|
|
|
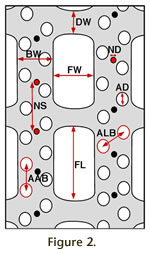
Materials and MethodsSpecies of biserial fenestrates that are characterized by dichotomous branching and branch linkage by dissepiments, either described in literature published from 1980 to 2007 or identified and characterized in unpublished post-1980 data sets available to us, were considered for this study. Species accepted for inclusion in the study (Appendix) had to meet certain criteria: 1) Assignment to genus by the author of the paper or data set based on the contemporary criteria of zooecial characteristics visible in thin sections plus geometry of the keel node sequence (i.e., whether in uniserial or biserial rows), plus 2) a minimum set of measurements made from the exterior surface in addition to information from thin sections used by the author for discrimination of fenestellid genera. Data for this study were compiled from nine sources using the criteria given above (McKinney and Kriz 1986; Snyder 1991; Holdener 1994; McColloch et al. 1994; Nakrem 1994; Nakrem 1995; Ernst 2001; Ernst and Schroeder 2007; and McKinney, unpublished data). Each of the 15 genera used in this study is represented by 12 to 273 composite OTUs from 1 to 9 species per genus (Appendix). Two species included in the study, Lyroporella serissima (Nakrem 1995) and Anastomopora anaphora (McColloch et al. 1994) are not biserial fenestellids but were intended to serve as reference points. Their branch organization differs from fenestellids in having three or more rows of zooecia except proximal to branch bifurcations. If the position of the two genera in morphospace were isolated from a tightly clustered cloud comprised of all the fenestellid species included herein, then any differences in position of the multispecies fenestellid genera within the cloud would indicate relatively trivial differences in gross morphology. If, however, the two extraneous species of Lyroporella and Anastomopora were within or contiguous with the fenestellid cloud, then any notable differences in distribution of multispecies fenestellid genera within the cloud could be interpreted to represent appreciable differences in gross morphology among genera. Lyroporella serissima (Nakrem, 1995) has branches organized like those of Polyporella, with two rows of zooecia distal to bifurcations but proliferating to three rows at an appreciable distance preceding the next bifurcation. Nakrem (1995) followed the understanding of McKinney (1994) that such lyre-shaped fenestrates are species of the genus Lyropora. McKinney (1994) had missed that Miller (1889) listed Fenestella (Lyropora) lyra Hall as type species of Lyropora and thought that Ulrich's 1890 designation of F. (Lyropora) quincucialis Hall as type species of Lyropora was the earliest. Thinking that F. (Lyropora) quincucialis Hall had been the earliest designated type species of Lyropora, Simpson's designation (Simpson 1897) of the same species as type species of Lyroporella Simpson, 1895 was considered to make Lyroporella a junior objective synonym of Lyropora. Lyroporella is a different genus from Lyropora based on the type species F. (Lyropora) lyra, which has polyserial zooecial rows in branches, requiring reassignment of Nakrem's (1995) species Lyropora serrisima to Lyroporella. The second is a species of Anastomopora, which typically has multiserial branches. However in some species – such as Anastomopora anaphora (McColloch et al. 1994) included here – branch segments begin with only two rows of zooecia after a bifurcation before interpolating additional rows of zooecia. McColloch et al. (1994) named the species Reteporidra anaphora. We have reassigned it to Anastomopora Simpson, 1897, because we consider Reteporidra Nickles and Bassler (1900) (nom. nov. pro Reteporella Simpson 1895, non Busk 1884) to be a junior subjective synonym of Anastomopora Simpson 1897. Morphological Characters
N It is beyond the scope of this paper to infer evolutionary significance of the differentiation of character states among fenestrate bryozoan genera; however, the relationship between skeletal morphology and biologically and ecologically significant soft-part characters has been described (e.g., Winston 1977, 1978; McKinney and Jackson 1991). Types of DataThe fundamental datum in this study is a "composite OTU." Each data point in the following figures of PCA scatter plots consists of a vector of the nine measured characters collected from the same colony. This datum represents an idealized OTU at the level of an individual module in the colony, but limitations do not allow for measurement of all nine characters from a single zooecium (e.g., meshwork vs. zooecial characters). Correlation of characters within a single composite may or may not have real significance. Composite OTUs in this study are represented by two kinds of data. Raw Measurements in Millimeters. For specimens where original measurements were available, most measurements were measured from multiple colony fragments, identified by the original author as conspecific (Appendix, Data Type = measured). Typically 12 to 24 measurements were available per species. In some cases many (100 to 200) measurements per species were available. In other cases very few (2 to 4) composite OTUs could be constructed from available raw measurements. Scores Reconstructed from Summary Statistics. For specimens where original measurements were not available, 12 composite OTUs were reconstructed using published summary statistics (mean, standard deviation, minimum and maximum values) using Normal Order Deviates (see Hageman et al. 2009). The goal of this exercise was to approximate the multivariate hyper-volume represented by 12 composite OTUs (Appendix, Data Type = reconstructed). No assumptions were made about reconstructed scores representing the distribution and covariate structure of original data, only that their overall position in multivariate space would approximate that of normally distributed samples with an equivalent mean and variance. Table 1 provides an example of the calculation. The steps include: 1. Obtain summary statistics for measured values associated with a published description, preferably with five or more observations per character, reflecting measurements compiled from three or more specimens (Table 1). 2. Identify the Normal Order Deviates for n= number of observations (here 12) from a sample with a normal distribution, mean of zero and standard deviation of one (Rohlf and Sokal 1981, table 37; Hageman 1992) see Table 2, column NOD. 3. Reconstruct 12 scores for each character using the equation:
This results in 12 scores that have the approximate distribution (mean and sample deviation) as the original measured sample (Table 2). 4. For each character and each putative species, compare values of observed vs. reconstructed mean, standard deviation, minimum, and maximum (Table 1). In some cases, some reconstructed values were modified by hand in order to better approximate the observed distribution and summary values (Table 2, highlighted red). 5. Within each column (character), reconstructed scores were randomized independently by column. Rows resulting from the independent randomization by columns provide 12 composite OTUs. As a result, there is no natural correlation among characters within a reconstructed OTU, but this process removes the primary linear relationship and helps to maximize the hyper-volume occupied by the samples in multidimensional morphospace. Combination of Raw and Reconstructed Data. In a few cases, where full suites of measured data were available for most specimens, but only a partial suite was available for one or two characters (e.g., unmeasurable from available material), the full suite of composite OTUs was created by reconstructing scores for the missing data using methods above. These taxa include: Apertostella crassata, Hemitrypa bohemicus, Hemitrypa mimicra, Hemitrypa tenella, and Laxifenestella digittata. Transformation of DataThe completed data set (1075 composite OTUs x 9 Characters) was transformed by standardizing the data (Z-score), where the mean and standard deviation were calculated across all OTUs for each character. The values for transformed OTUs are expressed in units of standard deviation from a mean of zero. This removes differences among characters measured at different scales, so that analyses will emphasize relative variation and not emphasize absolute scale. Principal Components AnalysisPrincipal Components Analysis was performed with the software PAST v. 1.81 (Hammer et al. 2001). Eigen values, the cumulative percentage of variance explained by each, the loading coefficients for each axis and PCA scores for each composite OTU on all nine axes were saved for evaluation. Contribution of Individual Characters to Principal Component Axes. The relative correlation (importance) of each of the nine morphometric characters to each of the PCA axes was evaluated by examining the loading coefficients. Important characters for PCA axes 1 to 5 (largest absolute values) were highlighted by annotating representative scatter plots with idealized illustrations of specimens with end-member states for the character. A variance-covariance matrix was also calculated using PAST v. 1.81 (Hammer et al. 2001) in order to evaluate relationships among characters. Generating Three-Dimensional (3-D) Scatter Plots and Animations. A variety of three- dimensional graphs were created using the "Scatterplot 3-D" function in JMP (PCA scores exported from PAST via text file), with the goal to: 1) characterize overall variation within and among all genera, 2) illustrate variation accounted for by different principal component axes, and 3) highlight differences among closely related genera (plotted in a morphospace defined by all genera). Animated files of three-dimensional scatter plots were generated using the following procedure: 1) graphs were rotated to a standard starting orientation with the intersect of negative axes in the lower center of screen, 2) the plot was saved as a jpeg file, 3) plot was rotated 6°, 4) steps 2 and 3 were repeated through 360°, generating 60 jpeg files, 5) all files were scaled 60% using the batch image converter application Resizeit v. 2.3.1 (SYWSoft), and 6) an animated gif file was created using the application GIFfun v. 4.2 (Stone Design Corp.), with a 0.13 second delay between frames. Data Subsets in Scatter Plots and 3-D Animations. The large amount of data employed in this study makes visual interpretation of the entire 1075 OTU data set difficult. Nevertheless, the following plots were generated: 1) All genera, PCA 1 vs. 2 vs. 3, 2) All genera, PCA 3 vs. 4 vs. 5 and 3) All genera, PCA 6 vs. 7 vs. 8. In order to see relationships among genera more clearly, subsets of the data (all calculated in the same multivariate space) were plotted individually. The genus Rectifenestella was used as a standard for comparison in many of these plots. |
|
| Fenestate Morphospace Plain-Language & Multilingual Abstracts | Abstract | Introduction | Materials and Methods Results | Discussion | Summary | Acknowledgements | References | Appendix Print article |