Systematic Paleontology
Type Locality. Cerdas, Department of Potosí, Bolivia.
Distribution. Cerdas beds, early middle Miocene, Friasian sensu stricto and/or Collorcuran SALMA, Department of Potosí, Bolivia (Villarroel 1978;
Swanson et al. 1987;
Marshall and Sempere 1991;
MacFadden et al. 1995;
Croft et al. 2009); Nazareno Formation, ?middle Miocene, ?Colloncuran SALMA, Department of Potosí, Bolivia (Oiso 1991;
Hérail et al. 1996); Quebrada Honda, Honda Group, middle Miocene age, Laventan SALMA, Department of Tarija, Bolivia (Croft 2007).
Referred specimens. GB-223, left ramus bearing m1-3, right ramus bearing bases of p4-m3, half of a left mandibular corpus bearing i1-2 and dp3, and a palatal piece with the lingual halves of M2-3; UF 133803, partial cranium with complete maxillary dentition; UF 225690, complete left mandible with partial right (i1, p4); UF 225691, partial left mandible with p4-m3; UF 225692, left m1/2 and various bone fragments; UF 225693, right M2-3 and other tooth fragments; UF 225694, symphysis with bases of i1, right p4-m2, left m1, two partial lower molars; UF 225695, left M1/2; UF 225696, left mandible with m2-3, right mandible with partial m2; UF 225697, left mandible with m2 and partial m3; UF 225698, right M3 and partial left upper molar; UF 225699, left m3, partial mandible with right m2, partial m3, and partial symphysis; UF 225700, i1; UF 225701, left mandible with partial i2; right mandible with m2-3; UF 225706, lower incisor; UF 225709, right M1/2 and left P4-M1, various partial upper teeth; UF 225710, partial left mandible and symphysis including left i1-m2 and right i1-p4; UF 225711, partial left mandible with m3, various postcranial bones; UF 225713, two left M1/2, M3, i1, other tooth fragments; UF 225714, right mandible with p4-m3, numerous lower teeth/tooth fragments; UF 225721, various postcranial bones; UATF-V-000816, left M3; UATF-V-000839, right M2; UATF-V-000858, right M2-M3; UATF-V-000862, left M1-M3 and left M2 (isolated); UATF-V-000843 left P3, right P3-M3; UATF-V-000840, right and left P3-M3; UATF-V-000823, left dp3-4, m1-2; UATF-V-000827, right and left m1-m3; UATF-V-000879, right m1-m3; UATF-V-000844, left p4-m3; UATF-V-000815, left m1-m3; UATF-V-000847, right i1, m3, left p4, m3; UATF-V-000829, right m1; UATF-V-000835, left m1; UATF-V-000839, left m1; UATF-V-000837, p4; UATF-V-000874, m3; UATF-V-000845, left m1-m3.
Revised Diagnosis of
'Plesiotypotherium' minus
Differs from M. choquecotense in having an I1 that is ovoid, pointed distally with no lingual sulcus, and having a relatively wide post-palatal notch that does not extend to the level of M3. The molars increase in length from M1 to M3.
Differs from P. achirense in presence of more ovoid P3 and rounder lingual edge of P4; 'P.' minus is also smaller (based on dental measurements, approximately 30% smaller than P. achirense).
Differs from P. majus in possession of shallower buccal sulci on premolars and presence of more ovoid premolars; also smaller in size (based on dental measurements, approximately 45% smaller than P. majus).
Differs from Altitypotherium paucidens in the presence of parallel maxillary diastemata, an elliptical first maxillary incisor (subtriangular in A. paucidens), presence of P3, and subparallel lingual lobes on lower molars.
Differs from A. chucalensis in the presence of parallel maxillary diastemata, a wider post palatal notch, an anteriorly placed buccal sulcus on P3, a more distinct buccal sulcus on P4, subparallel lingual molar lobes, and a large M1 middle lobe.
Differs from Eotypotherium chico in having a rounder P3, a more subtriangular P4, in M3 being the longest tooth, in having the root of zygomatic arch above M2, and in having the middle lobe well exposed in all three molars; it is also larger (based on dental measurements, approximately 40% larger than Eotypotherium chico).
Differs from Eutypotherium in the presence of parallel diastemata, a postpalatal notch that does not extend to level of M3, an ovoid I1 with no lingual sulcus, one labial sulcus on premolar ectolophs, and smaller size (approximately 20% smaller than Eutypotherium lehmannnitschei, based on dental dimensions).
Differs from Typotheriopsis in presence of a shallow suborbital fossa and an ovoid I1 with no lingual sulcus; also approximately 30% smaller based on dental measurements.
Differs from Pseudotypotherium in the presence of a postpalatal notch that does not extend to level of M3, a shallow suborbital fossa, one labial sulcus on premolar ectolophs, a lesser degree of molar imbrication, proportionally shorter teeth with little exposure of the middle lobes on the maxillary molars, and smaller size (based on dental measurements, approximately 20% smaller than Pseudotypotherium exiguum).
Differs from Mesotherium in the presence of shallow suborbital fossa, ovoid I1 with no lingual sulci, lack of lingual sulcus on P4 (Mesotherium has one), absence of elongate M1, and lesser degree of molar imbrication. 'P.' minus is approximately 25% smaller than M. maendrum.
Differs from Caraguatypotherium in a more pronounced sagittal crest, a P4 without a lingual sulcus, lesser degree of molar imbrication, and smaller size (at least 20% smaller based on dental dimensions).
Description. Skull—The partial cranium of UF 133803 is in fair condition; the nasals, the anterior portion of the neurocranium, and the zygomatic processes of the temporal bone are not preserved. Much of the basicranium is missing except for portions of the right tympanic and the two pterygoids. The top of the neurocranium has the left and right parietals and squamosals, the sagittal crest, and a small portion of the frontal bone with an orbital process (right side only). The facial cranium preserves the right and left maxillae with full dentitions.
The palate exhibits some damage near the sagittal plane and restoration compound fills this damaged region. Many features of the palate are still intact, and at least six neurovascular foramina are present. It is apparent that UF 133803 had prominent incisive foramina; the left side is filled with restoration compound but the foramen is partially visible on the right and is approximately 7.5 mm long. The palate itself curves strongly toward the premolars and first two molars, then flattens slightly as it nears M3 (Figure 2). It is damaged on the left side between I1 and P3 (i.e., the diastema) but is plainly visible on the right. The diastemata are parallel and measure approximately 14.4 mm in length (Table 2).
 The palatal notch nearly reaches the level of the M3 and is acute (in contrast to the nearly 90° notch of P. achirense). The distance of the palatal notch from the back of the palate is much less than the width of the M3.
The palatal notch nearly reaches the level of the M3 and is acute (in contrast to the nearly 90° notch of P. achirense). The distance of the palatal notch from the back of the palate is much less than the width of the M3.
The nasal bones are missing, and the floor of the nasal cavity (typically formed by the maxilla) has been eroded away. This provides a view of the bases of the of the cheek teeth (Figure 3). The bases of the premolars and molars match the occlusal surfaces, indicating that these teeth are ever-growing (hypselodont), rootless, and would not have changed significantly with wear.
 Both zygomatic arches of UF 133803 are intact but the right shows more damage on the distal aspect of the temporal process than is seen on the left. The zygomatic bones are quite broad and heavy, and each has its root perpendicular to the anterior edge of M1.
Both zygomatic arches of UF 133803 are intact but the right shows more damage on the distal aspect of the temporal process than is seen on the left. The zygomatic bones are quite broad and heavy, and each has its root perpendicular to the anterior edge of M1.
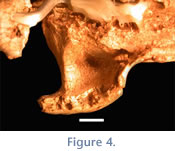
 The broad inferior surface present on both left and right arches apparently would have provided a large area for the origin of the masseter muscle. The suborbital fossa, present on the dorsal surface of the arch, is subtriangular and shallow, particularly in comparison with some other mesotheriines (e.g., Typotheriopsis) (Figure 4). The major axis of the fossa is transverse. A transverse strut of bone demarcates the posterior border of the suborbital fossa. The infraorbital foramen is subcircular, of medium size (relative to other mesotheriines), and measures 4.3 x 4.1 mm (Table 3;
Figure 5).
The broad inferior surface present on both left and right arches apparently would have provided a large area for the origin of the masseter muscle. The suborbital fossa, present on the dorsal surface of the arch, is subtriangular and shallow, particularly in comparison with some other mesotheriines (e.g., Typotheriopsis) (Figure 4). The major axis of the fossa is transverse. A transverse strut of bone demarcates the posterior border of the suborbital fossa. The infraorbital foramen is subcircular, of medium size (relative to other mesotheriines), and measures 4.3 x 4.1 mm (Table 3;
Figure 5).
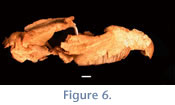 Only the posterior portions of the right and left frontals are present; the right orbital process that extends 13 mm posterolaterally to a broken distal edge. The frontals contact the parietals in an M-shaped suture; a thin tongue of the frontals extends posteriorly between the anterior extensions of the parietals. The prominent sagittal crest is approximately 39 mm in length and shows some minor damage (Figure 6). The left parietal shares a partial sinuous suture with the squamosal that is ~16.9 mm in length. The squamosal portion of the right zygomatic arch has two distinct neurovascular foramina. The glenoid fossa on the right side appears to have been compressed in an anteroposterior direction.
Only the posterior portions of the right and left frontals are present; the right orbital process that extends 13 mm posterolaterally to a broken distal edge. The frontals contact the parietals in an M-shaped suture; a thin tongue of the frontals extends posteriorly between the anterior extensions of the parietals. The prominent sagittal crest is approximately 39 mm in length and shows some minor damage (Figure 6). The left parietal shares a partial sinuous suture with the squamosal that is ~16.9 mm in length. The squamosal portion of the right zygomatic arch has two distinct neurovascular foramina. The glenoid fossa on the right side appears to have been compressed in an anteroposterior direction.
 Only the pterygoid wings and part of the basisphenoid are present on the sphenoid. Of the two sides, the left pterygoid complex is in better condition; although the lamina of the medial and lateral pterygoid plates are broken, the scaphoid fossa is preserved, which has been described as the origin of the tensor veli palatini muscle in toxodontians (Gabbert 2004). The right petrosal bulla is inflated but the entrance to the tympanic cavity has been crushed. The bulla is wider transversely than it is anteroposteriorly. The cerebellar aspect of the petrosal can be seen on both left and right sides. Due to damage of the neurocranium and basicranium, it is possible to see features of the petrosal bone inside the cranial cavity: the internal auditory foramen, subarcuate fossa, and the groove for the superior petrosal sinus (Figure 7).
Only the pterygoid wings and part of the basisphenoid are present on the sphenoid. Of the two sides, the left pterygoid complex is in better condition; although the lamina of the medial and lateral pterygoid plates are broken, the scaphoid fossa is preserved, which has been described as the origin of the tensor veli palatini muscle in toxodontians (Gabbert 2004). The right petrosal bulla is inflated but the entrance to the tympanic cavity has been crushed. The bulla is wider transversely than it is anteroposteriorly. The cerebellar aspect of the petrosal can be seen on both left and right sides. Due to damage of the neurocranium and basicranium, it is possible to see features of the petrosal bone inside the cranial cavity: the internal auditory foramen, subarcuate fossa, and the groove for the superior petrosal sinus (Figure 7).
Upper Dentition—In general, the teeth of UF 133803 are in excellent condition and exhibit few instances of breakage. Both left and right incisors are in excellent condition. They are elliptical with the major axis at approximately a 40-50° angle relative to the sagittal plane (Figure 6). No sulci or plications are present on the incisors. Incisors are hypselodont; the alveolus of each incisor is obvious because neither the nasal bones nor the floor of the nasal cavity is intact. A thin coat of cementum covers the lingual face of the incisor.
Both molars and premolars are hypselodont, as indicated by the open-ended bases of these teeth, which are visible due to the broken skull bones (Figure 3;
Table 4). The length of the cheek tooth row is ~5 cm. The left P3 is completely intact, whereas the right P3 has a small chip missing from both the posterior lingual and posterior buccal faces. Based upon the complete left P3, this tooth is subtriangular with a strong anteriorly placed sulcus on the buccal face of the tooth, forming a distinct parastyle. A more shallow sulcus is found centrally on the buccal face of the P3. No cementum is present on the right P3 but the morphology of the left P3 suggests that it was present. This tooth is featureless on the occlusal surface. Enamel evenly covers the anterior, buccal, and lingual faces of the tooth. The P3 is wider than it is long (Table 5). The P4 is intact on both the left and right sides. The P4 is also subtriangular, but less so than the P3, as the protocone lingual face is more rounded. A strong anterior sulcus is present on the buccal face of the P4, forming a parastyle. As in the P3, another sulcus is present on the central buccal face of the P4, but is not as pronounced. These sulci form the base of a prominent pillar evident in the occlusal outline of the P4. A thin coat of cementum is present on the buccal face of the tooth, and a much thicker coat is present on the lingual face. In an even coat, enamel covers the anterior, buccal, and lingual faces of the tooth, the posterior edge appears to be only covered in cementum. The P4 is wider than it is long (Table 5).
 As with all mesotheriines, UF 133803 exhibits three lingual lobes on all molars (Figure 8). The length of the lingual lobes increases in size anteroposteriorly (i.e., the anterior lobe is the narrowest). The overall orientation of the molar lobes is subparallel, with the anterior lobe subtly directed posterolingually and the posterior lobe subtly directed more anteriorly in all molars. The molars increase in length mesiodistally, with M3 being the longest. Molar cementum is thicker lingually and thinner buccally. An even thickness of enamel is present on the anterior, buccal, and lingual faces of the molars. Other than the lobes themselves, no other occlusal features are found on these teeth, and no sulci or plications are apparent on the lingual surfaces of the molars.
As with all mesotheriines, UF 133803 exhibits three lingual lobes on all molars (Figure 8). The length of the lingual lobes increases in size anteroposteriorly (i.e., the anterior lobe is the narrowest). The overall orientation of the molar lobes is subparallel, with the anterior lobe subtly directed posterolingually and the posterior lobe subtly directed more anteriorly in all molars. The molars increase in length mesiodistally, with M3 being the longest. Molar cementum is thicker lingually and thinner buccally. An even thickness of enamel is present on the anterior, buccal, and lingual faces of the molars. Other than the lobes themselves, no other occlusal features are found on these teeth, and no sulci or plications are apparent on the lingual surfaces of the molars.
The M1 is the shortest in the molar series. The more centrally placed buccal sulcus is the strongest sulcus on this surface of the tooth. This sulcus forms the base of a prominent rib that forms the paracone fold of the M1. The anterior lobe of M1 is the smallest of any of the molar teeth.
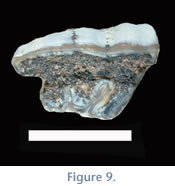 The middle lobe is trapezoidal in shape. The M2 is longer than the M1 and also exhibits a prominent centrally located sulcus forming the parastyle region, which is the largest of the three molar teeth. The M2 is the widest tooth in the cheek tooth series. The middle lobe is rounded and is the largest middle lobe of the three molars. The M3 is the longest and narrowest tooth in the cheek tooth series and has a trapezoidal outline due to the posterior projection of the ectoloph. The buccal sulcus is also apparent on the M3, but is less prominent than that on the previous two molars. The parastyle region is prominent on the M3. The middle lobe is the smallest and the most triangular in shape. The posterior ectoloph lobe is subtle in some teeth (e.g., UF 133803;
Figure 2 and
Figure 8), but more prominent in others (e.g., UF 225698;
Figure 9).
The middle lobe is trapezoidal in shape. The M2 is longer than the M1 and also exhibits a prominent centrally located sulcus forming the parastyle region, which is the largest of the three molar teeth. The M2 is the widest tooth in the cheek tooth series. The middle lobe is rounded and is the largest middle lobe of the three molars. The M3 is the longest and narrowest tooth in the cheek tooth series and has a trapezoidal outline due to the posterior projection of the ectoloph. The buccal sulcus is also apparent on the M3, but is less prominent than that on the previous two molars. The parastyle region is prominent on the M3. The middle lobe is the smallest and the most triangular in shape. The posterior ectoloph lobe is subtle in some teeth (e.g., UF 133803;
Figure 2 and
Figure 8), but more prominent in others (e.g., UF 225698;
Figure 9).
UF 133803 exhibits a degree of imbrication of 1.12 (Table 4;
Flynn et al. 2005). As mentioned above, the enamel is equally thick on all faces of the tooth where it is present, but there is none on the posterior surface of the cheek teeth and this is most obvious in M3. The cementum is thicker lingually and a thinner buccally. The cementum is thick on the middle lobes of the molars, particularly that of M3.
 Lower Dentition—Although characters of mesotheriine lower dentition do not allow for clear discrimination among genera (Croft 2007), lower teeth have been used to diagnose 'P.' minus (Villarroel 1978). From the Cerdas locality, 24 partial lower dentitions appear to pertain to 'P.' minus. Measurements for these specimens are provided in
Table 6. The sample represents the entire lower tooth row (Figure 10). The i1 is variable in shape, ranging from sub-ovoid to sub-triangular to trapezoidal. The enamel covers only the anterior face of the incisor and in one specimen (UF 225701), a sulcus is present on the medial surface of the tooth. The second incisor is elliptical in outline and less variable in shape, with enamel covering only the labial surface of the tooth. The p4 is triangular with a slight buccal sulcus demarcating the trigonid from the talonid; enamel is found on the buccal and lingual faces of the p4. The trigonid is smaller and narrower than the talonid. Both regions of the tooth show some variability in shape: the trigonid ranges from slightly ovoid to having a rounded point mesially; the talonid of the p4 ranges from rounded to more distally elongate. The three molars exhibit a buccal sulcus that nearly reaches the lingual edge of the tooth; a small isthmus of dentin persists between both lobes through later stages of wear. The molars also exhibit two lingual sulci, one along the posterior edge of the trigonid near the dentin isthmus and the other with various degrees of depth on the talonid near to where it tapers to its distal edge
Lower Dentition—Although characters of mesotheriine lower dentition do not allow for clear discrimination among genera (Croft 2007), lower teeth have been used to diagnose 'P.' minus (Villarroel 1978). From the Cerdas locality, 24 partial lower dentitions appear to pertain to 'P.' minus. Measurements for these specimens are provided in
Table 6. The sample represents the entire lower tooth row (Figure 10). The i1 is variable in shape, ranging from sub-ovoid to sub-triangular to trapezoidal. The enamel covers only the anterior face of the incisor and in one specimen (UF 225701), a sulcus is present on the medial surface of the tooth. The second incisor is elliptical in outline and less variable in shape, with enamel covering only the labial surface of the tooth. The p4 is triangular with a slight buccal sulcus demarcating the trigonid from the talonid; enamel is found on the buccal and lingual faces of the p4. The trigonid is smaller and narrower than the talonid. Both regions of the tooth show some variability in shape: the trigonid ranges from slightly ovoid to having a rounded point mesially; the talonid of the p4 ranges from rounded to more distally elongate. The three molars exhibit a buccal sulcus that nearly reaches the lingual edge of the tooth; a small isthmus of dentin persists between both lobes through later stages of wear. The molars also exhibit two lingual sulci, one along the posterior edge of the trigonid near the dentin isthmus and the other with various degrees of depth on the talonid near to where it tapers to its distal edge
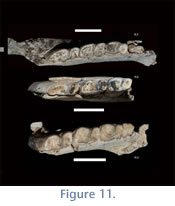 Deciduous Lower Teeth. Seven partial mandibles from Cerdas bear deciduous teeth. Only deciduous premolars (i.e., no deciduous incisors) are known at this time. The lower premolars resemble their adult counterparts although they are smaller in size. The dp3 (present on UF225704 and UATF-V-000823) is bilobed and resembles the adult p4 in having a triangular trigonid and a rounded talonid. The dp4 is also bilobate and resembles adult m1, but is smaller, with a trigonid more rounded than that of dp3. The talonid of dp4 is the most distinguishing feature of this tooth: the distal portion of the tooth is quite round, almost circular. In addition to size, distinction can be made among little worn adult m1s and dp4s by comparing talonid shape (dp4s have a very round distal end compared to p4s or m1s, which are more square; see
Figure 11.1, 11.3) and the depth of the buccal sulcus (deeper on dp4s than on adult p4s).
Deciduous Lower Teeth. Seven partial mandibles from Cerdas bear deciduous teeth. Only deciduous premolars (i.e., no deciduous incisors) are known at this time. The lower premolars resemble their adult counterparts although they are smaller in size. The dp3 (present on UF225704 and UATF-V-000823) is bilobed and resembles the adult p4 in having a triangular trigonid and a rounded talonid. The dp4 is also bilobate and resembles adult m1, but is smaller, with a trigonid more rounded than that of dp3. The talonid of dp4 is the most distinguishing feature of this tooth: the distal portion of the tooth is quite round, almost circular. In addition to size, distinction can be made among little worn adult m1s and dp4s by comparing talonid shape (dp4s have a very round distal end compared to p4s or m1s, which are more square; see
Figure 11.1, 11.3) and the depth of the buccal sulcus (deeper on dp4s than on adult p4s).
Tooth Wear in 'P.' minus. General wear trends seen in the Cerdas lower dentition sample include the obliteration of the enamel faces on the dp3 and wear of the trigonid enamel on dp4. The only other noticeable features likely associated with wear are a deepening of the lingual sulcus on the distal edge of the talonid, and sharpening of the distal point of the m3 talonid.
Billet et al. (2008) noted that the lingual sulcus deepens with wear in the deciduous teeth of trachytheriines. Although definitive wear categories could not be identified in the Cerdas material, specimens that exhibited both permanent and deciduous dentition had smaller adult teeth (shorter and narrower). Those specimens without associated deciduous teeth had larger (longer and broader) teeth, particularly in the first lower molar. The teeth associated with deciduous dentition and considered to be early wear are 10-15% shorter in length and ca. 5% narrower in width than those with a greater degree of wear (Table 6).
Remarks. We refer the maxillary specimens listed above to 'P.' minus based on dental similarities in maxillary tooth lobe shape and orientation and the following characters: relatively large infraorbital foramen; an angle of the major axis of the incisor near 50°; a low degree of imbrication; and slightly posterolaterally divergent diastemata.
Villarroel (1974a) indicated that the genus Plesiotypotherium could exhibit slightly posterolaterally divergent diastemata, although the type exhibits parallel diastemata.
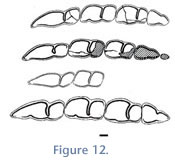 The assignment of mandibular specimens to 'P.' minus is based on similarity with the P. minus holotype in size and in morphology of the molar series (i.e., the presence of both an anterior and talonid lingual sulcus;
Figure 12). As indicated by our phylogenetic analysis (below), the Cerdas sample does not share an exclusive relationship with specimens from Achiri that constitute the genus Plesiotypotherium; we thus cannot place the Cerdas sample in the genus Plesiotypotherium. The older age of the Cerdas sample is compatible with this assignment. However, further morphological and metric evaluation of the Cerdas sample in relation to the similarly aged Nazareno sample could clarify the taxonomic status of 'P.' minus.
The assignment of mandibular specimens to 'P.' minus is based on similarity with the P. minus holotype in size and in morphology of the molar series (i.e., the presence of both an anterior and talonid lingual sulcus;
Figure 12). As indicated by our phylogenetic analysis (below), the Cerdas sample does not share an exclusive relationship with specimens from Achiri that constitute the genus Plesiotypotherium; we thus cannot place the Cerdas sample in the genus Plesiotypotherium. The older age of the Cerdas sample is compatible with this assignment. However, further morphological and metric evaluation of the Cerdas sample in relation to the similarly aged Nazareno sample could clarify the taxonomic status of 'P.' minus.
We refer the specimen assigned to Microtypotherium cf. M. choquetotense by
Villarroel (1978) to 'P.' minus because the specimen has no characters that uniquely suggest it pertains to Microtypotherium (e.g., m2 proportions > 1.60), or that suggest that it is more closely related to that species than to 'P.' minus. Additionally, this specimen falls well within the size range of the 'P.' minus sample (Figure 12).
 Mesotheriinae, gen. et sp. indet.
Mesotheriinae, gen. et sp. indet.
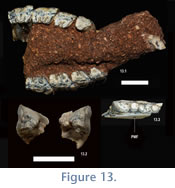
Referred Specimens. UF 225707, right mandibular fragment bearing m2-3 (Figure 13); UATF-V-000840 partial rostrum with right and left I1, ?P4-M3 (left I1 and both ?M3 loose; see below for alternate interpretation of the tooth homologies;
Figure 13).
Comments. These specimens are the smallest mesotheriines at the Cerdas locality (Figure 13). Both are clearly mesotheriine based on their dental morphology: UF 225707 exhibits no fossettids in either the trigonid or talonid (unlike trachytheriine mesotheriids), a slight lingual sulcus on the m2 talonid is present and the maxillary dentition exhibits imbrication (a feature of mesotheriines). Observation of dental and gnathic morphologies indicates that one specimen (UF 225707) is apparently an adult whereas UATF-V-000840 is a juvenile with a full adult tooth row other than an unerupted M3 (Figure 13).
 UF 225707 does not differ in morphology from unworn specimens of m2-3 seen in the 'P.' minus sample: both teeth exhibit a rounded trigonid and a slightly pointed talonid, with the m3 exhibiting a more elongate and pointed talonid. The lingual enamel face appears either worn or broken off in this specimen. The enamel that frames the labial sulcus is also worn or broken. The mental foramen of this specimen sits squarely underneath the trigonid of the m3. In Trachytheriinae, placement of the mental foramen underneath the distal m2 is suggestive of an advanced stage of development, such as adulthood (Billet et al. 2008).
Villarroel (1974a) illustrated the posterior mental foramen on Plesiotypotherium achirense (an adult specimen) beneath the right m2 trigonid, but mentioned nothing about it in the text.
UF 225707 does not differ in morphology from unworn specimens of m2-3 seen in the 'P.' minus sample: both teeth exhibit a rounded trigonid and a slightly pointed talonid, with the m3 exhibiting a more elongate and pointed talonid. The lingual enamel face appears either worn or broken off in this specimen. The enamel that frames the labial sulcus is also worn or broken. The mental foramen of this specimen sits squarely underneath the trigonid of the m3. In Trachytheriinae, placement of the mental foramen underneath the distal m2 is suggestive of an advanced stage of development, such as adulthood (Billet et al. 2008).
Villarroel (1974a) illustrated the posterior mental foramen on Plesiotypotherium achirense (an adult specimen) beneath the right m2 trigonid, but mentioned nothing about it in the text.
 Since the position of the posterior mental foramen in adults of both P. achirense and Trachytheriinae is beneath the m2, it is likely that UF 225707 represent an adult. UF 225707 exhibits a smaller m2 than any known specimen of E. chico, although the m3 falls within the range of the diminutive Chucal species, which is 20-25% smaller than the other species at Chucal (Figure 14.1). Such a significant size difference with an erupted m3 suggests that UF 225707 is a different species. Also, this specimen is considerably smaller than the adult dentitions associated with deciduous teeth in the 'P.' minus sample (Figure 15).
Since the position of the posterior mental foramen in adults of both P. achirense and Trachytheriinae is beneath the m2, it is likely that UF 225707 represent an adult. UF 225707 exhibits a smaller m2 than any known specimen of E. chico, although the m3 falls within the range of the diminutive Chucal species, which is 20-25% smaller than the other species at Chucal (Figure 14.1). Such a significant size difference with an erupted m3 suggests that UF 225707 is a different species. Also, this specimen is considerably smaller than the adult dentitions associated with deciduous teeth in the 'P.' minus sample (Figure 15).
UATF-V-000840 is more difficult to interpret, though we tentatively interpret it as including P3-M3 (only the dentition is described here, pending further preparation). In addition to the incisors, five cheek teeth are preserved on each side. The posteriormost are either unerupted or just recently erupted (Figure 13). The erupted maxillary teeth, particularly the molars, also resemble specimens of Eotypotherium chico in size and morphology (Figure 14.2), including a fossette on the triangular P3, a quadrate P4 with an enclosed middle lobe, and extensive lingual exposure of the middle lobes of both M1 and M2.
The unworn M3s associated with this specimen clearly indicate it is an example of early wear. Plots of the M1 and M2 of UATF-V-000840 indicate that these teeth are much smaller than fully adult specimens from Cerdas. Based on dental dimensions, these upper teeth are 11-14% shorter in length than more worn teeth and 9% smaller in width than the teeth of UF 133803 (the skull).
Given the presence of a trilobed P4 and a fossette on the P3, it is also possible that UATF-V-000840 is a juvenile bearing dP3-4 and M1-3. Francis (1965) reported that the dP2-3 of Pseudotypotherium (his DM2-3) was subtriangular and that the dP4 was subquadrate, similar to the permanent P4.
Francis (1965) also noted that the dP4 of Mesotherium was trilobed, similar to the permanent M1, although the deciduous tooth was much shorter than the adult molar. We favor an interpretation of P3-M3 for two reasons: first, the teeth interpreted as M1-2 are very similar in size; given the interpretation of
Francis (1965) this would not be expected if the teeth were dP3-M1. Second, due to missing bone, the full lengths of the teeth are visible in buccal view and there is no indication of roots (suggestive of deciduous teeth in some typotheres) nor are replacement teeth evident in the teeth that could be interpreted as P3-4 (see
Billet et al. 2008 and
Billet et al. 2009 for remarks on deciduous typothere teeth).
Remarks. These two unassociated specimens are substantially smaller than any others at Cerdas. They likely represent a second species, but it is also possible that at least UATF-V- 000840 is a juvenile of 'P.' minus. Until more specimens are found, we cannot confidently interpret the identities of these teeth in this specimen.
Phylogenetic Position of 'P.' minus
To evaluate the position of 'P.' minus relative to P. achirense, we performed a phylogenetic analysis of mesotheriines based on the data matrix of
Croft et al. (2004;
Table 7 and
Table 8), but coded 'P.' minus separately from Plesiotypotherium from Achiri (= P. achirense and P. majus). The analysis resulted in 103 equally most-parsimonious trees of 77 minimum possible and 202 maximum steps. The strict consensus tree resembles that of
Croft et al. (2004) for the younger mesotheriines from Argentina and Chile but shows less resolution near the base of the tree (Figure 15).
'P.' minus is part of a polytomy at node 4 that includes Altitypotherium, M. choquecotense, and a clade of all other mesotheriines except Eotypotherium chico (which is the basal-most mesotheriine). Character state changes at node 4 include: rodentlike rostrum shape (character #1:2), a very developed zygomatic plate (character #8:2), a subtriangular P3 (character #26:1), and a subtriangular and short P4, without a central fossette (character #19:1). For three characters (#3, 5, and 11) at least one member of the polytomy could not be coded. Seven characters (#1, 2, 4, 8, 19, and 26) are equivocal at this node; character state changes using ACCTRAN and DELTRAN optimization are reported in
Table 9.
'P.' minus shares no derived character state with Plesiotypotherium (P. achirense). 'P.' minus differs from P. achirense in displaying primitive character states for six characters: diastemata shape (character # 4:1), shape of I1 and presence of sulcus (character # 16:0), shape of P4 and lack of fossette (character # 19:1; P. achirense is polymorphic for this trait, exhibiting both states 1 and 2), the ratio of i1/i2 ('P.' minus is polymorphic for this character (# 22), exhibiting both states 0 and 1), section of i1 (character # 23:1), and lingual exposure of M1 middle lobe (character # 31:0). 'P.' minus is excluded from the clade of higher mesotheriines (including P. achirense; node 5,
Figure 13) by the absence of two derived character states: i1 subtriangular, trapezoidal or elliptical in section and exhibiting a lingual sulcus of varying depths (character # 23:2, 3, 4) and extensive lingual exposure of the M1 middle lobe (character # 31:1).
'P.' minus shares 21 derived states with both Altitypotherium species and 25 with Microtypotherium. 'P.' minus differs from Altitypotherium in having a better-developed suborbital fossa (character # 6:2), an oval I1 (character #16:0), and subparallel sides of the M1 middle lobe (character # 30:1). 'P.' minus differs from Microtypotherium in exhibiting m2 proportions between 1.60 and 2.30 (character # 32:0); Microtypotherium exhibits a unique state, with m2 proportions less than 1.60 (character # 32:2).
The 'P.' minus sample from Cerdas does not pertain to the genus Plesiotypotherium, nor can it be referred to Microtypotherium or Altitypotherium based on this phylogenetic analysis. Rather, the sample appears to represent another branch within the radiation of small early to middle Miocene mesotheriines common in Bolivia and northern Chile. Additional craniodental material of this species and other early mesotheriines should help clarify relationships among these forms and potential links with later species.
Variation in Small Mesotheriines: The Effects of Tooth Wear and Ontogeny on Taxonomy
Sources of dental variation in small mesotheriines are not clear, making it difficult to determine whether differences in tooth size are due to size sexual dimorphism, ontogenetic change, or tooth wear. There are two potential models for variation in tooth size due to ontogenetic factors: 1) non-hypselodont mammals, where deciduous teeth are generally smaller than adult teeth and exhibit a unique morphology; and 2) euhypsodont rodents, which show variability in the size of their dentition throughout ontogeny, though the morphology of the deciduous tooth does not vary greatly from the adult tooth (Kramarz 2002).
Because our sample consists primarily of lower teeth and because they have few distinguishing features useful for taxonomy, we explored two potential sources of variation in the Cerdas 'P.' minus sample: dental wear and ontogeny (potentially as indicated by posterior mental foramina placement). Wear has long been known to be a confounding factor in understanding taxonomic variation in mesotheriines.
Francis (1965) noted that previous workers, such as
Cabrera (1937,
1939), had remarked upon the widening of the molars during the extended development of mesotheriines: "...un franco engrosamiento dentario que se prolonga durante todo el período de desarrollo del animal" (p. 8). Ontogenetic patterns (age of epiphyseal closure, dental eruption sequences) are not well documented for mesotheriines, although
Billet et al. (2008) described an intriguing developmental sequence in the movement of the posterior mental foramen in trachytheriine mesotheriids. The position of the posterior mental foramen has been documented in other typotheres (see
Simpson 1967), but
Billet et al. (2008) demonstrated that as individuals of Trachytherus grow, this foramen moves from beneath the dp4 (in young individuals) to beneath the talonid of the m2 (in older individual).
Until recently, tooth wear had not been classified into meaningful stages within a sample of typotheres.
Croft et al. (2003) documented that wear has such a great effect on tooth dimensions in archaeohyracid typotheres that it must be considered prior to identifying specimens. These authors split the Tinguiririca sample into wear categories due to the high coefficient of variation in this single species sample as a whole. By splitting the sample into broad wear categories, the CVs were greatly reduced. Recently,
Billet et al. (2008) recognized a series of eight ontogenetic/wear stages for a very large sample of trachytheriine mesotheriids from Salla, Bolivia. This study showed striking changes in tooth size due to wear for both maxillary and mandibular molars. The
Croft et al. (2003) and
Billet et al. (2008) studies have shown that wear has a profound effect on the size of teeth throughout the lifetime of an individual in certain typotheres. These studies suggest that dental wear can skew both the perception of dental variation and possibly the taxonomic composition of the total sample. When metrically evaluating a sample of typothere dentitions, the effects of wear must be considered prior to any assessment of population variation
Croft et al. (2003).
Although the studies of Croft et al. (2003) and
Billet et al. (2008) showed that dental dimensions changed with wear, the pattern presented by each was slightly different.
Croft et al. (2003) found that M1-2/m1-2 of archaeohyracids became shorter and broader with wear, and that M3/m3 increased in length.
Billet et al. (2008) noted that the trachytheriine M1 shortened with wear, and that the M3 lengthened with wear. These authors also reported that m1-2 broadened with wear and m2-3 lengthened (Billet
et al. 2008).
In the Cerdas 'P.' minus assemblage, similar wear-based trends were observed. We could only assess wear on mandibular molars because we did not have a large sample of maxillary teeth. It is not possible to establish more than two wear categories with the Cerdas mesotheriines due to a lack of wear-related changes in occlusal features (e.g., fossettids) on the lower dentition. Fortunately, many specimens had both deciduous and adult teeth preserved in the lower tooth row; the presence of deciduous premolars can demonstrate the degree of wear present on adult molars based on age, the assumption being that if deciduous premolars are present in the tooth row, then the adult molars have erupted more recently and thus have less wear than those unassociated with deciduous teeth. This pattern of maintaining the dp4 with a fully erupted set of molars is a pattern seen in modern ungulates with rapid growth as a part of their life history pattern (Smith 2007). It is only after the full set of molars has erupted that the anterior dentition begins replacement (Smith 2007).
Two groups were identified within the 'P.' minus sample: adult molars associated with deciduous teeth or a newly erupted m3 and those without.
Table 10 lists the univariate statistics for the 'P.' minus sample as a whole, as well as for the two wear groups. As for other typothere groups, when wear is not considered, the CVs are very high for each tooth position. When those specimens are grouped by wear, CVs decrease substantially. Even these two wear groups still have very high coefficients of variation, however, suggesting that one or more unrecognized age/wear categories may be present.
In a plot of m1 dimensions (Figure 14.2), specimens with deciduous or newly erupted teeth are smallest, as would be expected. The remaining specimens are wider and longer. This trend is also apparent in m3 and less so in m2 (Figure 14.2). It should theoretically be possible to erect at least one more wear category for these mesotheriines that would be indicative of a middle phase of wear (Figure 14.2), but without occlusal features that vary with wear (as in archaeohyracids or Trachytherus), this may not be possible.
These data demonstrate that, as mesotheriid teeth undergo wear, they do change shape, a phenomenon reported for other typotheres (see
Croft et al. 2003;
Billet et al. 2008). In this sample, the m1s of juveniles (with less wear) were 26% shorter and 23% narrower than adult m1s, juvenile m2s were 19% shorter and narrower, and juvenile m3s were 22% shorter and 24% narrower (Table 6;
Figure 14.2). Adult teeth associated with deciduous teeth or those that are newly erupted are the smallest overall (shortest and narrowest; e.g., UATF-V-000879) for all tooth positions.
Smith (2007) reported data for a series of ungulates showing that eruption patterns varied with life history patterns. She noted that in ungulates that had rapid growth rates, such as Antidorcas (the springbok), deciduous teeth are replaced only after permanent molars have erupted (Smith 2007). This pattern fits what is seen here in the 'P.' minus sample; in UATF-V-000879, dp4 is maintained through the eruption of m3. This suggests that 'P.' minus might have had a rapid growth as part of its life history, but without more data regarding the replacement of the anterior deciduous dentition, this remains speculative.
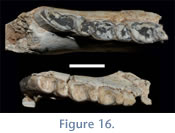 In addition to the lengthening and widening of teeth that occurs in more worn specimens, another dental feature emerges with wear: the presence of a sulcus on the lingual faces of the talonids. This feature is found on all three molars. It is absent or very subtle in younger specimens with less wear, but obvious on older individuals (Figure 16).
In addition to the lengthening and widening of teeth that occurs in more worn specimens, another dental feature emerges with wear: the presence of a sulcus on the lingual faces of the talonids. This feature is found on all three molars. It is absent or very subtle in younger specimens with less wear, but obvious on older individuals (Figure 16).
The posterior mental foramen is visible in six 'P.' minus specimens. Because it is visible in so few, we used it as a confirmatory method of relative age/development. No specimens from Cerdas have a posterior mental foramen below the m2 talonid (other than UF 225707, referred to Mesotheriinae indet.). In those specimens, in which the foramen is preserved, the most distal position in which it is found is below the m2 trigonid. In UATF-V-000823, a specimen with deciduous teeth, two mental foramina are present, the posterior under dp4, the anterior under dp3. Specimen UATF-V-000879, with retained dp4, exhibits the posterior mental foramen under the m1 talonid. Three specimens have the posterior foramen between m1 and m2 (UF 225690, 225691, and 225714).
 A very worn specimen, UATF-V-000815, has a mental foramen under the trigonid of m2 (with wear determined by presence of lingual grooves and overall larger size). Based on the work of
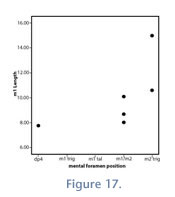
Billet et al. (2008) and a specimen with deciduous teeth (UATF-V-000823), it is clear that mental foramina could be useful for establishing relative individual ages. When dividing the Cerdas sample into those specimens bearing deciduous dentition (with smaller permanent teeth) and those with only permanent dentition (larger teeth), an intermediate group appears (Figure 14.2). Those intermediate specimens also exhibit a posterior mental foramen at the m1-2 junction, indicating that they are not of advanced age (Figure 17). If it is true that these animals add adult molars before replacing the anterior deciduous dentition, it seems reasonable to assume that the posterior mental foramen would "move" further back as larger anterior teeth were added to the arcade.
A very worn specimen, UATF-V-000815, has a mental foramen under the trigonid of m2 (with wear determined by presence of lingual grooves and overall larger size). Based on the work of
Billet et al. (2008) and a specimen with deciduous teeth (UATF-V-000823), it is clear that mental foramina could be useful for establishing relative individual ages. When dividing the Cerdas sample into those specimens bearing deciduous dentition (with smaller permanent teeth) and those with only permanent dentition (larger teeth), an intermediate group appears (Figure 14.2). Those intermediate specimens also exhibit a posterior mental foramen at the m1-2 junction, indicating that they are not of advanced age (Figure 17). If it is true that these animals add adult molars before replacing the anterior deciduous dentition, it seems reasonable to assume that the posterior mental foramen would "move" further back as larger anterior teeth were added to the arcade.
Comparisons with other Bolivian and Chilean Mesotheriines. The Cerdas, Chucal, and Nazareno assemblages all have at least one striking similarity: at least one large and one small mesotheriine. At Chucal, the two species of Altitypotherium overlap in size to some degree, but they clearly differ in morphology (particularly the presence of P3); Eotypotherium chico is markedly smaller, by 20-25%, compared to Altitypotherium (Croft et al. 2004).
C hucal. Both A. paucidens and A. chucalensis exhibit low and consistent sample variation (Table 10), as typically characterizes single species samples (Table 10,
Figure 18;
Cope and Lacey 1992,
1995;
Cope 1993,
Plavcan and Cope 2001). Eotypotherium chico exhibits small CVs for all but three dental measurements: M1 and m1 width, and m3 length (Table 10,
Figure 18). These high CVs are likely attributable to small sample size but wear also could be a factor (e.g., SGO PV 5158 exhibits a rather long m3; see
Table 10). In general, the overall low CV values for Chucal taxa lend support to the validity of these species.
hucal. Both A. paucidens and A. chucalensis exhibit low and consistent sample variation (Table 10), as typically characterizes single species samples (Table 10,
Figure 18;
Cope and Lacey 1992,
1995;
Cope 1993,
Plavcan and Cope 2001). Eotypotherium chico exhibits small CVs for all but three dental measurements: M1 and m1 width, and m3 length (Table 10,
Figure 18). These high CVs are likely attributable to small sample size but wear also could be a factor (e.g., SGO PV 5158 exhibits a rather long m3; see
Table 10). In general, the overall low CV values for Chucal taxa lend support to the validity of these species.
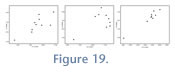 Nazareno. The only published study from Nazareno is that of Oiso (1991), though we were able to study additional specimens from this locality in UATF collections. The UATF Nazareno sample is within the range of variation of the
Oiso (1991) sample (Figure 19). We evaluated CVs from Nazareno in three ways: using the Oiso data alone, using the UTAF data alone, and evaluating the combined samples. The Oiso (1991) data have very high CVs for all tooth positions (>10;
Table 11). It appears that at least two taxa are present, one large and one small, and possibly more. Our reasoning for suggesting at least two taxa is that some of the smaller specimens have fully erupted third molars, indicating a later juvenile stage or adulthood. The UATF dataset has much lower CVs, all < 10, even when corrected for small sample sizes. The combined assemblage has very high CVs. The bivariate plot of m1 and m2 dimensions shows a size distribution pattern similar to that reported for Chucal and Cerdas: one large and one small size group (Figure 19), though the pattern is less apparent in the m3 plots.
Nazareno. The only published study from Nazareno is that of Oiso (1991), though we were able to study additional specimens from this locality in UATF collections. The UATF Nazareno sample is within the range of variation of the
Oiso (1991) sample (Figure 19). We evaluated CVs from Nazareno in three ways: using the Oiso data alone, using the UTAF data alone, and evaluating the combined samples. The Oiso (1991) data have very high CVs for all tooth positions (>10;
Table 11). It appears that at least two taxa are present, one large and one small, and possibly more. Our reasoning for suggesting at least two taxa is that some of the smaller specimens have fully erupted third molars, indicating a later juvenile stage or adulthood. The UATF dataset has much lower CVs, all < 10, even when corrected for small sample sizes. The combined assemblage has very high CVs. The bivariate plot of m1 and m2 dimensions shows a size distribution pattern similar to that reported for Chucal and Cerdas: one large and one small size group (Figure 19), though the pattern is less apparent in the m3 plots.