|
|
|
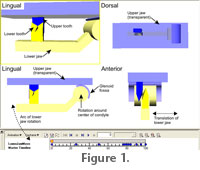
METHODSEight carnivoran species were chosen to include a range of dental and jaw joint morphologies. They fall into four basic morphologies: Group 1) upper – well-developed carnassial with a very small first molar; lower – only a well-developed carnassial (Acinonyx jubatus and Crocuta crocuta); Group 2) upper – scimitar-shaped carnassial and a single molar; lower – carnassial and a single posterior molar (Mustela lutreola and Vormela peregusna); Group 3) both upper and lower have a carnassial with two posterior molars (Canis aureus and Alopex lagopus); and Group 4) upper – carnassial and two molars; lower – carnassial and a single posterior molar (Genetta genetta and Herpestes ichneumon). Carnassial and post-carnassial upper and lower tooth rows of one side were scanned using a Nextec Hawk 3D laser scanner (Evans et al. 2007). These scans are viewable in the MorphoBrowser database, an online dental morphology database. Left and right condyle and post-glenoid surfaces and incisor positions were also scanned, maintaining correct relative position and orientation between all features. Tooth surfaces were exported as text point files, interpolated using GIS software (Surfer for Windows v. 8.0, Golden Software, Colorado, USA), and then exported as VRML (Virtual Reality Modelling Language) files. Scans were exported as STL files and converted to VRML using Rhino for Windows v. 3.0 (McNeel, Washington, USA). Upper and lower teeth and joint surfaces were aligned in CosmoWorlds for Windows v. 2.0 (Silicon Graphics, Inc., California, USA), and CosmoWorlds was used for the jaw rotation and translation movements in the occlusal reconstructions.
Two main measurements of the jaw movement were taken: the angle between initial tooth contact and centric occlusion, and the lateral movement of the jaw between initial tooth contact and centric occlusion. The second of these was standardised by the distance between the outer edges of the two glenoid fossae, termed the outer glenoid fossae distance. The shape and curvature of the large attrition facets of Acinonyx and Crocuta were examined by comparing them to a plane in 3D space. Species in Group 1 were examined as they have the largest attrition facets and are the easiest to examine for this part of the study, but this means that the results may not be applicable to the other groups. |
|