|
|
|
ResultsSedimentological Data
Depositional unit A (cross-laminated sand). This depositional unit is characterized by cross-laminated fine to very fine sandstones (Figure 3,
Figure 4.1-4.2,
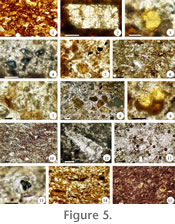
Table 1). Commonly, these sandstones are bounded by finer-grained horizons, from which they are separated by sharp to hummocky contacts; there is one example where an A sandstone is underlain by a massive sandstone (Figure 3, CH-89, unit 8). Some A units can be traced stratigraphically between sections (Figure 3, CH-79, unit 17 and CH-93, unit 18). The A sandstones are poorly sorted and contain ferruginized horizons and intraformational clasts. Other less important macrofeatures include small rooting structures, root halos, and rare mud drapes, which include concentrations of fossil organic matter. Petrographic analysis of these sandstones documents abundant concentrically-arranged iron-oxide nodules in a matrix whose main micro-feature is partial to complete coating by reddish iron-rich cement, whereas interstitial spaces are sometimes filled with chalcedony cement (Figure 5.1).
Interpretation of A depositional units. Based upon the presence of cross laminations and intraformational clasts, A sandy depositional units represent fluvial deposits of low to medium flow speed (Miall 1996). In particular, based on the fact that these sandstones are commonly adjacent (in a vertical sense) to finer-grained horizons, their deposition implies an ephemeral, energetic discharge of sediments, and, in this sense, they may represent levee/crevasse deposits typical of fluvial environments (Miall 1996; Boggs 2006). Such a change in the intensity of sedimentary discharge with respect to vertically adjacent depositional units, and therefore the origin of these types of deposits, can be the result of changes in slope, sediment size, and/or the presence of easily erodable bank materials (Mader 1985). Some A depositional units can be traced stratigraphically between sections, which is consistent with their being sheet-like deposits formed during flooding of the overbank. The abundance of macro- and micro-scopic accumulation of iron indicates well-oxygenated depositional conditions, which is consistent with the sudden discharge of these fluvial deposits (Miall 1996; Boggs 2006). The presence of rooting structures and root halos indicates subaerial exposure and plant colonization of some of these sandstones. A volcanic origin for these sediments is inferred from the abundance of volcanic rock fragments and feldspars, hornblende, and pyroxene. Smectite and kaolinite in the A depositional unit probably originated from rapid weathering of available local or regional volcanic source rocks, and their co-occurrence in these strata is consistent with variable (mild to moderate) weathering and diagenetic intensity and duration regulated by the availability of water in the environment (Bohor and Triplehorn 1993; Moore and Reynolds 1997). In particular, kaolinite indicates better drained, more acidic, more extensively weathered, and more diagenetically altered soils than smectite (Bohor and Triplehorn 1993; Moore and Reynolds 1997). Kaolinite may have also formed in situ by acidification of the soil through plant material decomposition (Khademi and Arocena 2008; Bohor and Triplehorn 1993). Depositional unit B (scour-fill silt and sand). This depositional unit includes siltstones and very fine sandstones that show scour and fill structures and also may contain well- to poorly-developed trough cross-laminations (Figure 3, Figure 4.1-4.2, Table 1). Typically, the lower limits of B depositional units scour the upper boundary of the preceding (underlying) unit or have a flat boundary, while laterally some units grade into A depositional units (e.g., Figure 3, CH-89, unit 18 and CH-93, unit 18). Towards the upper contact of some B depositional units, lenses of ashy material and small-scale channel-like structures that grade into the succeeding unit are present. B depositional units are poorly sorted and contain intraformational clasts, spherical iron oxide accumulations, clay- and organic-rich layers, and other organic remains, including poorly preserved leaves and twigs usually concentrated at the base of the unit. Petrographic data document a massive to finely-laminated silt- and organic-rich matrix that includes abundant volcanic grains, such as elongated glass shards and other angular fragments (Figure 5.6). These units also include bentonite bubbles, angular to subangular feldspar crystals, which are sometimes coated by iron-rich cement, and microscopic intraformational clasts (Figure 5.7-5.8). Other less abundant microfeatures are small pyroxene crystals, opaque particles, and concentrically arranged iron oxide accumulations and halos, which sometimes are rimmed by chalcedony cement (Figure 5.9). XRD analysis of the sediments indicates the presence of smectite, illite-smectite, kaolinite-smectite, and albite (Appendix 2, Appendix 3). Interpretation of B depositional units. Based on the features observed, B depositional units are interpreted as small channel-like incisions formed during episodes of rapid erosion followed by more or less gentle deposition (Miall 1996). Deposits of this kind are associated with fluvial environments and can be produced by means of high energy ephemeral streams of intermittent sedimentary discharge (Boggs 2006). Occurrence of these types of streams and their associated sedimentary deposits is promoted by an abrupt increase of sediment load, resulting in rapid clogging and avulsion across a floodplain (Boggs 2006). B and A depositional units are gradational deposits differing in discharge intensity of stream sedimentation cutting across the banks of a floodplain, where the former represents a more energetic deposit based on the deeply scoured top of the preceding unit (Miall 1996). B depositional units are increasingly more common up section (Figure 3), which is consistent with a gradual development of a braided stream pattern (Boggs 2006). Macro- and micro-features, such as the presence of lenticular bedding and intraformational rip-up grains support a fluvial origin for the sediments and sudden deposition from sediment-saturated streams during flooding events. A volcanic origin for the sediments is inferred from the abundance of volcanic rock fragments, feldspar crystals, and bentonite bubbles. XRD analysis also supports the volcanic characteristics of the sediments. In particular, formation, fluvial transport, and accumulation of smectite in poorly-drained, topographic low areas is interpreted to result from rapid weathering and burial of available local or regional volcanic source rocks during periods with great availability of water (Awad and Breir 1993; Yemane et al. 1987a; Moore and Reynolds 1997). In addition, based on the co-occurrence of smectite, illite-smectite, and kaolinite-smectite clay mineralogy, B depositional units probably correspond to rapid burial (Moore and Reynolds 1997). The presence of albite also supports a volcanic origin and is consistent with moderate weathering and rapid burial (Moore and Reynolds 1997). Depositional unit C (massive silt to sand). This depositional unit consists of massive to faintly-laminated clayey siltstones to fine sandstones (Figure 3, Figure 4.1-4.2, Table 1). Unweathered crystalline volcanic material is a common characteristic of most C depositional units, while fewer units also include spherical iron oxide cement/nodules and clayey intraformational clasts. C units are generally well sorted and rarely fossiliferous, preserving thin (up to 2 cm) layers and lenses that include plant impressions and poorly-preserved compressions. Some C depositional units are part of an upward-fining sequence, while others are typically bound by reworked deposits of coarser or equal grain size. Bases of C units are non-scoured and vertical transitions to units of other depositional units are commonly sharp (~ 1-2 cm at CH-89, units 9-10). Laterally, some C units can be traced stratigraphically between sections, especially in the upper parts of all sections (e.g., CH-79, unit 20, CH-93, unit 23, CH-89, unit 24). Petrographic analysis shows a massive to faintly laminated silt-rich matrix that is filled with angular elongate volcanic rock fragments, including abundant detrital glass shards and feldspar (mainly sanidine), which results in a "splintery" texture (Figure 5.10-5.11). Also present are very fine sand-size angular to subangular hornblende and pyroxene crystals, and bentonite bubbles that in some cases are rimmed by chalcedony or iron-rich cement and contain crystal inclusions (Figure 5.12-5.13). XRD analysis of C depositional units indicates the presence of feldspathic volcanic material (e.g., anorthite) and smectite (Appendix 4). Interpretation of C depositional units. Massive sedimentary structure and upward-fining grain-size distribution within C depositional units indicate deposition by gravity (Miall 1996). Based on this, C depositional units are interpreted as airfall ashes. Presence of abundant volcanic rock fragments including feldspar, hornblende, and pyroxene supports the volcanic nature of these horizons (Moore and Reynolds 1997; Boggs 2006), while the common occurrence of bentonite bubbles is consistent with deposition by aerial means (Lentfer and Torrence 2006). The abundance of C depositional units, some of which can be traced stratigraphically between sections (e.g., Figure 3, CH-79, unit 20; CH-93, unit 23, CH-89, unit 24), is consistent with airfall ash deposition on ample parts of the landscape. Evidence of slight fluvial reworking in some C depositional units is indicated by scarce intraformational clasts and poorly developed ripples and cross-laminations on upper bedding plane surfaces. Contorted plant fossils of various sizes occur in several of these airfall deposits, which is consistent with rapid in situ burial of vegetation by aerially deposited ash (Spicer et al. 1985; Wing et al. 1993; Johnson 2007). A volcanic origin for C depositional units is supported by smectite and anorthite (plagioclase feldspar) documented by XRD analysis. A neoformational origin for smectite is most likely considering the volcanic origin of the sediments, but, in those units with evidence of fluvial transport, a detrital origin and deposition in topographic lows can also be assumed (Wilson 1999). Anorthite is consistent with moderate to mild weathering conditions (Moore and Reynolds 1997).
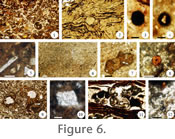
Depositional unit D (laminated clay to sand). This depositional unit consists of ripple cross-laminated siltstones and fine sandstones and horizontally-laminated claystones and mudstones that include variable amounts of sand-, silt-, and clay-size particles (Figure 3,
Figure 4.1-4.2,
Table 1). Several of the D depositional units show more than one type of lamination or have parts with massive structure, which commonly grade upward from cross, to ripple, to horizontal laminations to massive. Some of these units exhibit upward fining of sediments. Additionally, a few thin units consist of laminated claystones and siltstones hardened by calcitic cement. Some D units are vertically and laterally adjacent to other D units that can be coarser or finer, or bound between finer-grained massive layers of the C depositional unit (e.g.,
Figure 3, CH-79, unit 26). This depositional unit may also preserve scattered rooting structures, root halos, plant and other unidentifiable organic remains, and is generally poorly sorted. Also, some units contain spherical iron-oxide accumulations, intraformational clasts, mottles, and pockets or poorly-developed layers of organic material, silt, and clay (~mud drapes). Petrographic analysis documents a clay-rich matrix that is structurally massive to horizontally-microlaminated and that contains angular to subangular volcanic rock fragments, irregularly-developed microscopic layers enriched in clay/organic content, and irregular to angular clasts of organic matter rimmed by chalcedony cement (Figure 5.14-5.15). Other, less conspicuous petrographic features are angular to subangular feldspar crystals coated with brown iron-rich cement, iron oxide concretions/nodules and halos, some of which are rimmed by chalcedony cement, and small crystals of pyroxene and hornblende.
Interpretation of D depositional units. Based upon the features observed, D depositional unit represents settling out of deposits from slow-moving, sediment-saturated streams (Miall 1996). This kind of sedimentary deposit is formed in fluvial environments and commonly represents overbank deposits (Miall 1996). Some D depositional units, individually or taken as small groups, show more or less upward fining concomitant with fading (laterally or vertically) of laminations. These features, to some extent, mimic parts of heterolithic cross stratification sequences that characterize deposits of a meandering river (Thomas et al. 1987). Other units appear as poorly defined, horizontally laminated, layers that are intercalated with finer-grained massive siltstones of the C depositional unit. These may represent parts of levees deposited by sediment-saturated streams, similar to a braided pattern (Boggs 2006). Some of these horizons are bioturbated and include abundant organic material, indicating subaerial exposure and colonization of their surface by plants and other organisms. Macro- and micro-features suggest an abundant supply of sediments from local volcanoes. Clay mineralogy supports the role of volcanic material as primary weathering products of detrital and also possibly neoformational origin (smectite) (Wilson 1999). Also present in some horizons is kaolinite, which implies extensive leaching of the sediments, in environments with abundant water, which is consistent with the interpretation that this depositional unit represents overbank parts of a floodplain associated with a variable water table (Bohor and Triplehorn 1993; Moore and Reynolds 1997). In addition, deposition of mesolite in D depositional units is probably related to erosion and fluvial transport of volcanics in uplands, as this mineral belongs to the zeolite group of tectosilicates, which are found as constituents of volcanic rocks, especially basalts (Artioli et al. 1986). Depositional unit E (massive clay, mud, and sand). This depositional unit is represented by massive, thin, organic-rich claystones, mudstones, and very fine sandstones with variable amounts of silt-size grains (Figure 3, Figure 4.1-4.2, Table 1). Other characteristics include abundance of lenses, pockets, and layers of organic, clayey, and silty material that obscure other features. In addition, these units are well to poorly sorted and exhibit irregular, fine color mottles. A number of these units are stratigraphically correlatable among all or two sections and consist of thin, generally irregular layers where leaves and leaflets and other plant and organic remains (root fragments and amorphous organic matter) are concentrated. In particular, E units that occur in the basal part of all three sections, where mainly leaves and leaflets preserved in firmly indurated layers are present, represent the boundary between Ash-IV and pre-Ash-IV strata (Figure 3, CH-79 units 3-4, CH-93, units 2-3, CH-89, units 2-3). E depositional units representing the boundary between Ash-IV and the strata immediately below are underlain and overlain by coarser-grained sediments of variable organic content. Other organic-rich E units are underlain by coarser-grained sediments and overlain by layers that consist almost entirely of organic matter or are intercalated with mainly unfossiliferous layers of variable grain sizes. Petrographic thin sections show a clay-rich matrix filled with poorly developed to irregular microlaminations and microlenses (~micro mud drapes) of organic matter along with less numerous volcanic rock fragments and feldspar crystals (Figure 6.2). Also present are subrounded to irregular organic clasts of various sizes and shapes (Figure 6.3-6.4). Volcanic rock fragments and feldspars are angular to subangular and typically coated by clay, chalcedony-, or iron-rich cement (Figure 6.5). Other micro-features in some E units include hornblende, pyroxene, iron oxide accumulations, and opaque material. A small number of sandy E units are dominated by rounded to subrounded organic clasts, volcanic rock fragments, and very fine sand-size angular to subangular feldspar crystals. XRD analysis indicates the presence of smectite and kaolinite-smectite (Appendix 7). Interpretation of E depositional units. E depositional units represent organic-rich lag deposits from streams (Miall 1996). In particular, organic-rich sandy and finer-grained E units bound by coarser-grained, organic-poor sediments can be interpreted as sheet-like deposits from shallow braided streams, associated with parts of a floodplain characterized by a high water table (Miall 1996). The presence of irregular to undulate organic-rich macro- and micro-layers that include some identifiable fossil plant parts (spores, root, and/or stem) is consistent with deposition by sediment-saturated streams. Sandier E units that include numerous intraformational clasts, clay and bentonite pockets, and iron-rich accumulations indicate fluvial deposition by sediment-saturated streams on better-drained surfaces and at more proximal parts of the floodplain (McLoughlin and Drinnan 1997). Mineralogical evidence inferred from thin sections, such as the presence of feldspars and pyroxene, is consistent with a volcanic origin for the sediments, which is also supported by data from the XRD analysis. In particular, the presence of smectite indicates the occurrence of poorly-drained areas, where deposits accumulated by fluvial transport of volcanic weathering products from elsewhere in the basin (Wilson 1999). In addition, the presence of kaolinite-smectite implies increasingly greater weathering under very humid conditions and better drainage (Moore and Reynolds 1997). Depositional unit F (bioturbated, massive clay to sand). This depositional unit is characterized by massive, intensely bioturbated claystones to very fine sandstones (Figure 3, Figure 4.1-4.2, Table 1). A number of F depositional units also display poorly-developed wavy laminations, which consist of layers rich in organic, clay-, and silt-size particles or a combination of these. The characteristic common to all F depositional units is widespread occurrence of rooting structures and other organic debris including leaf impressions and compressions. Other macroscopic features of some of these units are fine color mottles, root halos, slicken-side surfaces, and pockets or lenses that are either made of organic-, clay-, or silt-rich sediments or a combination of these. F depositional units are generally well sorted, are bound by coarser- or finer-grained sediments or a combination of these, and commonly grade vertically and laterally into organic-rich horizons (E and G depositional units), or, as in the pre-Ash-IV strata (Figure 3, CH-79, units 1-2, CH-93, unit 2, CH-89, unit 1), are stratigraphically continuous between sections. Petrographically, the F depositional units are characterized by a massive to weakly-laminated clay- and organic-rich matrix (Figure 6.6). Some units also display microscopic lenses/pockets of organic-rich material and irregular to subrounded grains of organic matter; especially abundant are root sections bordered by iron-rich cement (Figure 6.7-6.9). Other microfeatures include subangular to subrounded volcanic rock fragments and feldspars. Also present in some units are blocky siderite crystals, concretionary iron-oxides, and opaque particles (Figure 6.10). XRD analysis of the sediments indicates the presence of smectite, kaolinite, and siderite (Appendix 8 and Appendix 9). Interpretation of F depositional units. F depositional units indicate plant-populated horizons associated with pedogenesis (Miall 1996). Abundance of macro- and micro- rooting structures, organic clasts possibly representing spores or other plant parts, and poorly preserved clay- and organic-rich microlaminations is consistent with fluvial deposition from suspension and at a slow pace alternating with plant growth intervals, in proximal/distal parts of a floodplain. Gradation of F depositional units into organic-rich horizons indicates an increasingly higher water table, while stratigraphically correlatable units (pre-Ash-IV) indicate more extensive and temporally durable soil formation than individual units. Occurrence of blocky siderite crystals in some of these horizons indicates formation in a reducing environment, low concentrations of aqueous sulfide, circum-neutral pH, and soil pCO2 > 100,000 ppmV (Pearson, 1979; Postman, 1982). However, concretionary iron-oxide nodules in F depositional units are consistent with more oxidizing conditions, slower sedimentation rate, and subaerial exposure. The often mottled appearance of the F units is consistent with temporal variation of the water table resulting in periods characterized by periodically alternating anoxic and oxic conditions. Macro and micro-features and XRD data indicate a volcanic origin for the source sediments. In particular, smectite and kaolinite, which are the most common phyllosilicates preserved in these units as well as in the other depositional units described, are consistent with variable weathering conditions depending upon availability of water in the environment and duration of erosion and deposition processes (Moore and Reynolds 1997). As such, their presence is consistent with rather limited weathering and rapid burial of sediments of volcanic origin (Moore and Reynolds 1997). The co-occurrence of all these minerals can result from rapid evolution of the fluvial landscape. As a result, these minerals represent moderate weathering and demonstrate the temporary nature of this soil-forming environment (Borchardt 1989 and references therein). Depositional unit G (lignified to semi-lignified organic matter). This depositional unit consists of thin layers of organic matter, including poorly-preserved plant fragments (Figure 3, Figure 4.1-4.2, Table 1). G depositional units are thinner than most other depositional units, not laterally correlatable between sections, and are bound by coarser-grained sediments from which they are separated by sharp erosional or gradational contacts. Petrographic thin sections document a matrix that consists of coarse to fine-laminated organic matter, where most of these laminations appear as undulating and convoluted layers not firmly appressed to each other (Figure 6.11). In the spaces left between these layers usually are found small rounded volcanic rock fragments (Figure 6.12). XRD analysis of G units indicates the presence of smectite and kaolinite (Appendix 10). Interpretation of G depositional unit. Based on the features observed, G depositional units represent in situ accumulations of organic matter (~lignites) in shallow, ephemeral pools of water located in parts of a floodplain characterized by a high water table (Miall 1996). The distinction between the E, F, and G depositional units is mainly based on the ratio of organic matter to mineral content within a horizon, while common to them is that their deposition occurred on parts of a floodplain characterized by a high water table (Miall 1996). In the case of the G depositional unit a consistently high water table prevailed, promoting the accumulation of organic matter and inhibiting oxidation. However, the microlayered structure of the G depositional unit, consisting of undulating and convoluted layers of variable thickness, indicates periods of intermittent deposition characterized by periodic subaerial exposure and drying (Boggs 2006). Origin of smectite and kaolinite in the G unit is most probably detrital and consistent with variable weathering of volcanic rocks (Moore and Reynolds 1997; Wilson 1999). In particular, kaolinite commonly accumulates in swampy environments characterized by abundant water leaching through sediments and, therefore, its presence is consistent with an environment with abundant precipitation and well-drained soils (Bohor and Triplehorn 1993). However, peat does not accumulate in well-drained soils. Therefore, kaolinite present in these horizons must have either been transported from well-drained environments or formed by leaching of organic acids and chelates related to the transformation of peat to coal in the burial environment (Bohor and Triplehorn 1993). Smectite forms in poorly drained soils rich in alkaline material and weathers to kaolinite under humid conditions, which is consistent with increasing weathering and rapid burial of the sediments in topographic lows of the floodplain (Moore and Reynolds 1997). Hiatuses: Formation of PaleosolsThree different paleosol types have been identified within the bottom and middle parts of the stratigraphic sections exposed along the Margargaria River (Figure 3). In particular, based upon evaluation of the relative importance of a series of pedogenic features/processes including relative organic matter content, soil structure and mineralogy, mottling, horizonation, and color, these three paleosols types are classified according to Mack et al. (1993). Morphological characteristics and lateral and vertical distribution of each paleosol are described below.
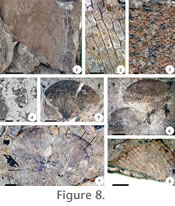
The basalmost units in all three stratigraphic sections, which correspond to the thin set of strata below Ash-IV (e.g., Figure 3, CH-79, unit 1-3), consist of a succession of poorly-developed, gley-colored, organic-rich, mudstones horizons that grade upward to a layer enriched in organic content (O-horizon) containing abundant leaves (Figure 7.5-7.6). Gley coloration refers to low chroma, greenish-blue-grey soil colors typical of wetland conditions (Mack et al., 1993). The roots (up to 3 cm wide and ~20 cm in long) consist of compacted organic matter oriented vertically and obliquely relative to the orientation of the strata (Figure 7.8). Based on these features and on the presence of homogeneously distributed elliptical to variously shaped bluish mottles and a structure consisting of coarse, wedge-shaped aggregates with slicken side surfaces and fossilized roots (Figure 7.5, 7.7), this succession is interpreted as a Vertisol (Mack et al., 1993). Vertical rooting structures indicate free-drainage, and therefore at least intermittent well-drained conditions (Retallack 1990). Slicken planes in this profile indicate shrink-swell processes probably related to periodic rise and fall of the local water table, or seasonal flooding from overland flow (Mack et al. 1993). These features may also be observed in poorly developed Gleysols as well as in Vertisols (Mack et al. 1993). Clay minerals present in the strata representing this Vertisol are dominated by smectite, kaolinite, and kaolinite-smectite, all most likely detrital in origin and representing variable weathering conditions (Appendix 7, Appendix 8). These data are consistent with soil formation upon proximal parts of a floodplain in a seasonal environment (Mack et al. 1993; Tabor and Montañez 2004). Sets of strata consisting of an organic-rich siliciclastic horizon(s) (A-horizon) that grade into a lignified/semi-lignified organic matter horizon (~lignite, O horizon) that apparently accumulated in situ occur in the middle part of all sections. Numerous compressions and impressions of fronds, leaves, and tabular roots are present in these profiles (Figure 7.10). The mineral layer(s) characteristically is structureless to faintly laminated, or sometimes blocky to prismatic, silty claystone to clayey fine sandstone of rather limited vertical extent (up to ~ 15 cm) (Figure 7.11). Siderite is present in the lower mineral horizon, while kaolinite and smectite are found in both the lower and upper horizons that make up these paleosols (Figure 6.10, Appendix 8, Appendix 9, and Appendix 10), which are classified as Histosols (e.g., Figure 3, CH-89, unit 11-13, Figures 7.9, 7.11; Mack et al. 1993). Other individual strata showing increasing organics consisting of variously preserved plant parts appear as partly eroded Histosols (e.g., CH-89, unit 17). Histosol horizons are characterized by their gley coloration, which is more intense as the organics concentrate. Small (up to 1cm across), subspherical reddish mottles occur in the lower layer of siliciclastic material (Figure 7.12). Discontinuous and irregular silt-rich layers or pockets commonly occur within the mineral horizon (Figure 7.13). Smectite is consistent with slight weathering and rapid burial of sediments of volcanic origin (Moore and Reynolds 1997). Kaolinite is a clay mineral precipitated in acidic solutions, depleted in bases, and its formation in soils is related to periods of around ten thousand years, precipitation exceeding evaporation, and its presence in the Histosols indicates greater weathering than smectite (Keller 1956; Wilson 1999). The episodic supply of volcanic ash onto the paleolandscape probably provided most of the material that, after leaching, weathered to kaolinite, which accumulated in lowland areas where small ponds developed. Additionally, it is also possible that leaching of organic acids from plant remains during peat formation equally contributed to the presence of kaolinite in the Histosols (Bohor and Triplehorn 1993). The deposition of volcanic ash results in impermeable horizons that promote pond development, which may have favored the formation of kaolinite in the lignites (Crowley et al. 1989, 1994). Co-occurrence of siderite and poorly consolidated plant remains, including tabular roots are consistent with in situ accumulation of organic rich horizons in low lying, waterlogged substrates characterized by a shallow, anoxic or dysoxic water table (Staub and Cohen 1978; Mack et al. 1993; Retallack 1990). The presence of Fe mottles indicates intermittent change between oxic and anoxic soil atmosphere (Vepraskas 1994), while pockets/lenses of ashy material are probably related to erosion by small-scale streams that traversed the landscape; as a result these paleosols appear to have formed by several separate episodes of sediment deposition and plant growth. Paleobotanical DataCollection of macrofossils from the volcaniclastic strata studied here yielded a flora that consists of vegetative and reproductive fern and angiosperm specimens, including Acrostichum (Pteridaceae), Marsilea (Marsileaceae), Equisetum (Equisetaceae), Salvinia (Salviniaceae), Cyclosorus (Thelypteridaceae), Blechnum (Blechnaceae), Cynometra (Fabaceae), Typha (Typhaceae), Alstonia (Apocynaceae), Pandanites (Pandanaceae), Hyphaene (Arecaceae), and Macaranga (Euphorbiaceae) (García Massini et al. 2006; García Massini and Jacobs 2009). In addition to these fossils, there are remains of other less well-preserved plants including ferns (e.g., Actiniopteris-like) and angiosperms (e.g., Rubus-like), which are mentioned here to more clearly show the stratigraphic distribution of ferns and angiosperms relative to each other. Ferns occur singly or in combination as the dominant vegetation in several units of all depositional units starting at around the middle part of all sections, except for the A depositional unit, which does not contain macrofossils (except for some root fragments). Angiosperms co-occur with the ferns as subordinate elements in most of these units, except for the lowermost units of all sections of Ash-IV and the thin set of strata below, where angiosperms dominate and ferns are absent. Ferns
Family Pteridaceae Kirchner, 1831
Description and type of preservation. This fossil consists of impressions and compressions of fragments of linear-lanceolate sterile and fertile pinnae (Figure 8.1). A few fossils consist of fragments of leaves showing petiolate pinnae. Diagnostic features of this fossil include reticulate venation without included free veinlets and acrostichoid arrangement of stalked sporangia (Figure 8.1-8.4). Additional Acrostichum leaf remains were previously described from the Margargaria River and other tuffaceous strata of the Chilga beds, but those did not include fertile specimens, and therefore they were assigned to a fossil species that includes only sterile remains (García Massini et al. 2006). Stratigraphic distribution and mode of preservation. Remains of the hydrophilic fern Acrostichum occur commonly lying flat in organic-rich layers (D, E, F, and G depositional units in CH-89; E, F, and G in CH-93; F and G in CH-79) in all three geologic sections (e.g., Figure 3, CH-79, unit 18-19) and embedded in one airfall tuff in the south section (C depositional unit) (Figure 3, CH-79, unit 20). Modern distribution and ecology.Extant members of this genus occur inland in wetland settings and in coastal environments associated with mangroves in acidic to neutral pH and high salinity organic- and clay-rich soils (Rao et al. 1973; Tomlinson 1986). Generally, the occurrence of Acrostichum today is promoted by increasing soil acidification and by both anthropogenic and natural ecological disturbance (Rao et al. 1973; Tomlinson 1986; Thanikaimoni 1987; Saenger 1998). The ecological distinctiveness that allows Acrostichum to inhabit these kinds of environments derives at least partially from its dispersal ability as a pioneer plant adapted to rapid colonization of disturbed habitats devoid of local vegetation (Tomlinson 1986; Saenger 1998). Paleoecological significance. In this regard, the presence of this fossil within the E, F, G, and C depositional units of the Ash-IV tuff sequence at the Margargaria River is consistent with the ecological role of modern Acrostichum.
Family Marsileaceae Mirbel, 1802 Specimens. CH-79-229B (Figure 8.5), CH-79-227 (Figure 8.6), CH-79-230A (Figure 8.7), and CH-79-32B (Figure 8.8). Description and type of preservation. This fossil consists of impressions and compressions of fan-shaped isolated individual leaflets and a few pairs of leaflets with discernible portions of a third and fourth leaflet diverging from a common straight petiole (Figure 8.5-8.7). Venation is dichotomous with veins that anastomose and form fusiform areoles that end in a marginal vein (Figure 8.8). Leaf morphology and venation pattern of the fossil are identical to extant Marsilea (Tryon and Tryon 1982). All other fossil Marsilea known worldwide were transferred to the morphogenus Marsileaphyllum based on a similar, but not identical, leaf morphology and venation (Nagalingum 2007). Stratigraphic distribution and mode of preservation. Marsilea fossils from the Margargaria River localities occur more or less flat along bedding planes in a ~10 cm, massive, overbank silt-rich layer (F depositional unit) in the middle part of the south section (Figure 3, CH-79, unit 18). This layer grades upward into a thin, poorly-consolidated lignite (G depositional unit) in which badly-preserved Marsilea leaflets are also found. Modern distribution and ecology. Extant members of Marsilea occur worldwide in temperate to tropical environments rooted in sandy but especially in muddy substrates in ponded settings and in ephemeral aquatic habitats from lowlands up to 1950 meters, such as in some areas of Ethiopia (Johnson 1986; Launert 2003). Marsilea's diversity is greater in Africa than in all other continents, and this has been interpreted to be the result of evolutionary processes related to adaptation to seasonally dry, semi-arid climates (Kornás 1985). Paleoecological significance. The presence of Marsilea in these layers is interpreted to represent ponded settings of ephemeral nature. In addition, Marsilea, a cosmopolitan member of the heterosporous water ferns, of high diversity in Africa, is considered a drought-tolerant taxon usually found in seasonal environments. On this basis, its presence in the Margargaria River localities indicates a seasonally dry climate (Kornás 1985; Launert 2003; Aldasoro et al. 2004; Nagalingum et al. 2007). Moreover, Marsilea's occurrence in Africa is negatively correlated with humidity, which led to the hypothesis that its greater diversity in the Namibia-South Africa region is due to the early development of deserts during the Cenozoic (Aldasoro et al. 2004), and this also supports the interpretation that seasonally dry climatic conditions should have characterized Marsilea's habitat in the Margargaria River.
Family Equisetaceae Michaux ex
De Candolle, 1804
Description and type of preservation. Fossil Equisetum has been identified based on the presence of dispersed fragments of impressions and compressions of hollow ribbed stems with nodes formed by a sheath of material representing the remains of whorled leaves (Figure 9.1- 9.2) (Tryon and Tryon 1982; Verdcourt 1999a; Smith et al. 2006). Stratigraphic distribution and mode of preservation. Equisetum stems occur rooted in situ or lying flat in organic-rich overbank reworked muddy and clayey layers (F and G depositional units) in the central and south sections (e.g., Figure 3, CH-79, unit 18). Modern distribution and ecology. Equisetum inhabits wetland settings and other habitats characterized by a high water table, such as by rivers and streams, in temperate to tropical latitudes including one species native to North, East, Central, and South Africa that occur at elevations as high as 2100 meters (Tryon and Tryon 1982; Verdcourt 1999a). This fern is characterized by being a rapid colonizer of disturbed habitats (Tryon and Tryon 1982; Verdcourt 1999a). Examples of modern Equisetum that dominated early colonization of volcanically disturbed environments by regenerating from buried rhizomes, broken plant fragments, and spores are known from temperate and tropical latitudes of North America (Spicer 1991; Weber et al. 2006). In addition, fossil Equisetum has been found elsewhere preserved in situ in swampy to marshy, floodplain, and volcanic paleoenvironments in association with other ferns, which has been interpreted to have resulted from rapid sedimentation (e.g., Collinson 1988; Falder et al. 1999; Falcon-Lang and Cantrill 2002; Cantrill and Hunter 2005 and references therein). Paleoecological significance. The presence of in-situ Equisetum stems in the same layer preserving Marsilea supports the interpretation that this setting was of ephemeral duration. Co-occurring with Marsilea and Equisetum is Acrostichum, and this also suggests that this setting resembled a short-lived inundated terrain, such as a floodplain where the water table was temporarily high or an ephemeral pond.
Family Salviniaceae Martynov, 1820 Specimens. CH-93-62B (Figure 9.3), CH-93-48A (Figure 9.4), CH-79-21A (Figure 9.5), CH-93-69B (Figure 9.6), and CH-93-81 (Figure 9.7). Description and type of preservation. Salvinia consists of dispersed individual ovate-obovate leaflet impressions and compressions (Figure 9.3-9.4). Also present are coarsely rugulate, ellipsoidal megaspores with a prominent 2-lobed acrollamella that obscures the trilete mark (Figure 9.5). A few specimens of Salvinia consist of two leaflets folded on top of each other from which emerge two filaments that resemble a submerged root-like leaf (Figure 9.4). Secondary venation consists of proximally-curved lateral veins that arise from a prominent midvein and anastomose, forming irregular to polygonal areoles that collect together into a marginal common vein (Figure 9.6-9.7). The areoles further anastomose into smaller components that appear clustered, resembling the bases for insertion of trichomes observed in extant taxa (Figure 9.7). This fossil has been assigned to extant Salvinia based on venation and morphology of leaflets, while megaspores also are morphologically identical to extant members of this genus (Tryon and Tryon 1982). Stratigraphic distribution and mode of preservation. Salvinia leaflets are mainly found lying flat or slightly curved along breaking planes in poorly laminated to massive mudstones, claystones, and lignites (F and G depositional units) in the central and north sections (e.g., Figure 3, CH-93, unit 11-13, CH-89, unit 17). These fossils are also poorly-preserved lying flat on some coarser-grained overbank layers in the central (E depositional unit; e.g., CH-93, unit 17) and north sections (D and E depositional units; e.g., Figure 3, CH-89, units 12-14). Salvinia megaspores are found in all sections in organic-rich claystones and lignites (F and G depositional units) (e.g., CH-79, unit 18). Modern distribution and ecology. Extant Salvinia is a rapidly growing taxon that inhabits rivers or other aquatic settings mainly in tropical latitudes including Africa with one species mainly native to the eastern part of the continent (Schelpe 1961; Tryon and Tryon 1982; Verdcourt 2000). Colonies of Salvinia that grow over generations form mats centimeters thick that create an anoxic environment below (Schelpe 1961). In addition, Salvinia co-occurs with Marsilea in dry regions of Africa, which is consistent with the fact that both ferns, independently, are negatively correlated with humidity (Aldasoro et al. 2004). Paleoecological significance. The presence of this fossil in overbank layers separated and capped by lignites supports previous data indicating frequent inundation and formation of ponded settings in the environment. In particular, Salvinia occurrence could have contributed to organic matter preservation in the lignite units, especially in the central section (CH-93, units 11-14) where this fern is most abundant. Salvinia leaflets and megaspores co-occur with the remaining ferns in these overbank layers and in similar layers in the north section (e.g., CH-89, unit 17), except for Marsilea, which only occurs with Salvinia megaspores in the south section (CH-79, units 18-19). This probably reflects landscape heterogeneity, suggesting that these two aquatic ferns were contemporaneous in the environment, but restricted to different ecological niches. Salvinia is not adapted to terrestrial conditions, in contrast with Marsilea (Schelpe 1961; Kornás 1985; Tryon and Tryon 1982; Schneider and Pryer 2002), and therefore it must have lived in a wetter part of the environment.
Family Thelypteridaceae
Pichi Sermoli, 1970
Description and type of preservation. This fern consists of alternately arranged, sterile and, apparently, fertile, bi-pinnate frond fragment impressions and poorly-preserved compressions that are gradually reduced and that are circinate in the bud (Figure 9.8-9.10). Possible fertile specimens from a locality collected elsewhere in the Chilga basin display a series of black rounded spots, reminiscent of sori, borne in two parallel rows on the secondary veins of each pinnule (Figure 9.10). Pinnules are pinnately veined and have a typical venation overall, in which the distal and proximal veins of adjoining pinnules meet at the sinus and fuse into a single vein that extends vertically and perpendicularly away from the costa (Figure 10.1). The following veins of each pinnule meet at the sinus, synchronically or nearly so, but do not fuse and end at its upper limit (Figure 10.1). Fossil fronds have been assigned to Cyclosorus (Thelypteridaceae) based on pinnule venation and pinnae arrangement and morphology (Tryon and Tryon 1982; Kramer and Green 1990). Stratigraphic distribution and mode of preservation. In situ Cyclosorus frond mats, where individuals embedded within an airfall tuff (C depositional unit) can be traced vertically and laterally for a short distance within this unit, occur as the dominant vegetation in the south section (Figure 3, CH-79, unit 20). Smaller Cyclosorus frond fragments embedded or lying-flat in airfall tuffs, lignites, and overbank layers (C, F, and G depositional units) are also found, singly or co-occurring with some or all of the other ferns in all three sections (e.g., Figure 3, CH-79, units 19; CH-93, unit 23; CH-89, unit 17) and in some reworked tuffs (D depositional unit) in the south and north sections (e.g., CH-79, unit 26; CH-89, unit 14). Modern distribution and ecology. Today's distribution of Cyclosorus includes swampy habitats in the tropics and subtropics of South America and Africa, especially the southeastern and western parts of this continent (Tryon and Tryon 1982; Kramer and Green 1990). This fern is characterized by its ability to colonize disturbed sites such as landslides and roadside banks (Tryon and Tryon 1982; Judd et al. 2007). Paleoecological significance. The presence of Cyclosorus in the Margargaria River is consistent with the pioneer ecological role of extant taxa in this genus (Barthel 1976; Riegel et al. 1999; Collinson 2002), and indicates the common occurrence of short-lived settings during the deposition of the Ash-IV sequence.
Family Blechnaceae Newman, 1844 Specimens. CH-93-90 (Figure 10.2), CH-87-8 (Figure 10.3-10.5), and CH-93-70 (Figure 10.6-10.7). Description and type of preservation. This fossil consists of impressions and compressions of linear lanceolate fertile and sterile dispersed individual pinnae (Figure 10.2-10.3). Fertile specimens, which were recovered at a different locality than the Margargaria river localities, show two rows of exindusiate sporangia arranged in a linear pattern in a comissure that runs parallel to the midvein (costa) (Figure 10.3). Stalked sporangia are preserved as compressions and contain discrete numbers of ellipsoidal monolete spores (Figure 10.4-10.5). Venation is free dichotomous, where secondary veins bifurcate twice (Figure 10.6). Secondary veins end at the margin forming a toothed-crenulate pattern (Figure 10.7). The presence of fossil Blechnum (Blechnaceae) from Ash-IV is inferred based on morphology of sterile and fertile fronds (Tryon and Tryon 1982). Stratigraphic distribution and mode of preservation. Numerous Blechnum pinnae occur flat or curved along bedding planes in lignites and overbank layers in the central section (F and G depositional units) (e.g., Figure 3, CH-93, unit 11-13) and in overbank layers in the north section (F depositional unit) (Figure 3, CH-89, unit 17); it co-occurs with all the other ferns, except Marsilea. Modern distribution and ecology. Blechnum is a cosmopolitan genus best represented in cooler temperate zones or higher altitudes in Africa (1500-2300 meters high) (Kramer and Green 1990; Aldasoro et al. 2004; Parris 2006). In addition, extant members of this family inhabit montane wet evergreen forests, wet grasslands, and swamps, and form thickets in disturbed sites in tropical latitudes (especially in the Southern Hemisphere) (Tryon and Tryon 1982; Kramer and Green 1990; Parris 2006; Judd et al. 2007). Paleoecological significance. The presence of Blechnum in the Ash-IV deposits at the Margargaria River localities, ~1800 m above sea level today, may indicate similar environmental conditions and substantial uplift of the Ethiopian Plateau by the Late Oligocene (Hoffman et al. 1997 and references therein). The ability of Blechnum as a pioneer taxon is consistent with the biology of the other ferns present in these deposits and, independently, also indicates a landscape characterized by a rapidly changing physiography. Angiosperms
Family Fabaceae Lindley, 1836
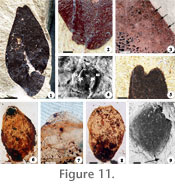
Description and type of preservation. This fossil consists of impressions and compressions of small-sized, slightly oblong-elliptic, dispersed leaflets with an oblique-acute base (Figure 11.1). Leaflets are asymmetrical, have a short, pulvinate, petiolule, a slightly eccentric primary vein, faint secondary brochidodromus venation, an entire margin, paracytic stomata, and an acute emarginate apex with a mucron (Figure 11.2-11.5). All these features combined associate this fossil with extant Cynometra (Mackinder 2005; Hawthorne and Jongkind 2006). Stratigraphic distribution and mode of preservation. Cynometra leaflets occur contorted along bedding planes or lying flat in weakly laminated clayey siltstones, mudstones, and silty claystones (D, E, and F depositional units) in the basalmost units of Ash-IV in all sections and in the thin stratum immediately below (pre-Ash-IV), respectively (e.g., Figure 3, CH-79, unit 3-5). Cynometra is qualitatively the most abundant fossil plant from these basal units. Modern distribution and ecology. Living Cynometra are trees or shrubs common to western and central Africa in lowland rain and swamp forest, often along rivers (Hawthorne and Jongkind 2006; Judd et al. 2007). Members of this genus are also present in seasonally dry forest, woodland, and bushland or thicket, where some species grow gregariously and form dominant stands (Mackinder 2005). Paleoecological significance. Because of the wide range of environments in which species of Cynometra can be found today, the paleoecological setting cannot be determined more narrowly than bushland to rainforest based upon this genus. However, additional Cynometra leaves have been found previously in the Guang River locality of the Chilga beds, approximately 20 meters stratigraphically below the Margargaria species described here, forming part of a parautochthonous plant assemblage that was interpreted to represent a riparian forest community (Pan 2007). Cynometra from the Margargaria River localities is morphologically indistinguishable from the Guang River fossil, and therefore, it may be assumed that they both represent the same taxon and inhabited comparable riparian environments separated in time and space.
Family Typhaceae Jussieu, 1789 Specimens. CH-79-4 (Figure 11.6, 11.9) and CH-79-26 (Figure 11.7-11.8). Description and type of preservation. This fossil consists of compressions of barrel-shaped seeds (Figure 11.6-11.9). Characteristic of these seeds is an operculum that occludes the proximal end, where a ragged area representing a collar of cells is seen in those individuals that lack an operculum (Figure 11.6-11.8). A darker zone of cells is commonly present towards the base of these seeds (Figure 11.6, 11.8) and may represent the chalazal region present in modern taxa. A micro-pitted area surrounding the perimeter of some of the specimens (Figure 11.9) is reminiscent of the part of attachment for bristles (part of the perianth) present in modern taxa. The features displayed by the fossil are consistent with extant Typha (Herendeen and Crane 1995; Judd et al. 2007). Stratigraphic distribution and mode of preservation. Numerous small (~0.1 cm long) Typha seeds laying along bedding planes in clayey organic rich and lignitic layers in the south section (F and G depositional units) (Figure 3, CH-79, unit 18-19) co-occur with Marsilea, Cyclosorus, Salvinia (megaspores), Acrostichum, and Alstonia. Modern distribution and ecology. Members of the family Typhaceae or "cat tails" are cosmopolitan taxa that inhabit river or sedge meadows, swamps, and fresh water marshes (Judd et al. 2007). They can be found in sandy, shallow-water settings with fluctuating water tables; in soils that may be saturated seasonally and undergo periodic drying, in habitats that may show moderate peat development and, especially, in disturbed habitats (Smith and Kaul 1986; Zomlefer 1994). In particular, an increase in the abundance of Typha has been related to the spread of swampland following forest disturbance in successional volcanic environments in New Zealand and in Mount Saint Helens in North America (Spicer 1991; Wilmshurst and McGlone 1996). One reason Typha may rapidly colonize open landscapes could be the large number of very small seeds produced by a single plant facilitating transportation by wind currents in open landscapes (Smith and Kaul 1986; Zomlefer 1994). Each wind-dispersed unit comprises an achene-like fruit plus bristles (persistent perianth or "pappus") that, after dispersal, ultimately split lengthwise to release the single seed (Smith and Kaul 1986). Paleoecological significance. Abundance of Typha seeds in the studied units indicates an open landscape with a shallow freshwater pond or a slow flowing stream with abundant aquatic vegetation, something that may have been promoted by volcanic disturbance of the environment. Co-occurrence of Typha with Salvinia megaspores, Marsilea, and Acrostichum, each of which are known to occur in swampy settings but within more or less discrete zones among these (i.e., emergent, shoreline-rooted vegetation), is consistent with high sedimentation rates that rapidly buried the vegetation allowing its preservation as a whole. Further, the occurrence of dicot leaves (Alstonia) towards the top of this layer is consistent with the idea that as the swamp or pond dried, the zone of rooted vegetation gradually increased.
Family Apocynaceae Jussieu, 1789
S Description and type of preservation. This fossil consists of lanceolate to oblanceolate petiolate leaf impressions and poorly-preserved compressions with an entire margin, a decurrent base, and an acute-acuminate apex (Figure 12.1-12.3). Venation is pinnate with many pairs of secondary veins coming off the main vein at a right to acute angle and parallel to each other (Figure 12.4). In addition, all secondaries run to the margin where they collect in a thick common marginal vein (Figure 12.4). These characters, especially the presence of a large and thick marginal vein where parallel secondary veins collect, are found in the extant genus Alstonia (Hawthorne and Jongkind 2006). Stratigraphic distribution and mode of preservation. Alstonia leaves occur lying flat or slightly contorted along bedding planes within organic-rich claystones and mudstones and semi-consolidated lignitic layers (G and F depositional unit) in the south and central sections (e.g., Figure 3, CH-93, unit 15, CH-79, unit 18-19). Leaves of Alstonia are also found embedded and completely curled up in some reworked and airfall tuffs (B, C, and D depositional units) in the south section (Figure 3, CH-79, units 20, 22, 24, 26). Modern distribution and ecology. Extant Alstonia are trees and shrubs that occur as pioneer taxa in primary and especially in secondary swamp successional forests (Hawthorne and Jongkind 2006). They have a buttressed base when they live in swampy settings, which are sometimes characterized by a seasonal climate, at low and medium altitudes in sub tropical to tropical latitudes, including west, central, and northeastern Africa and southern Asia (Hawthorne and Jongkind 2006; Judd et al. 2007; African Flowering Plants Database). Paleoecological significance. The presence of Alstonia is consistent with the occurrence of an early successional open swampy environment of reduced extent, which supports the paleoenvironmental interpretation based on other angiosperm and fern taxa (Marsilea, Salvinia, Acrostichum, Typha) in the same strata.
Family Pandanaceae Brown, 1810b
Description and type of preservation. This fossil is characterized by narrow, strap-like, leaf impressions and compressions that appear M-shaped in transverse section and that have their margins ornamented with numerous regularly-spaced, hook-like, upwardly-oriented, small teeth (spines) (Figure 12.5, Figure 13.1). Other features include numerous parallel veins of one order, pairs of which appear to be connected by perpendicularly to slightly obliquely (weakly-developed) secondary veins (Figure 13.1). Some specimens display a keel, or a slightly inflexed or thickened part of the leaf (Figure 13.1), where additional spines are present in modern Pandanus, but it appears that these have been lost in the fossil. Despite the absence of this last character, the morphological features present in this fossil are consistent with its affinity to Pandanites, which is the fossil representative of the extant genus Pandanus (Hawthorne and Jongkind 2006; Kvaček and Herman 2004; Burger 1967). Stratigraphic distribution and mode of preservation. Pandanites occurs lying flat or slightly curved along bedding planes within organic rich claystones and mudstones (G, E, and F depositional units) in the central and north sections (Figure 3, CH-93, unit 11-13, CH-89, unit 17). Modern distribution and ecology. Modern Pandanaceae are trees, climbers, or shrubs with prop roots that inhabit lowland swamps including coastal environments in oceanic islands, riverine forests, and the shores of rivers in evergreen forests up to 1400 meters in the Old and New World tropics and subtropics (Burger 1967; Beentje 1993; Hawthorne and Jongkind 2006). It also occurs in secondary forest as gregarious patches along streams and around swamps (Beentje 1993; Hawthorne and Jongkind 2006). Paleoecological significance. The presence of Pandanites is consistent with a moist and warm environment in a landscape traversed by rivers and dominated by swamps. Pandanites would have concentrated where the water table was high.
Family Arecaceae
Schultz-Schultzenstein, 1832 Specimen. CH-93-99A (Figure 13.2). Description and type of preservation. This fossil consists of compressed petiole fragments that are armed with robust spines of similar size (Figure 13.2). The spines in some specimens are paired and regularly spaced while, in others, it is not possible to determine due to poor preservation (Figure 13.2). Spines are either curved or perpendicular to the petiole, and have relatively broad, flat, arcuate bases that connect with adjacent ones (Figure 13.2). The presence of armed petioles bearing spines with these characteristics indicates affinities with the palm genus Hyphaene (subfamily Coryphoideae) (Hawthorne and Jongkind 2006; Pan et al. 2006; Uhl and Dransfield 1987). Stratigraphic distribution and mode of preservation. Hyphaene petiole fragments occur embedded at variable orientations in airfall and reworked tuffs in the central (B and C depositional units; CH-93, unit 23-24) and north sections (C depositional unit; Figure 3, CH-89, unit 24, 26). Modern distribution and ecology. Hyphaene occurs in Africa in arid to semi-arid climates, particularly associated with seasonal streams (Burger 1967; Uhl and Dransfield 1987; Hawthorne and Jongkind 2006). Hyphaene species are found in gallery and coastal forests, floodplains, palm savannahs, and along creeks behind mangroves (Burger 1967; Hawthorne and Jongkind 2006). Paleoecological significance. The presence of Hyphaene in Ash-IV is consistent with a riparian environment, but as palms have been previously found occurring in early colonization stages of volcanically disturbed environments during the Cenozoic (e.g., see Collinson 2001 for references), its presence probably also reflects adaptation to disturbance. A Hyphaene species different from the taxon described here has been found in older strata in Chilga in both airfall tuff and organic-rich mudstones (Pan et al. 2006), consistent with Hyphaene from Chilga characterizing seasonal, ecologically unstable, environments.
Family Euphorbiaceae Jussieu, 1789 Specimens. CH-79-236 (Figure 13.3, 13.5) CH-79-20B (Figure 13.4), and CH-79-232 (Figure 13.6). Description and type of preservation. This fossil consists of leaf impressions and poorly-preserved compressions that are broadly lanceolate, have a cuneate to obtuse base, an acute apex, and an entire margin (Figure 13.3). Venation is tri-nerved (actinodromous), where the primary veins are prominent and robust (Figure 13.4). Intersecondary veins split off from the lateral main veins, arch, and reach the margin independently (Figure 13.4). Tertiary venation is difficult to observe but it looks more or less scalariform (alternate percurrent) (Figure 13.4). The petiole is relatively long and appears hollow (Figure 13.5). Two small dots at the end of the petiole in a few specimens represent extrafloral nectaries (Figure 13.5). Also, disc-shaped glandular hairs are present in some specimens (Figure 13.6). The range of features displayed (i.e., extrafloral nectarines and disc-shaped glandular hairs) associates this fossil to both Macaranga and Mallotus of the Euphorbiaceae; however, the absence of stellate or stellately bundled hairs, the leaf shape, and a long petiole indicates a closer relationship with Macaranga (Kulju et al. 2007). Stratigraphic distribution and mode of preservation. Macaranga occurs flat or contorted, sometimes completely curled up, in some airfall tuffs (C depositional unit) in the south and north sections (e.g., Figure 3, CH-79, unit 20, CH-89, unit 24). Modern distribution and ecology. Extant Macaranga is a fast growing pioneer species of tropical to subtropical montane evergreen forest, and may locally be common to dominant in wetter vegetation types, such as swamp forests, gallery forests, and open seasonally swampy settings (Hawthorne and Jongkind 2006; Judd et al. 2007). It regenerates vigorously in clear-felled areas, secondary forest, forest edges, riverine forests, and especially in ecologically disturbed places (Slik and van Welzen 2001). Paleoecological significance. The presence of fossil Macaranga is consistent with the presence of an open landscape with at least seasonally abundant water in a periodically disturbed area. In addition to the fossils already described, there are other plant remains that cannot be assigned confidently to any known group, although they display enough features to allow suggestion of affinities to extant taxa. These fossils are briefly commented on here since they provide additional information about the distribution of ferns and angiosperms within the studied geologic sections. In addition, taphonomy of these additional plants generally corresponds with depositional setting. For example, lignitic strata preserve mostly horizontal and flat-lying fossil assemblages, whereas airfall deposits preserve ferns and angiosperms embedded in ash, sometimes curled, and rarely horizontal. In most cases plant fossils appear to have come from local communities determined by disturbance and/or edaphic setting. After ferns, the most abundant fossil plants present in these sections are impressions and compressions of angiosperm leaves, leaflets, and, to a lesser extent, fruits. Many of the leaves and leaflets are characterized by their pulvinate petioles or petiolules and the fruits consist of pods of various sizes and shapes, as in the Fabaceae or bean family (Judd et al. 2007). These fossils are found in all depositional units (except A, which, other than for the presence of rooting structures, is non-fossiliferous), but are most abundant in overbank layers (D, E, and F depositional units). Leaves and leaflets are most common in the basalmost part of all sections of Ash-IV and the thin set of strata below (CH-79, unit 3-5, CH-93, unit 2-4, CH-89, unit 2-4), where ferns are absent and where they co-occur with the leaves of Cynometra, discussed above. These assemblages form a continuous mat of leaves that are concentrated in two indurated clayey mudstone to siltstone layers that represent the boundary between Ash-IV and pre-Ash-IV. These units are stratigraphically continuous between sections and probably represent forest litter accumulation (E depositional unit; CH-79, unit 3-4, CH-93, unit 2-3, CH-89, unit 2-3). The widespread occurrence of Fabaceae remains in all sections may be related to their efficiency as N2-fixing plants for obtaining soil nutrients, which allows them to thrive in environmentally unstable habitats (Schulze et al. 2005). Whole to fragmented Fabaceae fruit compressions and impressions occur in several horizons throughout the sections, but appear more abundant in some overbank layers (F depositional unit) (i.e., CH-79, unit 18). Despite the common presence of legume fruits in some horizons, fern and parallel-veined monocot-like leaf impressions and compressions dominate in the overbank and lignitic horizons (D, E, F, G depositional units) occurring in the middle part of all sections. In these, ferns and monocots co-occur with impressions and compressions of Pandanites, a possible grass flower, poorly-preserved dicot leaves and leaflets (some of which are covered with extensive fungal hyphae networks), armed palm petioles, and fragments of plicate palm fronds (e.g., CH-79, unit 18-19, CH-93, unit 11-14, CH-89, unit 12-14). In most cases, ferns in these horizons are so abundant that it is not uncommon to find numerous dispersed sporangia, spores, megaspores, frond fragments, and other parts completely carpeting the fossil deposit. Some of these remains could have been produced by ferns other than those already described, including a fern that is preserved as contorted to flat-lying compressions in an overbank layer in the south section (F depositional unit, CH-79, unit 18). This fern consists of poorly-preserved creeping rhizomes bearing slightly grooved stems terminated in flabellate tufted fronds that divide dichotomously into linear segments. This fossil is most similar morphologically to Actiniopteris (Pteridaceae), a terrestrial taxon commonly found in rather arid settings in East Africa (e.g., lava hillsides, bushland and woodland, dry forests) (Johns 1991; Verdcourt 1999b; Smith et al. 2006). If this fossil is in fact an ancient representative of extant Actiniopteris, then its presence would indicate a seasonally arid environment, which would agree with the inferred environmental conditions based on the presence of other taxa (e.g., Marsilea) present in the same fossil horizon. Palm impressions and compressions, including Hyphaene, other spiny petioles of the Coryphoid and Calamoid type (Uhl and Dransfield 1987), and fragments of plicate fronds occur in airfall and the reworked tuff horizons (C and D depositional unit) towards the top of the central and north sections (CH-93, unit 23-25, CH-89, unit 24); plicate palm fronds are also found in stratigraphically equivalent airfall tuff layers in the south section (e.g., CH-79, units 24, 26). In addition, other palm frond fragments are found in some lignitic and airfall tuff horizons in all sections (e.g., CH-79, unit 18, CH-93, unit 14, CH-89, unit 26). Co-occurring with these palms and some of the described ferns (e.g., Cyclosorus) present in a number of tuff layers, are impressions of three morphotypes of poorly preserved dicot petiolate leaves, one of which consists of stipulate compound leaves with a serrate margin that are reminiscent of the extant genus Rubus (Rosaceae) (Hawthorne and Jongkind 2006). This is a cosmopolitan genus, with representatives in Africa occurring commonly in wet mountain regions (up to 3000 meters); some of these taxa are known as early colonizers of open patches of primary and secondary forest successions following volcanic or other types of disturbance (Spicer 1991; HHawthorne and Jongkind 2006; Wilmshurst and McGlone 1996; African Flowering Plants Database). Another monocot that is part of this plant assemblage consists of organically-preserved large hollow septate stems (culms), filled with ashy material, with enlarged internodes, much like bamboos (Poaceae) (Judd et al. 2007). This is present in two reworked tuffs (D depositional unit) (CH-79, units 24, 26) in the south section. Extant bamboos are known as successful colonizers (Judd et al. 2007), based on which an early colonizer role is suggested for the fossils present within Ash-IV. A previous record of bambusoid grasses preserved in airfall tuff layers from the Miocene of Kenya is consistent with the interpretation that bamboos were associated with disturbed environments in the past (Jacobs and Kabuye 1987). The distribution of plants within the three geologic sections discussed here indicates that riparian vegetation preserved as a forest litter accumulation horizon, which includes the genus Cynometra within the legumes family, typical of seasonal environments, and other dicot angiosperms of unknown affinity, characterized the early depositional stages of Ash-IV and pre-Ash-IV. The occurrence of plant assemblages consisting of in situ and transported aquatic and terrestrial ferns and a few angiosperms typical of ponded or wetland settings distributed in the lignitic horizons around the stratigraphic middle part of all sections is interpreted to represent a series of fluvial environments characterized by high water table inundated floodplains. The airfall and slightly fluvially-reworked horizons in the upper part of all sections is characterized by ecological pioneer taxa and other individuals of less clear affinity, among which are in situ preserved ferns and angiosperms, which is interpreted as indicating open ash-covered floodplains where airfall ash deposition preserved early ecological stages of plant community succession. In summary, the Ash-IV unit and the thin set of strata below it (pre-Ash-IV) contain a succession of plant communities that reflect the temporal physiographic variations of the environment resulting from increasingly frequent volcanic activity. |
|