|
|
CLADISTIC ANALYSISA parsimony analysis was performed using PAUP* 4.0 beta 10 using a modified version of Wilson's (2002) character matrix. A total of 32 sauropod genera and two outgroup taxa were scored for 230 morphological characters (Appendix 3, Appendix 4). Sauropod genera containing multiple species were scored based on a combined knowledge of the morphology of all the species. Five Early Cretaceous taxa included in this analysis were not among the 27 original taxa used in Wilson's analysis. These include Pleurocoelus, Paluxysaurus, Cedarosaurus, and Venenosaurus from North America and Phuwiangosaurus from Asia. Some modifications were made to Wilson's (2002) original character matrix for this analysis. The number of sacral vertebrae contributing to the acetabulum was omitted as a character from this analysis, because titanosaurians exhibit a different condition from other sauropods for this character that was not described in Wilson's (2002) character list. Therefore, new taxa added in the present analysis could not be evaluated for this state. An additional derived character state was added to describe the orientation of mid-caudal neural spines in sauropods (character 132, Appendix 3), to account for the condition observed in Cedarosaurus and Venenosaurus, changing it from a binary character to multistate. The multistate coding Wilson (2002) used to describe the shape of the distal scapular blade cannot account for the variation in expansion of the distal scapula that exists in sauropods. Wilson introduced two derived morphological states for this character, of which one is unique to rebbachisaurids. Camarasaurus, Jobaria, and Brachiosaurus share the other derived state, a rounded expansion on the acromial side of the distal scapula. Applying the character state descriptions explicitly as they were originally written would require all other sauropod taxa that do not exhibit either derived state to be coded as primitive, which supposes that the distal scapular blade is not expanded on the acromial side. Clearly this is not the case for taxa such as Paluxysaurus and Haplocanthasaurus, in which the distal scapula is expanded (but not rounded) on the acromial side. The first derived character state was re-worded to reflect a more general coding of presence or absence of an expansion on the acromial side of the distal scapula (Appendix 3). The derived state describing distal radial breadth, approximately twice mid-shaft breadth, was found more widely distributed than among the titanosaurians, contra Wilson's (2002) original analysis. Brachiosaurus and Paluxysaurus also show the derived condition, and among titanosaurians, some possess a ratio much greater than 200% (e.g., Alamosaurus) and others less than 200% (e.g., Malawisaurus). Malawisaurus was coded as primitive by Wilson for this character – distal breadth slightly larger than mid-shaft breadth – but the distal transverse breadth of the radius in this taxon is at least one and one-half times the mid-shaft breadth and is perhaps as much as 80% greater. The proximal and distal ends of the radius are clearly more expanded in the titanosaurians Opisthocoelicaudia (Borsuk-Bialynicka 1977, text-figure 8, plate 9) and Alamosaurus (Gilmore 1946, figure 9) than in Brachiosaurus and Paluxysaurus. But as noted above, the radius in Opisthocoelicaudia and Alamosaurus is also broader at mid-shaft than in the latter two taxa, which results in the similar ratios. Comparing other forelimb measurement ratios confirms that the radius of Alamosaurus is more robust than that of Brachiosaurus and Paluxysaurus (Appendix 1), and the ratio of minimum shaft circumference to length in the radius of Opisthocoelicaudia as reported by Borsuk-Bialynicka (1977, p. 46) sharply contrasts from the same ratio in Paluxysaurus and, in fact, exceeds that reported for any other sauropod with the exception of Saltasaurus. Cedarosaurus and Venenosaurus would both be primitive for this character in Wilson's (2002) study. In the case of the latter two taxa this is a function of a reduction in the breadth of the distal radius. Based on these observations, a character comparing distal and mid-shaft transverse breadths of the radius was not included in the present phylogenetic analysis. The shape of the preacetabular process of the ilium was not used as a character in this analysis, because primitive and derived states could not confidently be differentiated for some taxa. Personal observations of Brachiosaurus at the Field Museum in Chicago (FMNH 25107) and measurements reported in the literature indicate that the transverse breadth of the femur at mid-shaft ranges from approximately 1.6 to 2.0 times the craniocaudal width for that taxon, which suggests to me that Brachiosaurus deserves a different derived coding from that which it received in Wilson's (2002) analysis (character 195, Appendix 4). The derived state of the character describing the transverse breadth of the distal fibular condyle relative to mid-shaft breadth has a homoplasious distribution in Wilson's (2002) analysis. Several taxa that were coded by Wilson as primitive for this character appear, in fact, to be closer in morphology to the derived condition (Shunosaurus, Apatosaurus, Camarasaurus, Malawisaurus, and Opisthocoelicaudia). Paluxysaurus and Phuwiangosaurus also appear to exhibit the derived condition. Re-scoring taxa, however, did not improve the informativeness of the character. Therefore, I regard this character as phylogenetically uninformative, and it was excluded from the final character-taxon matrix. For an explanation of all other characters and character coding strategies, see Wilson (2002) and references therein. Results
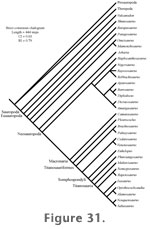
A strict consensus of cladograms one step longer than the most parsimonious ones (101 topologies) indicates a significant loss in resolution among titanosauriform clades (Figure 33). In addition to large amounts of missing data, much of the instability in relationships among titanosauriforms can be attributed to a few characters where large amounts of homoplasy exist. For instance, the character describing the shape of cranial caudal neural spines requires three independent losses or four independent acquisitions of the derived state to explain its distribution among terminal taxa.
Two characters in the phylogenetic analysis describe the number of presacral vertebrae in sauropods. The amount of homoplasy in the character state distributions may, in part, be a function of missing data in the character matrix as a whole. More likely it indicates that these characters are also phylogenetically uninformative; deleting these characters from the analysis has no effect on the topology of the most parsimonious cladograms. The new taxon from North America described here, Paluxysaurus, is hypothesized to be the sister taxon of Brachiosaurus but closely related to the three other North American Early Cretaceous sauropods included in the analysis (Cedarosaurus, Venenosaurus, and Pleurocoelus). Brachiosaurus plus the four North American Early Cretaceous taxa are all basal to Somphospondyli. The phylogenetic position of Paluxysaurus is not robustly supported. Only a single synapomorphy–relatively transversely broad femur at mid-shaft–supports the sister relationship between Paluxysaurus and Brachiosaurus. Wilson (2002) defines the group Titanosauriformes by eight derived characters shared by, "Brachiosaurus, Saltasaurus, their common ancestor, and all of its descendants" (Wilson and Sereno 1998, p. 51). Paluxysaurus is clearly a member of Titanosauriformes. It possesses most of the titanosauriform synapomorphies. It shares none of the derived characters that are currently used to define the groups Somphospondyli or Titanosauria. Within Titanosauriformes, Paluxysaurus is a basal member of the group. The Arundel sauropod also shares many of the titanosauriform synapomorphies but is here resolved as basal to both Brachiosaurus and Paluxysaurus due to primitive aspects in the morphology of the femur and the internal pneumatic structure of the presacral vertebrae. Bootstrap Analysis
|
|