OTHER MUSCLES OF THE HEAD
This section describes the remaining muscles of the head that are not innervated by the trigeminal nerve (cn5). Most of these muscles are instead innervated by the facial (cn7), glossopharyngeal (cn9), or hypoglossal (cn12) nerves. They include the superficial muscles of the neck and throat in addition to the depressor mandibulae. In general, these muscles have received less attention than those innervated by the trigeminal nerve.
Aspects of the throat musculature have been described repeatedly (Ruge 1897;
Osawa 1898;
Edgeworth 1907,
1935;
Byerly 1925;
von Wettstein 1931,
1932;
Lubosch 1933;
Lightroller 1939;
Haas 1973;
Gorniak et al. 1982;
Al-Hassawi 2004,
2007), but most notably by
Rieppel (1978) who dissected two specimens.
Schwenk (1986) also described some of the throat musculature within a detailed description of the tongue, based on histological sections taken from one adult specimen. This largely agreed with the findings and homologies of
Rieppel (1978) but also contributes new information.
Depressor Mandibulae (mDM)
The m. Depressor Mandibulae is perhaps the most noteworthy of the muscles innervated by the facial nerve (cn7) because of its obvious function in controlling jaw opening (Gorniak et al. 1982).
Günther (1867) considered this muscle to be integral to the shearing jaw movement, but this was not supported by the detailed findings of
Gorniak et al. (1982). Despite its prominence in Sphenodon, descriptive accounts of the m. Depressor Mandibulae are often brief (e.g.,
Osawa 1898;
Ruge 1896;
Lubosch 1933;
Edgeworth 1935;
Anderson 1936;
Haas 1973) or absent (e.g., Lightroller 1939;
Wu 2003), although more detailed descriptions are provided by
Byerly (1925),
Gorniak et al. (1982),
and Al-Hassawi (2004,
2007).
Throckmorton (1978) found that the muscle remained active in the squamate Uromastyx after the jaws had started to close, perhaps to control the position of the quadrate during static biting (Herrel et al. 1998), to modify bite force, or control the speed of jaw closure. No evidence of similar activity was found in Sphenodon (Gorniak
et al. 1982).
The m. Depressor Mandibulae originates from the posterodorsal edge of the parietal and squamosal, and from a small mid-line portion of connective tissue (Figure
21, Figure 22,
Figure 28,
Figure 29,
Figure 31, and
Figure 33) (Byerly 1925;
Gorniak et al. 1982;
Al-Hassawi 2004,
2007).
Gorniak et al. (1982)
described the dorsal third as being thin, the middle third as being thick, and
the ventral third as tapering towards the attachment on the lower jaw (Figure
21, Figure 22,
Figure 23,
Figure 25,
Figure 29,
Figure 31,
Figure 34, and
Figure 35). This is confirmed by examination of specimens BMNH 1969.2204 and BMNH 1972.1.22.3. Specimen BMNH 1969.2204 shows that most of the origin is from the squamosal,
with the area of origin decreasing medially.
Al-Hassawi (2004,
2007) reported the presence of ridges and pits on the bony sites of origin but did not cite a specific specimen. Examination of Sphenodon skeletal material does not fully confirm this observation (e.g., LDUCZ x036, LDUCZ
x1176). The left squamosal of LDUCZ x036 is roughened along its dorsal edge but the right squamosal is not.
Both Edgeworth (1935) and
Haas (1973) described the m. Depressor Mandibulae as undivided, and
Abdala and Moro (2003) coded it as such in their cladistic analysis but others have reported that there is evidence of subdivision (e.g.,
Gorniak et al. 1982;
Al-Hassawi 2004,
2007; specimens BMNH 1969.2204, and BMNH 1972.1.22.3). The muscle consists of a thin portion with a more medial origin, m. Depressor Mandibulae Medialis (mDMM, posterior of
Gorniak et al.
[1982] and externus of
Al-Hassawi [2004,
2007]) and a larger fleshy portion with a more lateral origin, m. Depressor Mandibulae Lateralis (mDML, anterior of
Gorniak et al.
[1982]; internus of
Al-Hassawi [2004,
2007]). There is no obvious connective tissue division (Gorniak et al.
1982) but there are certainly differences in texture and colour. Previous authors have described the division being most obvious at the point of insertion on the lower jaw (Gorniak et al. 1982;
Al-Hassawi 2004,
2007). Fibres from the m. Depressor Mandibulae Medialis may be up to three times longer than those of the m. Depressor Mandibulae Lateralis (Table 2) (Gorniak
et al. 1982).
According to
Gorniak et al. (1982), the fibres from the m. Depressor Mandibulae Medialis curve around the end of the lower jaw and insert on its ventral surface after curving around the fibres of the m. Depressor Mandibulae Lateralis, which attach to the posterolateral end of the lower jaw. In both cases the more superficial the fibres, the further forward on the lower jaw they attach (Gorniak
et al. 1982). Al-Hassawi (2004,
2007) also describes a double insertion but one that is more localised and restricted to the posterodorsal end of the lower jaw. Examination of specimen BMNH 1969.2204 and BMNH
1972.1223 corresponds more closely to the description of
Al-Hassawi (2004,
2007), and demonstrates that the fibres of both portions converge into a short tendon, which inserts onto a very small area on the posterolateral tip of the lower jaw (retroarticular process) (Figure 35).
Al-Hassawi (2004,
2007) suggested that a depression on the dorsal surface of the retroarticular process was related to the insertion of the m. Depressor Mandibulae, but here we describe this as an attachment point for the middle part of the m. Pterygoideus Typicus. In some individuals a bony tubercle is present near this position (DGPC2,
Figure 36), but again this is perhaps more likely to be associated with the m. Pterygoideus Typicus.
Al-Hassawi (2004,
2007) describes a muscle, referred to as the "Cervicomandibularis (Cm)," originating from the dorsal crest of fatty tissue and muscle fascia, overlapping the neural spines of the atlas and axis, and inserting on the retroarticular process. As labelled
in her illustration (2004, p. 116; 2007, p. 116)
it suggests this is represents a posterior portion of m. Depressor Mandibulae (e.g. see
Haas 1973).
Constrictor Colli (mCCol)
The m. Constrictor Colli ("m. Sphincter Colli" in
Poglayen-Neuwall
1953 and Haas 1973) runs around the anterior and mid-neck regions (Figure 39) and attaches to the muscle fascia of the posterior portions of the m. Depressor Mandibulae (Versluys 1898;
Fürbringer 1900;
Lubosch 1933;
Poglayen-Neuwall
1953; Haas 1973;
Al-Hassawi 2004,
2007). Anteriorly this muscle in adults is continuous with the m. Intermandibularis (Rieppel 1978). The anterior portion of the m. Constrictor Colli described by
Lubosch (1933) is therefore probably the posterior part of the m. Intermandibularis although this is questioned by
Haas (1973) because, as drawn by
Lubosch (1933), it is too superficial. It is innervated by the facial nerve (cn7) (Haas
1973).
Verbindungsbündel (mVer)
This strip of muscle connects the posterior portion of m. Depressor Mandibulae to the anteroventral part of the m. Constrictor Colli. Correspondingly it is innervated by the facial nerve (cn7). It was reported by both
Lubosch (1933, p. 599) and
Haas (1973) and also appears to have been figured by
Lightroller (1939, figure19).
Intermandibularis (mInm)
This muscle is innervated by both the trigeminal nerve (cn5) and the facial nerve (cn7) and is therefore described in the previous section (Poglayen-Neuwall 1953;
Haas 1973).
Stylohyoideus (mSty)
The m. Stylohyoideus is inervated by the hyomandibular ramus of cranial nerve 7, the facial nerve (Rieppel
1978). This muscle originates from the distal part of the ceratohyal, and after extending posteriorly it inserts either onto the base of the ceratobranchial (Byerly
1925), or merges with the ventral part of the m. Constrictor Colli (Ruge 1896;
Rieppel 1978) (Figure 39).
Ceratohyoideus (mCeh)
This muscle is innervated by the glossopharyngeal nerve (cn9) (Rieppel
1978) contra Byerly (1925). It is located between the m. Pterygoideus laterally and m.Hyoglossus medially running between the anterior edge of the distal part of the first ceratobranchial and the posterior edge of the ceratohyal (Figure 39) (Rieppel
1978).
Geniohyoideus (mGhy)
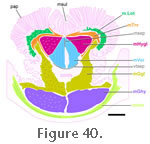 In general the m. Geniohyoideus (mGen) is a longitudinal muscle found between the lower jaw, first ceratobranchial and entoglossal process (Byerly 1925;
Edgeworth 1935;
Rieppel 1978;
Schwenk 1986). It lies deep to the m. Intermandibularis but superficial to the m. genioglossus (Figure 39 and
Figure 40). As
Schwenk (1986) described, it does not contribute to the tongue but probably plays a role in supporting it. Its muscle fibres are large in cross-sectional area, a fact that helps distinguish it from neighbouring muscles in the section (Schwenk 1986). In general the m. Geniohyoideus (mGen) is a longitudinal muscle found between the lower jaw, first ceratobranchial and entoglossal process (Byerly 1925;
Edgeworth 1935;
Rieppel 1978;
Schwenk 1986). It lies deep to the m. Intermandibularis but superficial to the m. genioglossus (Figure 39 and
Figure 40). As
Schwenk (1986) described, it does not contribute to the tongue but probably plays a role in supporting it. Its muscle fibres are large in cross-sectional area, a fact that helps distinguish it from neighbouring muscles in the section (Schwenk 1986).
According to (Rieppel 1978) it comprises two parts: a posteromedial m. Geniohyoideus 1 (mGhy 1) and a more lateral m. Geniohyoideus 2 (mGhy 2). There seems to be some uncertainty regarding the exact origins of the different parts (Figure 39), and
Schwenk (1986) could not distinguish the two parts at all.
For the squamate Ctenosaura,
Oelrich (1956) described the innervation of this muscle (="m. mandibulohyoideus I and II") as being from the hypoglossal nerve (cn12).
Geniohyoideus 1 (mGhy1). The m. Geniohyoideus 1 orginates from the lateral surface of the entoglossal process and then extends posterolaterally to insert onto the medial region of the ceratobranchial and the anterolateral edge of the basihyal (Figure 39) (Rieppel
1978). Rieppel (1978) also describes this part as originating from "the midline raphe between the symphysis and the tip of the entoglossal process"; however, in his illustration, this region of the m. Geniohyoideus is labelled as belonging to part 2 (Figure 39).
Geniohyoideus 2 (mGhy2). The m. Geniohyoideus 2 originates from the ventromedial surface of the anterior region of the dentary (Figure 39) (Rieppel
1978). As mentioned above this part may also include the fibres that originate from the midline raphe between the symphysis and the tip of the entoglossal process (Rieppel
1978, figure1). Attachment occurs on the anterior edge of the central portion of the first certatobranchial.
Hyoglossus (mHygl)
This muscle has been described many times (Byerly 1925; Lubosch 1933;
von Wettstein 1931, 1932; Edgeworth 1935;
Rieppel 1978;
Schwenk 1986). It is innervated by the hypoglossal nerve (cn12) and originates from the central part of the anterodorsal surface of the first ceratobranchial (Figure 39,
Figure 40) (Rieppel 1978). From there it travels anteriorly into the body of the tongue just posterior to the site of the transverse tendinous band (Schwenk 1986). It is possible that some fibres may end ventrally at the band itself (Rieppel 1978) but otherwise this muscle is a continuous column. Coronal histological sections demonstrate that the anterior part
of the m. Hyoglossus becomes an important component of the tongue situated dorsolaterally to the m. Verticalis and ventral to the m. Transversalis for most of the tongue's length (Figure 39 and
Figure 40) (Schwenk 1986). In ventral view the hyomandibular artery crosses the muscle from its posterolateral corner towards its anteromedial extent (Figure 33). Contraction of the m. Hyoglossus probably assists retraction of the tongue (Schwenk 1986).
Genioglossus (mGgl)
Again this muscle has been examined repeatedly (Byerly 1925;
Lubosch 1933;
Edgeworth 1935;
von Wettstein 1931,
1932;
Rieppel 1978;
Schwenk 1986). It lies deep to the m. Geniohyoideus (Byerly
1925; Lubosch 1933) and is innervated by the hypoglossal nerve (cn12) (Rieppel 1978). It runs from the anterior end of the throat towards the transverse tendon before contributing significantly to the ventrolateral body of the tongue (Rieppel 1978;
Schwenk 1986).
Rieppel (1978) described the origin of the m. Genioglossus as comprising two separate but adjacent anterior heads on the anteromedial surfaces of the lower jaws (Figure 39). The most medial head arises from the midline raphe whereas the lateral
head arises from the medial surface of the dentary near the jaw symphysis.
Schwenk (1986) observed only a single origin but suggested the difference might be explained by intraspecific variation.
Once within the tongue, muscle fibres of the m. Genioglossus can be observed to travel posteriorly, dorsally, and anteriorly (Schwenk 1986, pers. comm. 2009) both ventral and lateral to the m. Hyoglossus (Figure 40) (Schwenk 1986). Anteriorly fibres from the m. Genioglossus, in conjuction with connective tissue, form the frenulum that attaches the tongue to the floor of the mouth and dorsally some of the fibres mingle with those of the longitudinalis and transversalis (Schwenk
1986).
According to Schwenk (1986,
2000; pers. comm. 2009) contraction of the posterior part of the m. Genioglossus results in tongue protrusion whereas contraction of the anterior part contributes to tongue retraction.
Episternohyoid Complex (mEshC)
This complex consists of muscles that insert on the posterior surface of the hyoid (Rieppel 1978). For the squamate Ctenosaura
Oelrich (1956) described these muscles as being innervated by the first spinal nerve.
Omohyoideus (mOmHy).
In general the m. Omohyoideus runs parasagittally along the midline (Figure 39) (Rieppel
1978). Originating from the scapulocoracoid ligament and the anteromedial portion of the scapula (Byerly 1925), it inserts into the ventral surface of the posterior part of the basihyal and into the posteroventral edge of the proximal part of the first ceratobranchial (Rieppel
1978).
Sternohyoideus (mSteHy). The m. Sternohyoideus (m. Cleidoepisterno Hyoideus in
Byerly [1925], m. Episterno-Hyoideus
Osawa [1898],
Rieppel [1978]) runs deep and obliquely to the m. Omohyoideus so that its anterior part is more laterally positioned (Figure 39) (Rieppel 1978;). After originating from the episternum and anterior surface of the medial portion of the clavicle (Byerly 1925) it inserts into the posterodorsal surface of the distal part of the first ceratobranchial (Osawa 1898;
Rieppel 1978).
Intrinsic Muscles of the Tongue
The muscles within the tongue have been described primarily by
Schwenk (1986) who identified three distinct units: m. Verticalis, m. Transversalis, and m Longitudinalis (Figure 40). All parts are innervated by the hypoglossal nerve (cn12) (Oelrich 1956).
 The tongue itself is fleshy with a shallow midline depression or sulcus (Figure 41.1) (Günther 1867;
Gorniak et al. 1982;
Schwenk 1986) and is covered in long filamentous papillae, some of which (termed gustatory papillae,
Schwenk 1986) have expanded heads and bear taste buds (Figure 40;
Schwenk 1986). Posteriorly it has two "limbs" positioned either side of the glottis and larynx. It is connected to the base of the mouth for almost all of its length (Figure 41.2). Connective tissue forms a significant part of the tongue's structure, and this must have an important bearing on flexibility and movement (Schwenk 1986). Note that the tip is not forked or bifurcated (Schwenk 1986,
1988 contra
Gauthier et al. 1988), a condition that supposedly represents the plesiomorphic state for lepidosaurs (Schwenk 1986,
1988). The tongue itself is fleshy with a shallow midline depression or sulcus (Figure 41.1) (Günther 1867;
Gorniak et al. 1982;
Schwenk 1986) and is covered in long filamentous papillae, some of which (termed gustatory papillae,
Schwenk 1986) have expanded heads and bear taste buds (Figure 40;
Schwenk 1986). Posteriorly it has two "limbs" positioned either side of the glottis and larynx. It is connected to the base of the mouth for almost all of its length (Figure 41.2). Connective tissue forms a significant part of the tongue's structure, and this must have an important bearing on flexibility and movement (Schwenk 1986). Note that the tip is not forked or bifurcated (Schwenk 1986,
1988 contra
Gauthier et al. 1988), a condition that supposedly represents the plesiomorphic state for lepidosaurs (Schwenk 1986,
1988).
During protrusion the tongue tip curves ventrally so that the dorsal surface of the foretongue, with its long papillae, is directed toward potential prey items (Schwenk 1986,
2000).
Verticalis (mVer). The m. Verticalis is a paired muscle that runs along the central core of the tongue (Figure
40). Anteriorly the left and right portions are generally separated by the median (sagittal) septum but posteriorly the laryngohyoid ligament and entoglossal process also lie between the two portions (Schwenk 1986). Dorsolaterally the m. Verticalis is bounded by the m. Hyoglossus. The anterior portion of the m. Verticalis lies above the m. Genioglossus whereas the posterior portion largely overlies the m. Geniohyoid as the m. Genioglossus become more laterally situated.
Osawa (1897) refers to this muscle as the m. Basihyalis Proprius.
Transversalis (mTrv). The m. Transversalis extends along the entire length of the tongue. In general it lies near the dorsal surface, above the m. Hyoglossus, but part of it extends ventrally against the lateral surface of the m.Hyoglossus bundle to insert into the ventral transverse septum (Figure
40). A third part extends ventrolaterally into and around the lateral side of the m. Genioglossus bundle. As a result the muscle can look triradiate in coronal section (Schwenk 1986).
Longitudinalis (mLot). This m. Longitudinalis (mLot) is a poorly defined set of muscle fibres that runs along the lateral edge of the tongue just beneath the layer of papillae, intermingled with connective tissue (Figure
40). The fibres are most numerous and prominent in the central portion of the tongue, being largely absent from the tip and posterior limbs (Schwenk 1986).
|