DESCRIPTION AND COMPARISONS
Horn Core
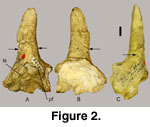 The left
horn core (52139A) is complete and contains some fragmentary adjoining frontal
bone (Figure 2). The horn core is short, nearly straight (no divergence), with a
slight posterior curvature (recurved). The horn core is medio-laterally
(transversely) compressed, with the medial side somewhat flattened and the
lateral side more convex, producing an ovate cross section. The relatively
smooth surface lacks both anterior and posterior keels and longitudinal grooves.
There is no evidence of torsion (spiral twist to horn core and horn sheath).
Sinus cavities extend into the basal portion of the horn core (Figure 2). The
cranium is broken immediately adjacent to the position of a postcornual fossa
(pf, Figure 2). If this fossa was small, it would probably be absent from the
portion of the fossil preserved. Until more complete specimens can be located,
there is no evidence that a postcornual fossa occurred on Sinocapra willdownsi
Extant Ovis, Capra, Hemitragus, and Oreamnos have a highly reduced to absent
postcornual fossa, whereas a shallow yet distinct fossa occurs on the holotype
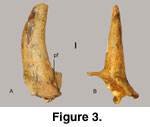
of S. minor (Figure 3;
Chen 1991).
The left
horn core (52139A) is complete and contains some fragmentary adjoining frontal
bone (Figure 2). The horn core is short, nearly straight (no divergence), with a
slight posterior curvature (recurved). The horn core is medio-laterally
(transversely) compressed, with the medial side somewhat flattened and the
lateral side more convex, producing an ovate cross section. The relatively
smooth surface lacks both anterior and posterior keels and longitudinal grooves.
There is no evidence of torsion (spiral twist to horn core and horn sheath).
Sinus cavities extend into the basal portion of the horn core (Figure 2). The
cranium is broken immediately adjacent to the position of a postcornual fossa
(pf, Figure 2). If this fossa was small, it would probably be absent from the
portion of the fossil preserved. Until more complete specimens can be located,
there is no evidence that a postcornual fossa occurred on Sinocapra willdownsi
Extant Ovis, Capra, Hemitragus, and Oreamnos have a highly reduced to absent
postcornual fossa, whereas a shallow yet distinct fossa occurs on the holotype
of S. minor (Figure 3;
Chen 1991).
 The horn core length is measured from the pedicle
base (where the burr would be located; arrows in
Figure 2; equals base of horn
sheath) to the tip. Medio-lateral and nasal-nuchal (antero-posterior)
measurements, taken at pedicle base, are included in
Table 1.
The horn core length is measured from the pedicle
base (where the burr would be located; arrows in
Figure 2; equals base of horn
sheath) to the tip. Medio-lateral and nasal-nuchal (antero-posterior)
measurements, taken at pedicle base, are included in
Table 1.
Discussion: Although the horn core of
Sinocapra willdownsi is unusually small relative to those of living caprines
and the extinct S. minor (Table 1), we interpret that the small horn
cores do not indicate a juvenile growth stage.
Figure 4 contains a series of
skulls showing ontogenetic growth stages for male and female Ovis canadensis
(Rocky Mountain bighorn) and illustrates the sexual dimorphism that is
expressed at an early age. Using even this small series of skulls, it is easy to
determine the sex and approximate age of the wild sheep.
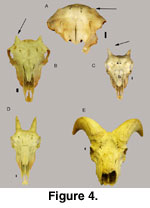 At about three months
of age, the horn core begins to appear on the frontal (Figure 4A). The horn core
develops more rapidly on males. At an early age the horn core has a wide pedicle
base yet little length (male and female); torsion begins early for males (arrow
in Figure 4C). The young female (Figure
4B) still has a wide base to the horn
core yet no torsion develops. These sexual dimorphic traits occur because the
horn core and horn sheath in male Ovis spp. is highly twisted at
maturity, providing the characteristic lateral curl to wild sheep rams (Figure
4E, Figure 5I-L).
Horn cores of female Ovis spp. are not twisted and are
more-or-less straight with a slight posterior curvature (Figure 4D). The horn
sheaths (therefore the horn cores also) of adult female Ovis spp. show
little growth after about 2.5 years age (Geist 1971). Using horn growth of
Ovis as a model for Sinocapra, it would appear that the individual
from Panaca was likely at full growth stage of development.
At about three months
of age, the horn core begins to appear on the frontal (Figure 4A). The horn core
develops more rapidly on males. At an early age the horn core has a wide pedicle
base yet little length (male and female); torsion begins early for males (arrow
in Figure 4C). The young female (Figure
4B) still has a wide base to the horn
core yet no torsion develops. These sexual dimorphic traits occur because the
horn core and horn sheath in male Ovis spp. is highly twisted at
maturity, providing the characteristic lateral curl to wild sheep rams (Figure
4E, Figure 5I-L).
Horn cores of female Ovis spp. are not twisted and are
more-or-less straight with a slight posterior curvature (Figure 4D). The horn
sheaths (therefore the horn cores also) of adult female Ovis spp. show
little growth after about 2.5 years age (Geist 1971). Using horn growth of
Ovis as a model for Sinocapra, it would appear that the individual
from Panaca was likely at full growth stage of development.
Table 1 illustrates the small size of the Panaca
caprine horn core in comparison to horn cores of other adult caprines including
Sinocapra minor (sex not designated by
Chen [1991]; see discussion
below), Ovis dalli (Dall’s bighorn; male and female), Ovis nivicola
(Snow sheep; male), and Ovis shantungensis (extinct bighorn; assumed to
be male), and the two ‘rupicaprines’ Neotragocerus (two species, extinct)
and Oreamnos (two species, one extinct; male and female). Clearly the
horn cores of the Panaca caprine are small, yet based on length and diameter
measurements, the horn core is considered to be from an adult individual (a
determination that is supported by tooth eruption; see below).
 There is a slight posterior curvature orientation
to the horn core of S. willdownsi, with no indication of torsion
(such as would be found with the Oioceros (extinct) and members of the
Bovini, Ovibovini, and many males of taxa within Caprini; e.g., Ammotragus
(aoudad), Pseudois (bharal), Ovis, and some forms of Capra
(goat); Figure 5). The surface of the horn core of S. willdownsi
is relatively smooth (as is found in Capra, Hemitragus (tahr),
Oreamnos, Ovis, and Sinocapra minor) and lacks the deep
grooves found with members of the Bovini, Ovibovini, Oioceros, and
Saiga.
There is a slight posterior curvature orientation
to the horn core of S. willdownsi, with no indication of torsion
(such as would be found with the Oioceros (extinct) and members of the
Bovini, Ovibovini, and many males of taxa within Caprini; e.g., Ammotragus
(aoudad), Pseudois (bharal), Ovis, and some forms of Capra
(goat); Figure 5). The surface of the horn core of S. willdownsi
is relatively smooth (as is found in Capra, Hemitragus (tahr),
Oreamnos, Ovis, and Sinocapra minor) and lacks the deep
grooves found with members of the Bovini, Ovibovini, Oioceros, and
Saiga.
The horn core of S. willdownsi is
medio-laterally flattened (ovate cross section), typical of most Caprini and
unlike the circular to nearly circular horn cores found within the ‘rupicaprine’
Capricornis (serow), Nemorhaedus (goral), Neotragocerus,
and Oreamnos, and also within Pachygazella (extinct), Oioceros,
Saiga, and the ovibovines. Medio-laterally compressed horn cores occur on
female caprines (e.g., Ovis and Hemitragus), perhaps because their
small horns are not used for extreme butting. In contrast, adult males of the
living Caprini use their horns for extensive butting. Their horn cores typically
have rounded lateral surfaces and anterior surfaces (contact when butting) that
range from comparatively flat (Ovis, Ammotragus, Pseudois,
and some forms of Capra) to highly ridged, forming an acute triangular
cross section (Hemitragus, most forms of Capra;
Figure 5 shows the
horn sheath expressions of the horn cores).
The horn core, type, for Sinocapra minor,
is larger than that of S. willdownsi, yet it, too, shows little posterior
curvature and lacks torsion (Figure 3). Similar horn cores were referred to
Sinocapra minor by Chen (1991; see
Teilhard de Chardin and Trassaert 1938).
They are all short, moderately straight with minor posterior curvature, lack
divergence, and have a comparatively wide pedicle base (one with a postcornual
fossa; Figure 3). Because of this wide pedicle base, we assume that the
illustrated horn cores for S. minor are from male members of a genus with
relatively small horn sheaths that are curved posteriorly and exhibit no
torsion. Because the pedicle base on the type for S. willdownsi is not
significantly larger than the burr base, as observed on S. minor horn
cores and typically found on male caprines, the Panaca caprine specimen is
considered to be a female.
Maxillary Teeth
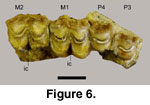 The right P3-M2
(52319 B) were recovered together in a small fragment of the maxilla (Figure 6).
The posterior edge of the M2 has a contact mark showing that there was an M3 in
the toothrow; this tooth was not recovered. The right P2 and left M1 were
recovered as isolated teeth. The molars lack entostyles. Crown height is
relatively low, mesodont. Strong labial ribs occur between the parastyle,
mesostyle, and metastyle. Distinct internal cavities occur between the protocone
and metaconule on the M1-2 (Figure 6).
The right P3-M2
(52319 B) were recovered together in a small fragment of the maxilla (Figure 6).
The posterior edge of the M2 has a contact mark showing that there was an M3 in
the toothrow; this tooth was not recovered. The right P2 and left M1 were
recovered as isolated teeth. The molars lack entostyles. Crown height is
relatively low, mesodont. Strong labial ribs occur between the parastyle,
mesostyle, and metastyle. Distinct internal cavities occur between the protocone
and metaconule on the M1-2 (Figure 6).
Discussion: The teeth do not show extreme
wear, indicating that the low crowns on the Panaca caprine are not to the result
of abrasion (Figure 6). The M2 comes into full functional wear at about two
years of age, and the M3 is in complete functional wear at about 3.0 to 3.5
years in Ovis spp. (Cowan 1940). Assuming a similar tooth eruption phase
in Sinocapra, the individual from Panaca would have been at least three
years old.
The observed occlusal characters of Sinocapra
willdownsi are consistent within Caprini but appear undiagnostic to genus.
Measurements of the molars and premolars of S. willdownsi are
compared with Ovis ammon (argali), O. canadensis, O.
catclawensis (extinct bighorn), O. dalli (female), O.
shantungensis, and O. zdanskyi (extinct bighorn) in
Table 2 (no
dentition is known for S. minor). The length and width of the
premolars of S. willdownsi fit within the variability observed
among these taxa. The size of the molars indicates some variation among genera.
These initial data imply that the teeth and toothrow of S. willdownsi
are at the small end of the observable range of living and some fossil caprines.
The mesodont characteristic may be of importance as the teeth of Sinocapra
willdownsi are distinctly lower crowned than any living Ovis spp.
or Oreamnos spp.
 Humerus
Humerus
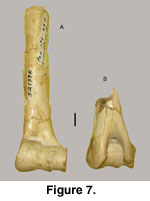
Fragmented left and right humeri (52319E) are
known (Figure 7). The left humerus preserves the distal three-fourths while the
right humerus is represented by only the distal quarter. Measurements are shown
in Table 3.
Ulna
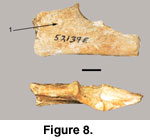
A fragment of the proximal end of the left ulna
(52319E) is recovered (Figure 8). The conical impression on the medial surface
(Figure 8.1), is consistent with a canine impaction but there is no
corresponding indentation on the lateral side.
 Lunar
Lunar
The right lunar (52319E) is complete but not
remarkably different from that of other taxa.
Metacarpal
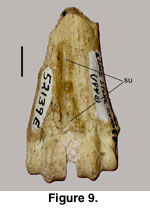
The distal fragment of a right metacarpal (52139E)
has a distinctive sulcus (su) with slight crests (not sharp) parallel to the
sulcus and is widely open (sagittal groove) at the distal epiphysis (Figure 9).
Although the entire bone is not preserved, there is enough (approximately the
distal third) preserved to indicate that the bone is from the right side and
progressively widens from the diaphysis at the distal end, as occurs on a
metacarpal and unlike the rapid expansion observed in metatarsals (see
discussion below).
 The width of the distal articular surface of the
metacarpal of Sinocapra willdownsi and other caprine taxa are included in
Table 4. The articular condyles of the first phalanges (verticilli of
Köhler
1993) are high and sharply delineated, as seen in Ovis, Oreamnos,
and other inhabitants of mountainous terrain.
The width of the distal articular surface of the
metacarpal of Sinocapra willdownsi and other caprine taxa are included in
Table 4. The articular condyles of the first phalanges (verticilli of
Köhler
1993) are high and sharply delineated, as seen in Ovis, Oreamnos,
and other inhabitants of mountainous terrain.
Discussion: Ovis spp. and
Oreamnos spp. do not have the prominent sulcus found on the metacarpal as
seen in Sinocapra willdownsi. Living Caprini, ‘Rupicaprini,’ and
Ovibovini have a superficial sulcus or none at all on the metacarpal. The
measurements of the distal articular surface of various caprines indicate that
the metacarpal of S. willdownsi is of about average size, compared to
other living caprines (Table 4). The
data presented for the metatarsal are similar.
Phalanges
The first, second, and third (fragment) phalanges
(52139E) appear to fit together and articulate precisely with the right
metacarpal fragment, and therefore, they are assumed to be medial manus
phalanges (third digit). An additional first phalanx is known but does not
adequately articulate with either the right metacarpal or the right metatarsal
and is therefore not described.
 The first phalanx measures 47.1 mm long (greatest
length between medial condyles) and 17.2 mm wide at the proximal end. The
additional first phalanx measures 49.2 mm long and 16.8 mm wide at the proximal
end. Interdigital ligament insertion ridges are short but prominent on both
specimens (Figure 10B.1).
The first phalanx measures 47.1 mm long (greatest
length between medial condyles) and 17.2 mm wide at the proximal end. The
additional first phalanx measures 49.2 mm long and 16.8 mm wide at the proximal
end. Interdigital ligament insertion ridges are short but prominent on both
specimens (Figure 10B.1).
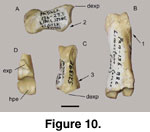
The second phalanx is short and broad (27.6 mm
long, following von den Driesch 1976), 13.9 mm wide at the proximal end. A postarticular plateau is lacking (Figure 10A.2), and the dorsal extensor
protuberance (dexp) is elongate (Figure 10C). The insertion of the interdigital
ligament is weak to almost absent (Figure 10C.3).
The third (distal) phalanx is fragmented, missing
the distal half. A robust protuberance for the extensor tendon insertion (exp)
is preserved (Figure 10D). A portion of the dorsal ridge is preserved distal to
the extensor tendon insertion and appears to be at a more horizontal angle
implying at least a slight convex appearance (typical of caprines, and not the
highly slanted and straight ridge as in cervoids). The horizontal platform of
the articular surface (hpe) is well established (Figure 10D).
Discussion: Although these phalanges are
not taxonomically diagnostic, they do indicate that the joint was designed for
tensor and flexor motion with a horizontal component that permits notable shock
absorbency (see Leinders 1979;
Köhler 1993). This implies that Sinocapra
willdownsi was more of a mountainous and rocky terrain climber and not an
animal adept at sustained, rapid locomotion on fairly level topography.
Femur
Highly fragmented left and right femora (52319E)
are known, but will not be discussed here.
Navicular-cuboid
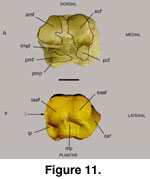 A complete right
navicular-cuboid (52139E; central and fourth tarsal) is recovered. The
comparative morphology used here is located in
Figure 11 and
Figure 12. The patterns
of the various articular facets and processes of the distal surface are within
the realm of variation observed in modern
Ovis canadensis and O. dalli,
and therefore, are of little use to distinguish the new taxon. However, the
articular facets and processes are distinctly unlike those found on cervoids and
antilocaprids (see Lawrence 1951;
Heintz 1970).
A complete right
navicular-cuboid (52139E; central and fourth tarsal) is recovered. The
comparative morphology used here is located in
Figure 11 and
Figure 12. The patterns
of the various articular facets and processes of the distal surface are within
the realm of variation observed in modern
Ovis canadensis and O. dalli,
and therefore, are of little use to distinguish the new taxon. However, the
articular facets and processes are distinctly unlike those found on cervoids and
antilocaprids (see Lawrence 1951;
Heintz 1970).
The plantar surface of 52139E contains the
calcaneal articular facet (caf), a flange with its length truncated distally by
the lateral groove of the tendon to the musculus peroneus longus (tmpl;
Figure 11,
Figure 12). This groove on the lateral surface or its truncation by the tmpl is rarely observed in Ovis, Ammotragus, Oreamnos
americanus (male and female), or the extinct Oreamnos harringtoni.
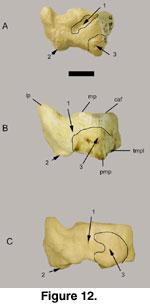 The medial process (mp;
Figure 11B,
Figure 12B) on the
anterior surface is a pronounced articular process with distinct dorsal and
plantar facets on Sinocapra willdownsi and on some specimens of
Oreamnos harringtoni (GRCA 509044, 509160, 509273). This differs from the
minute projections observed on Ovis spp. and Oreamnos americanus.
The medial process (mp;
Figure 11B,
Figure 12B) on the
anterior surface is a pronounced articular process with distinct dorsal and
plantar facets on Sinocapra willdownsi and on some specimens of
Oreamnos harringtoni (GRCA 509044, 509160, 509273). This differs from the
minute projections observed on Ovis spp. and Oreamnos americanus.
The navicular-cuboid of Sinocapra willdownsi
is square in shape as in Ovis spp., whereas it is medio-laterally wider
in Oreamnos spp. (with its wider foot for mountain climbing). This width
on Oreamnos comes from the lateral expansion of the antero-external
metatarsal facet (amf) and the postero-external metatarsal facet (pmf) along
with their adjacent ligament attachments (Figure
12A). Widening of the
Oreamnos navicular-cuboid situates the pmf near the middle of the plantar
surface, as viewed posteriorly, whereas in Sinocapra willdownsi and
Ovis spp. this process is at the lateral-plantar border of the bone (Figure 11.2). The widening within Oreamnos spp. is also caused by the medial
widening of the internal astragal articular facet, a character not found in
Sinocapra willdownsi or in Ovis spp. This character indicates that
the foot of Sinocapra willdownsi was not as wide and sturdy as that of
the mountain goats, but is comparable with the mountain sheep.
The insertion of the tendon on the plantar side
(Figure 11.1) is centrally located on Sinocapra willdownsi, yet continues
as a groove proximally. No such groove is observed in Ovis spp. or
Oreamnos spp.
The attachment area for the long medial ligament,
the plantar ligament, and the long lateral ligament is robust and extensive on
Oreamnos spp. and Ovis spp., yet is minute on Sinocapra
willdownsi (Figure 11.3). Various
muscles attach to these ligaments with tendons. Although the ligaments restrict
the amount of absolute movement at the joint, the tendons and muscles attaching
on the plantar side permit various flexor and tensor motions of the foot and
digits (see
Sisson and Grossman, 1938
and subsequent editions).
The
internal process (ip) of the internal astragal articular facet (iaaf) permits
the attachment of a number of tendons on the plantar and medial sides (Figure 11B,
Figure 12B). This process also allows for the formation of a groove that
accommodates passing the tendon of the tibialis posterior. It is in this groove
that a ligament attaches (in part the tarso-metatarsal ligament). In Ovis,
the ligament attachment blends in with the form of the groove. The anterior end
of the groove contains a large bony lump for this ligament attachment, similar
to, but not as massive as, that found on Oreamnos.
Discussion: The navicular-cuboid 52319E is similar in size and shape to that
found on Ovis and dissimilar to that of Oreamnos. Although similar
to that of Ovis, differences indicate a different animal and one that,
although an inhabitant of mountainous terrain, was not as adept to this habitat
as Ovis spp. and Oreamnos americanus are today.
Metatarsal
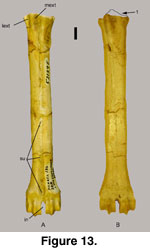 The right metatarsal (52139E) is nearly complete
(Figure 13). A sulcus (su) occurs on the dorsal surface and proceeds from
proximal to distal epiphyses, distinct most of its length, prominent and open
the distal third with crests on both sides of the sulcus, and widely open
(sagittal groove) at the distal epiphysis (Figure 13A), a character typical of bovids but not cervoids (Heintz 1970;
Leinders 1979;
Köhler 1993). This sulcus
is wide and shallow to almost non-existent on Ovis spp. and Oreamnos
spp. The goat-like caprines (Capra, Hemitragus) possess a slightly
more distinct sulcus at the distal end than do the sheep-like caprines (Ovis;
along with the ovibovines and ‘rupicaprines’). The volar or plantar surface
(posterior of Heintz 1970) has almost no trace of a groove except near the
epiphyses (Figure 13B), similar to that observed on Ovis spp. and
Oreamnos spp.
The right metatarsal (52139E) is nearly complete
(Figure 13). A sulcus (su) occurs on the dorsal surface and proceeds from
proximal to distal epiphyses, distinct most of its length, prominent and open
the distal third with crests on both sides of the sulcus, and widely open
(sagittal groove) at the distal epiphysis (Figure 13A), a character typical of bovids but not cervoids (Heintz 1970;
Leinders 1979;
Köhler 1993). This sulcus
is wide and shallow to almost non-existent on Ovis spp. and Oreamnos
spp. The goat-like caprines (Capra, Hemitragus) possess a slightly
more distinct sulcus at the distal end than do the sheep-like caprines (Ovis;
along with the ovibovines and ‘rupicaprines’). The volar or plantar surface
(posterior of Heintz 1970) has almost no trace of a groove except near the
epiphyses (Figure 13B), similar to that observed on Ovis spp. and
Oreamnos spp.
The distal end flares abruptly with a broad ‘V’
from the epiphysis to the intertrochlear notch (in) on 52139E. The connections
of the lateral extensor tendon (lext) and the medial extensor tendon (mext) are
prominent on Sinocapra willdownsi (Figure 13A). This character set is not
significant in Ovis spp. but is robust in Oreamnos spp.
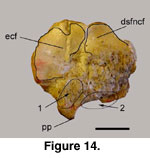 The posterior sub-facet of the navicular-cuboid
facet (psfncf) is completely abraded, and it cannot be determined if this facet
was raised or level with the rest of the naviculo-cuboid facet (Figure 13B.1
and Figure 14.2). The posteromedial projection (pp;
Figure 14) is prominent and has a
hooked-appearance on 52139E. However, this is due to abrasion and to the deep fossa below. Due to the abraded state of the fossil, this character (= character
93 of Gentry 1992) cannot be used. The articular condyles for the first
phalanges (verticilli of Köhler 1993) are high and sharply delineated, as found
with Ovis, Oreamnos, and other mountainous terrain inhabitants
(Figure 13).
The posterior sub-facet of the navicular-cuboid
facet (psfncf) is completely abraded, and it cannot be determined if this facet
was raised or level with the rest of the naviculo-cuboid facet (Figure 13B.1
and Figure 14.2). The posteromedial projection (pp;
Figure 14) is prominent and has a
hooked-appearance on 52139E. However, this is due to abrasion and to the deep fossa below. Due to the abraded state of the fossil, this character (= character
93 of Gentry 1992) cannot be used. The articular condyles for the first
phalanges (verticilli of Köhler 1993) are high and sharply delineated, as found
with Ovis, Oreamnos, and other mountainous terrain inhabitants
(Figure 13).
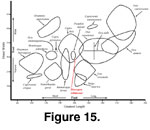 The metatarsal is neither excessively long nor
short, nor narrow or wide, when compared with other caprines (Table 5). To
observe its size relative to other caprines, we made a comparison between the
overall length and the width of metatarsals, at the distal end.
Figure 15
illustrates this comparison among the various caprines. The length/distal width
ratio of Sinocapra willdownsi is analogous to those of some Capra ibex
and Pseudois nayaur, and not all that dissimilar with some Ammotragus
lervia and Ovis orientalis. These similarities, although not helpful
with the identification of the specimen, do indicate that S.
willdownsi had a relatively short and wide foot, one very capable of
mountain terrain. The size of the metatarsal on S. willdownsi
indicates that the individual described here was not a large or heavy animal.
The metatarsal is neither excessively long nor
short, nor narrow or wide, when compared with other caprines (Table 5). To
observe its size relative to other caprines, we made a comparison between the
overall length and the width of metatarsals, at the distal end.
Figure 15
illustrates this comparison among the various caprines. The length/distal width
ratio of Sinocapra willdownsi is analogous to those of some Capra ibex
and Pseudois nayaur, and not all that dissimilar with some Ammotragus
lervia and Ovis orientalis. These similarities, although not helpful
with the identification of the specimen, do indicate that S.
willdownsi had a relatively short and wide foot, one very capable of
mountain terrain. The size of the metatarsal on S. willdownsi
indicates that the individual described here was not a large or heavy animal.
