane,
due to the studies of the Bohemian successions in the Prague Basin, especially
by authors such as Barrande in the 19th century and more recently by
Havlíček (Havlíček 1977; Havlíček
et al. 1994). During the Early-Middle Ordovician, Bohemia obviously formed
part of the European massif association, supporting the conclusions of Fortey
and Cocks 2003. As indicated by the cluster analysis, the European massifs
remain more closely related to each other than any other plate or plates during
the entire Ordovician, and only by the Middle Silurian does Bohemia switch
association (Figure 3, bottom right).
During the Caradoc, ordination plots indicate, however, that Bohemia was that of
the European massifs most closely associated with the Iapetan and Gondwanan
plates (Figure 4), a situation which
developed during the Caradoc-Ashgill.
The orthide data confirm the conclusions of Fortey
and Cocks (2003), who summarized trilobite and brachiopod data from Bohemia,
and noted that the terrane was probably close to, however, perhaps tectonically
separate from West Gondwana by the Tremadoc. Greatest oceanic separation was
achieved during the Caradoc, followed by an interval when Bohemia drifted
progressively closer to Baltica.
